
Attenuation of Aeromonas hydrophila Infection in Carassius auratus by YtnP, a N-acyl Homoserine Lactonase from Bacillus licheniformis T-1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Plasmid and Reagents
2.3. Cloning and Sequencing of the AHL Lactonase Gene Ytnp
2.4. Expression and Purification of YtnP
2.5. Bioassay of AHL-Lactonase Activity
2.6. YtnP Inhibition of A. hydrophila Biofilm Formation
2.7. Determination of Minimum Inhibitory Concentration
2.8. Virulence of A. hydrophila Inhibited with YtnP
2.9. Effect of YtnP against Infection of A. hydrophila Infection
2.10. Nucleotide Sequence Accession Number
3. Statistical Analysis
4. Results
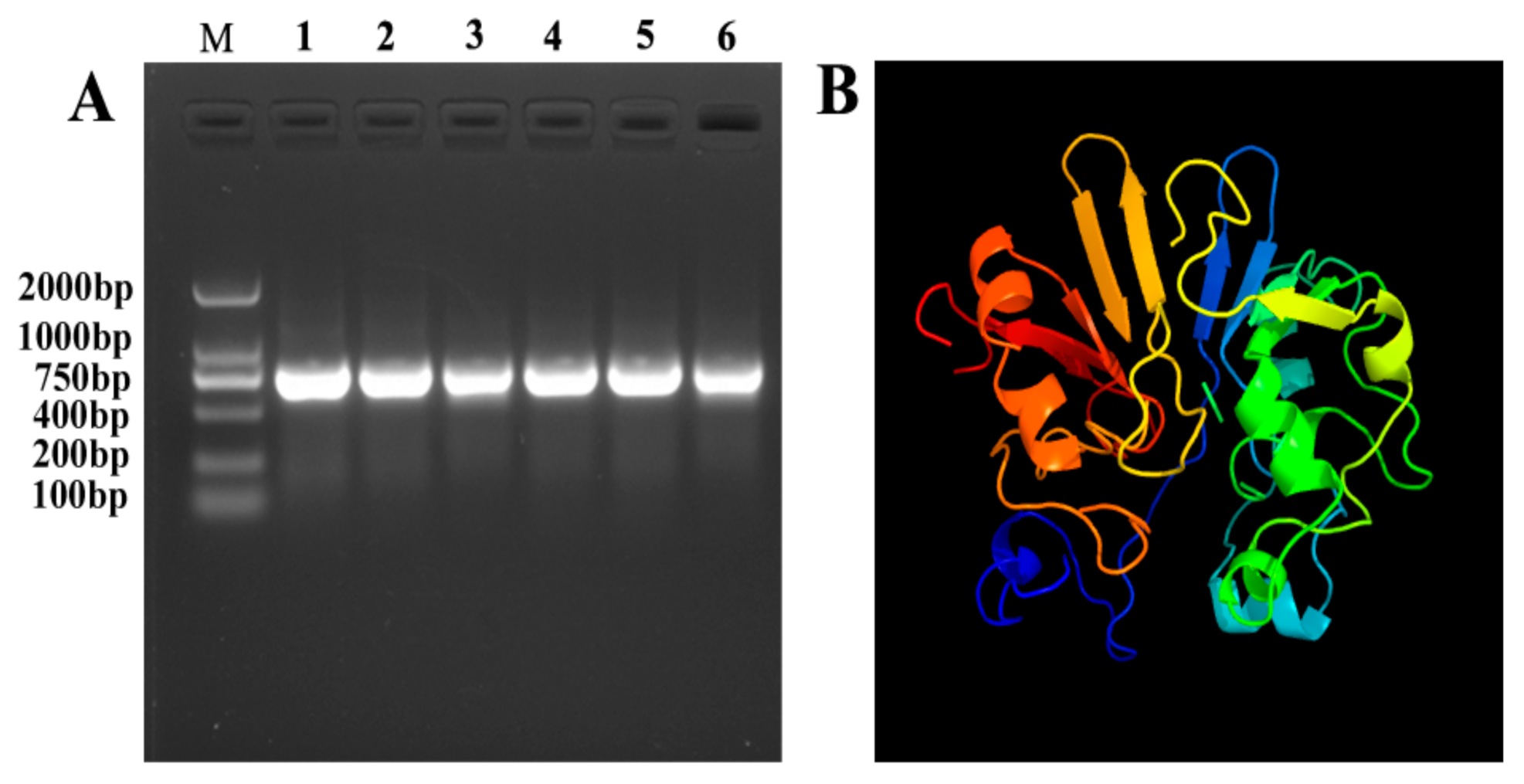
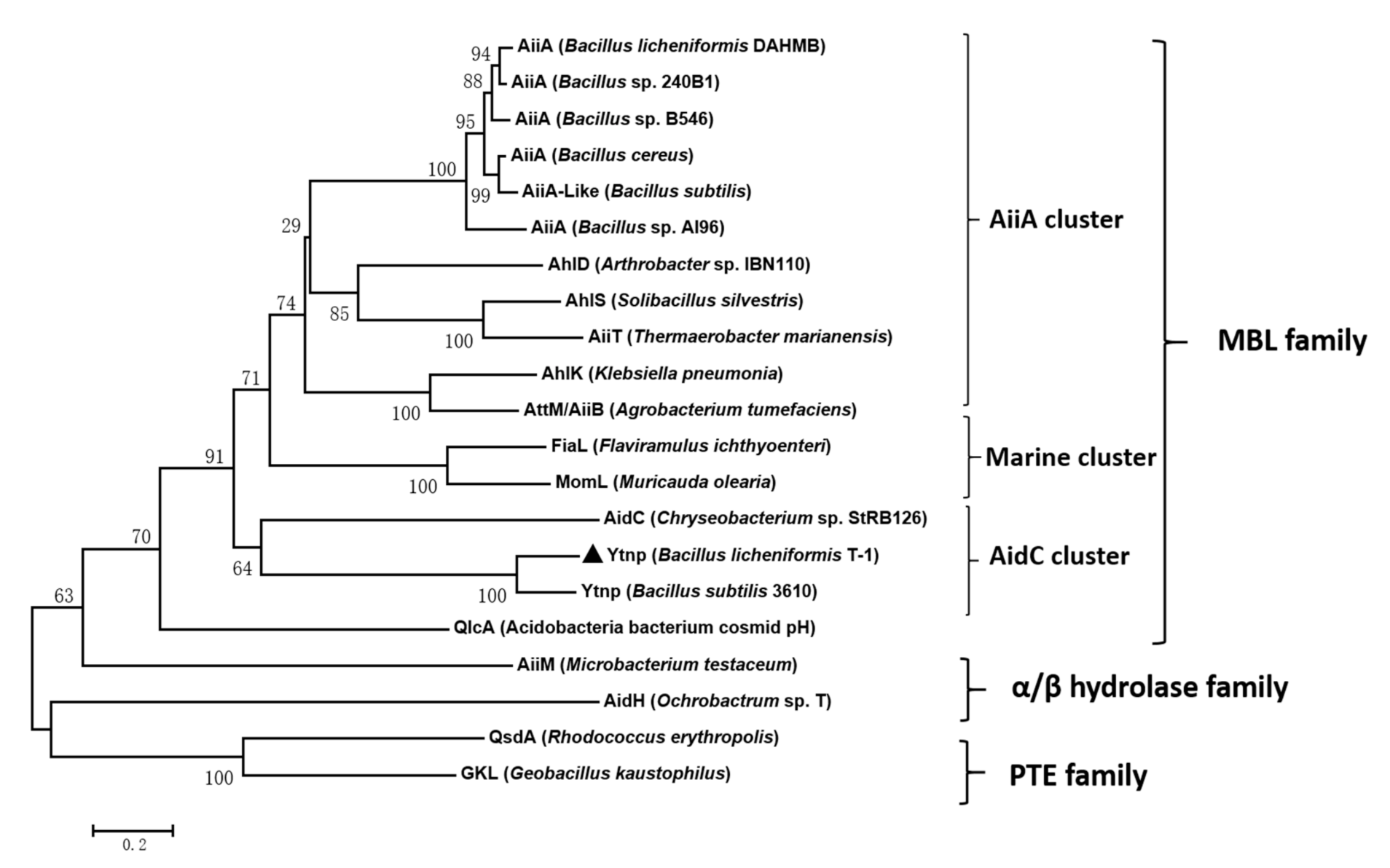
4.1. Gene Clone and Sequence Analysis of ytnp
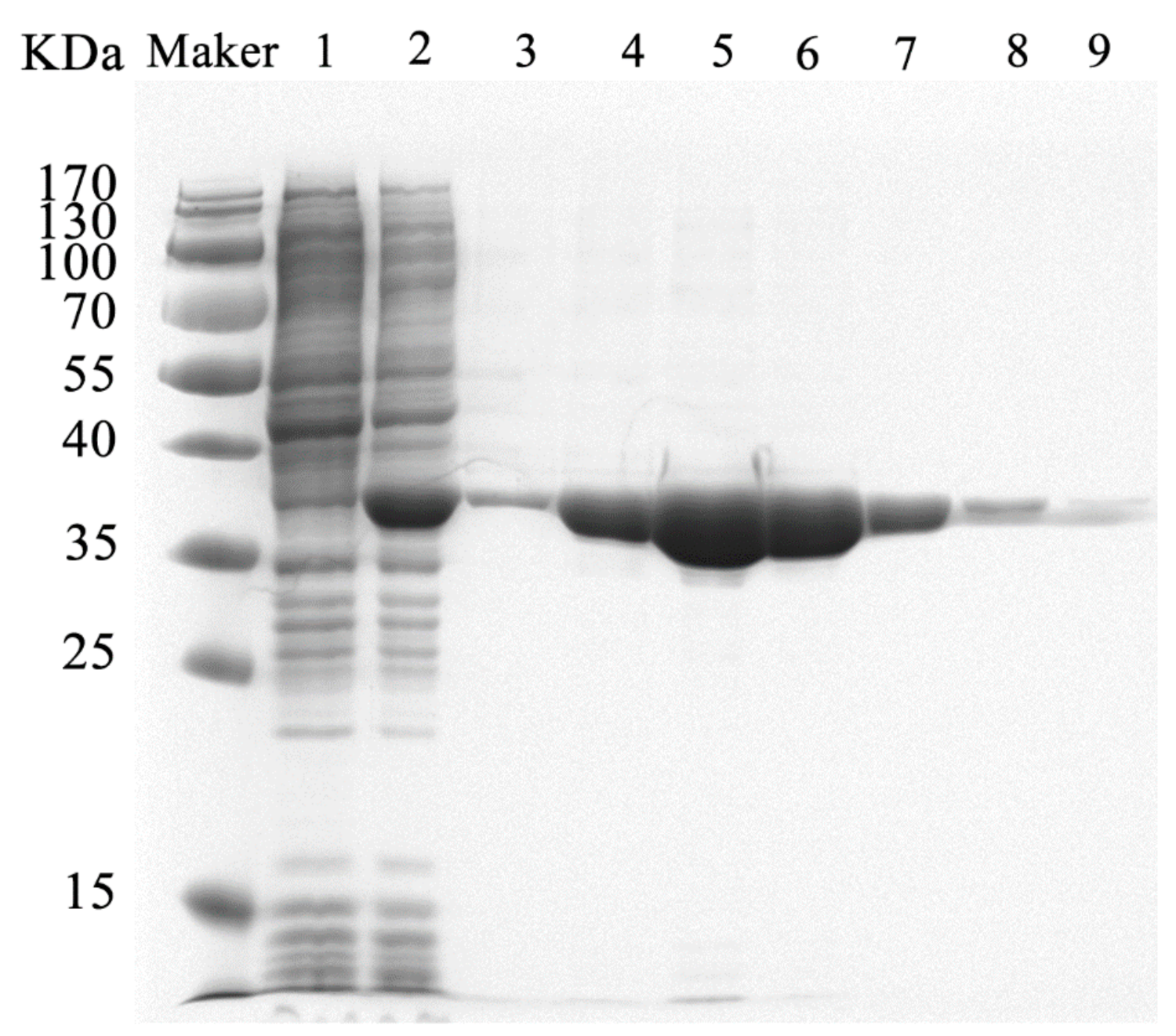
4.2. Expression and Purification of YtnP
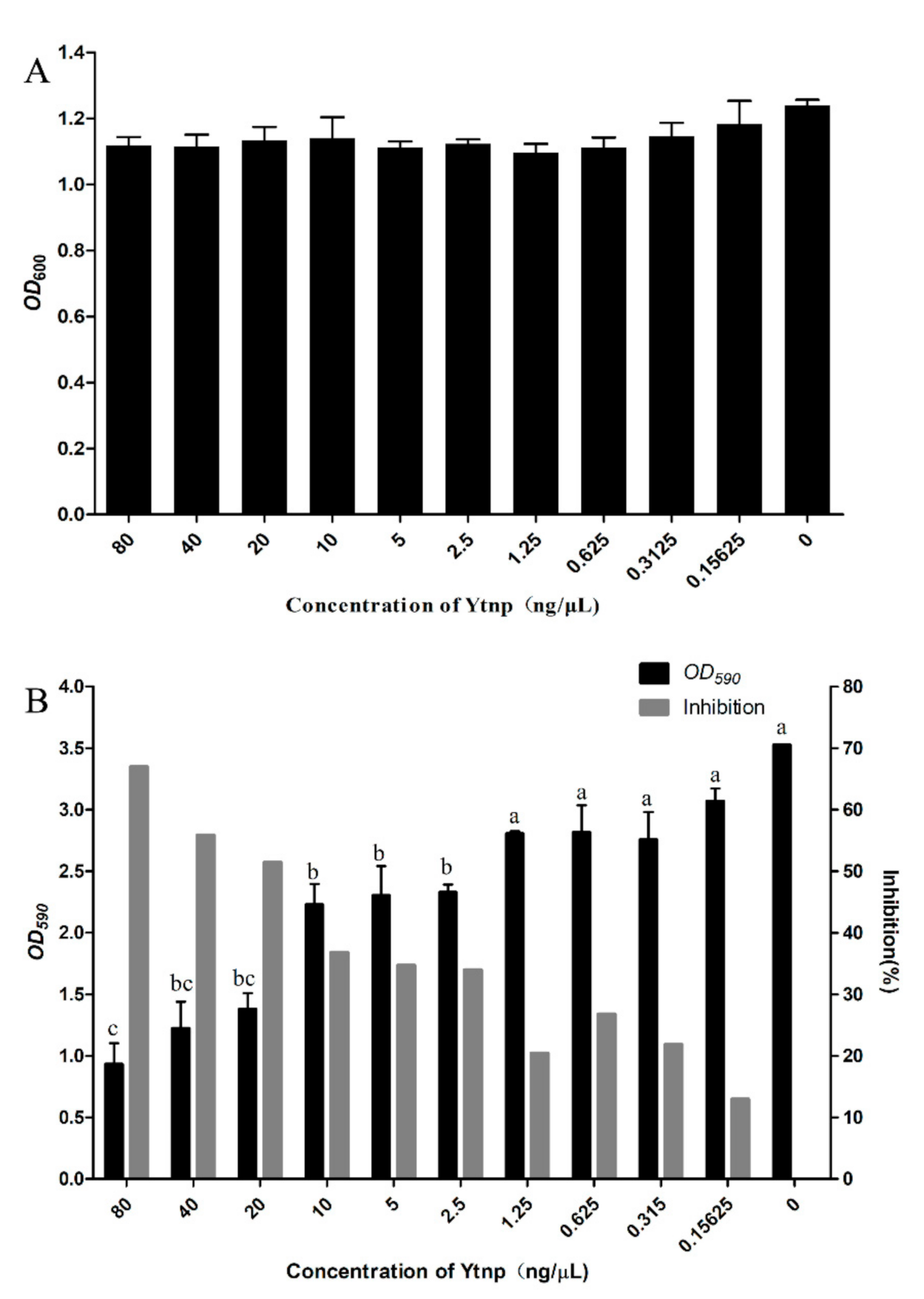
4.3. YtnP Inhibition of A. hydrophila Biofilm Formation
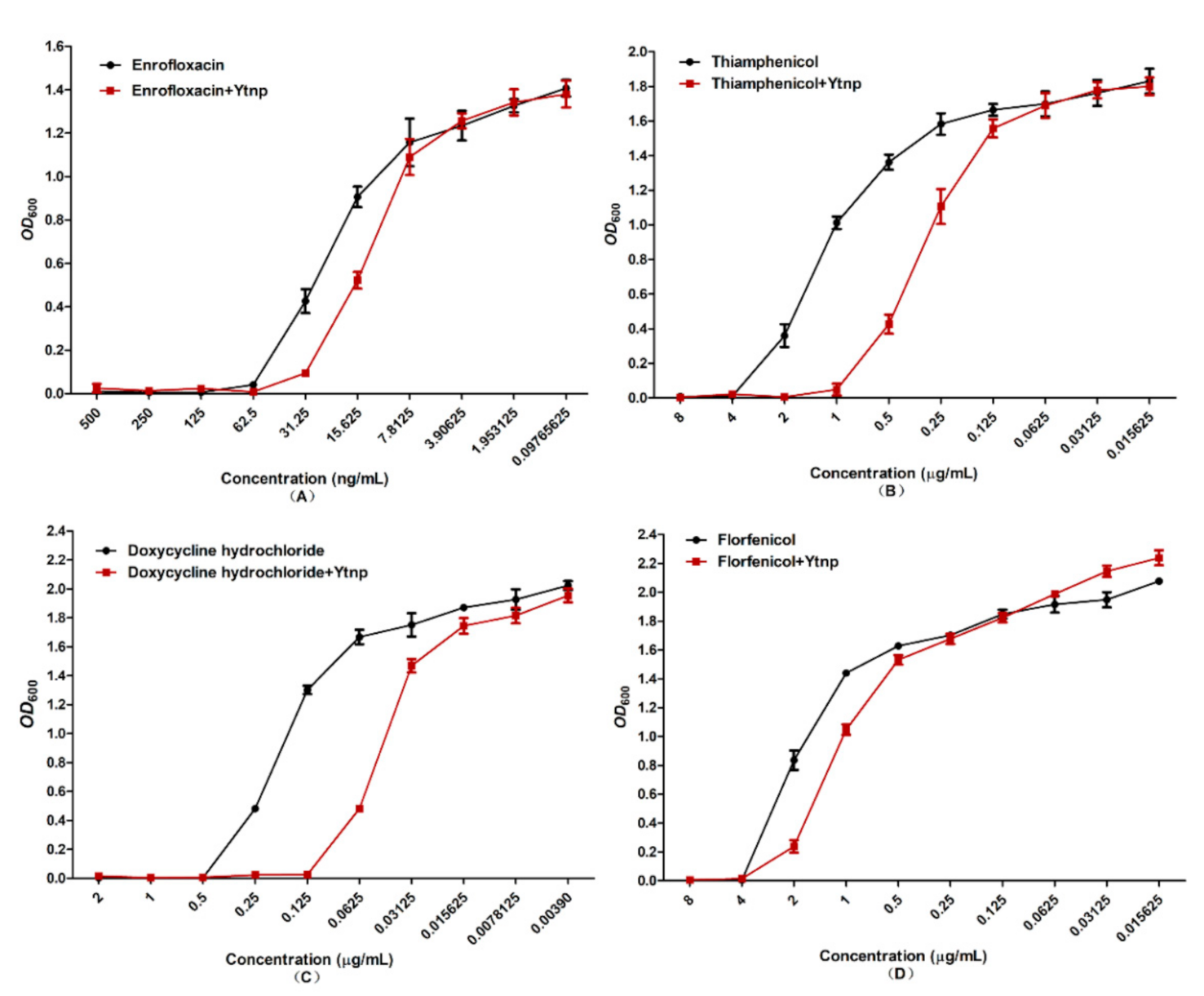
4.4. Synergistic Effect of YtnP and Antibiotics on MIC of A. hydrophila
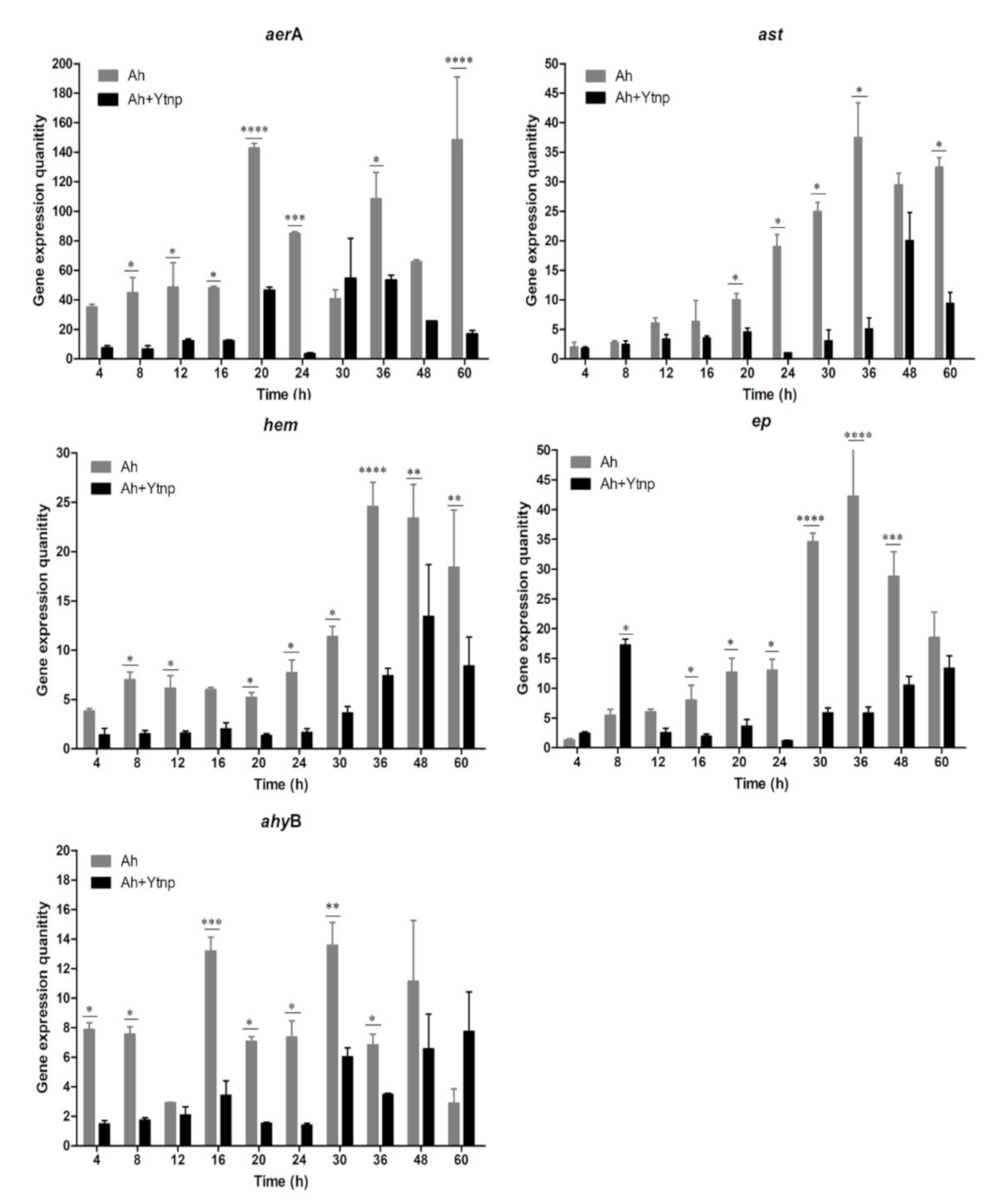
4.5. Virulence Gene of A. hydrophila Inhibited with YtnP
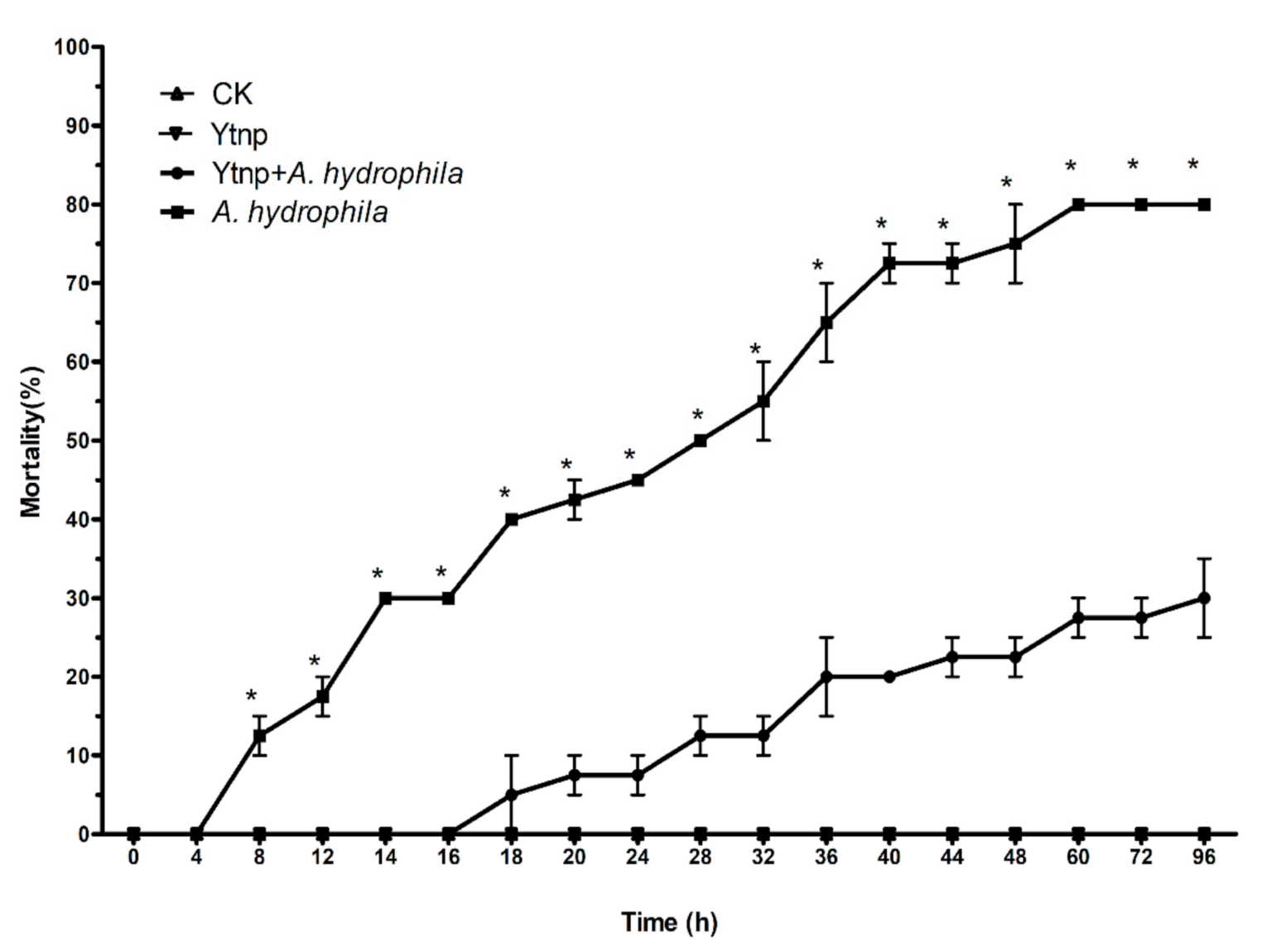
4.6. Co-Injection of Recombinant YtnP and A. hydrophila in C. auratus
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, Y.H.; Wang, L.Y.; Zhang, L.H. Quorum-quenching microbial infections: Mechanisms and implications. Philos. Trans. R. Soc. Lond. 2007, 362, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Zhang, L.H. Quorum sensing and quorum-quenching enzymes. J. Microbiol. 2005, 43, 101–109. [Google Scholar]
- Jiang, G.; Su, M. Retracted article: Quorum-sensing of bacteria and its application. J. Ocean Univ. China 2009, 8, 385–391. [Google Scholar] [CrossRef]
- Wang, N.; Gao, J.; Liu, Y.; Wang, Q.; Zhuang, X.; Zhuang, G. Realizing the role of N-acyl-homoserine lactone-mediated quorum sensing in nitrification and denitrification: A review. Chemosphere 2021, 274, 129970. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A new class of homoserine lactone quorum-sensing signals. Nature 2016, 454, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, X.H. Quorum Quenching Agents: Resources for Antivirulence Therapy. Mar. Drugs 2014, 12, 3245–3282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.; Azevedo, V.; Bertero, M.G.; Bessières, P.; Bolotin, A.; Borchert, S.; et al. The complete genome sequence of the Gram-positive bacterium Bacillus subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macek, B.; Mijakovic, I.; Olsen, J.V.; Gnad, F.; Kumar, C.; Jensen, P.R.; Mann, M. The serine/threonine/tyrosine phosphoproteome of the model bacterium Bacillus subtilis. Mol. Cell. Proteom. 2007, 6, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.; Yepes, A.; Garcia-Betancur, J.C.; Westedt, I.; Mielich, B.; Lopez, D. Streptomycin-induced expression in Bacillus subtilis of YtnP, a lactonase-homologous protein that inhibits development and streptomycin production in Streptomyces griseus. Appl. Environ. Microbiol. 2012, 78, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Morohoshi, T.; Kato, M.; Fukamachi, K.; Kato, N.; Ikeda, T. N-Acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 2008, 279, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, N.; Borde, A.; Wölfel, S.; Kerman, B.; Larsson, A. Quantification of protein concentration by the Bradford method in the presence of pharmaceutical polymers. Anal. Biochem. 2011, 411, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Saeki, E.K.; Kobayashi, R.K.T.; Nakazato, G. Quorum sensing system: Target to control the spread of bacterial infections. Microb. Pathog. 2020, 142, 104068. [Google Scholar] [CrossRef]
- Christensen, L.D.; Gennip, M.V.; Jakobsen, T.H.; Givskov, M.; Bjarnsholt, T. Imaging N-acyl homoserine lactone quorum sensing in vivo. Methods Mol. Biol. 2011, 692, 147–157. [Google Scholar]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Brackman, G.; Coenye, T. Quorum sensing inhibitors as anti-biofilm agents. Curr. Pharm. Des. 2015, 21, 5–11. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.H.; Byun, Y.; Park, H.D. 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci Rep 2015, 5, 551–558. [Google Scholar] [CrossRef]
- Goo, E.; An, J.H.; Kang, Y.; Hwang, I. Control of bacterial metabolism by quorum sensing. Trends Microbiol. 2015, 23, 567–576. [Google Scholar] [CrossRef]
- Khajanchi, B.K.; Sha, J.; Kozlova, E.V.; Erova, T.E.; Suarez, G.; Sierra, J.C.; Popov, V.L.; Horneman, A.J.; Chopra, A.K. N-Acylhomoserine lactones involved in quorum sensing control the type VI secretion system, biofilm formation, protease production, and in vivo virulence in a clinical isolate of Aeromonas hydrophila. Microbiology 2009, 155, 3518–3531. [Google Scholar] [CrossRef] [Green Version]
- Natrah, F.M.I.; Alam, M.I.; Pawar, S.; Harzevili, A.S.; Nevejan, N.; Boon, N.; Sorgeloos, P.; Bossier, P.; Defoirdtac, T. The impact of quorum sensing on the virulence of Aeromonas hydrophila, and Aeromonas salmonicida, towards burbot (Lota lota, L.) larvae. Vet. Microbiol. 2012, 159, 77–82. [Google Scholar]
- Junker, L.M.; Clardy, J. High-throughput screens for small-molecule inhibitors of Pseudomonas aeruginosa biofilm development. Antimicrob. Agents Chemother. 2007, 51, 3582. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Peng, M.F.; Tong, W.T.; Zhang, Q.H.; Song, Z.F. The quorum quenching bacterium Bacillus licheniformis T-1 protects zebrafish against Aeromonas hydrophila infection, Probiotics and Antimicrobial Proteins. Probiotics Antimicrob. Proteins 2020, 12, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Shuxin, Z.; An, Z.; Hongping, Y.; Weihua, C. Bacillus sp. QSI-1 Modulate Quorum Sensing Signals Reduce Aeromonas hydrophila Level and Alter Gut Microbial Community Structure in Fish. Front. Cell. Infect. Microbiol. 2016, 6, 184. [Google Scholar]
- Lee, S.J.; Park, S.Y.; Lee, J.J.; Yum, D.Y.; Koo, B.T.; Lee, J.K. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl. Environ. Microbiol. 2002, 68, 3919–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, W.; Zhou, S.; Zhu, W.; Zhuang, X. Quorum quenching bacteria Bacillus sp. QSI-1 protect zebrafish (Danio rerio) from Aeromonas hydrophila infection. Sci. Rep. 2014, 4, 5446. [Google Scholar] [CrossRef] [Green Version]
- Chen, B. Study on Mechanism to Inhibit Quorum Sensing of Bacillus licheniformis T-1 and Its Biological Characteristics; Shanghai Ocean University: Shanghai, China, 2015. [Google Scholar]
- Vinoj, G.; Vaseeharan, B.; Thomas, S.; Spiers, A.J.; Shanthi, S. Quorum-Quenching Activity of the AHL-Lactonase from Bacillus licheniformis DAHB1 Inhibits Vibrio Biofilm Formation In Vitro and Reduces Shrimp Intestinal Colonisation and Mortality. Mar. Biotechnol. 2014, 16, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Bai, F.; Han, Y.; Chen, J.; Zhang, X.H. Disruption of quorum sensing in Vibrio harveyi, by the AiiA protein of Bacillus thuringiensis. Aquaculture 2008, 274, 36–40. [Google Scholar] [CrossRef]
- Pan, J.; Huang, T.; Yao, F.; Huang, Z.; Powell, C.A.; Qiu, S.; Guan, X. Expression and characterization of aiiA gene from Bacillus subtilis BS-1. Microbiol. Res. 2008, 163, 711–716. [Google Scholar]
- Jayalekshmi, H.; Omanakuttan, A.; Pandurangan, N.; Vargis, V.S.; Maneesh, M.; Nair, B.G.; Kumar, G.B. Clove bud oil reduces kynurenine and inhibits pqs A gene expression in P. aeruginosa. Appl. Microbiol. Biotechnol. 2016, 100, 3681–3692. [Google Scholar] [CrossRef]
- Adriana, P.R.; Pinto, U.M.; Andrea, D.O.B.R.; Bazzolli, D.M.S.; Vanetti, M.C.D. Quorum sensing regulated phenotypes in Aeromonas hydrophila, ATCC 7966 deficient in AHL production. Ann. Microbiol. 2016, 66, 1117–1126. [Google Scholar]
- Garde, C.; Bjarnsholt, T.; Givskov, M.; Jakobsen, T.H.; Hentzer, M.; Claussen, A.; Sneppen, K.; Ferkinghoff-Borg, J.; Sams, T. Quorum sensing regulation in Aeromonas hydrophila. J. Mol. Biol. 2010, 396, 849–857. [Google Scholar] [CrossRef]
- Lynch, M.J.; Swift, S.; Kirke, D.F.; Keevil, C.W.; Dodd, C.E.; Williams, P. The regulation of biofilm development by quorum sensing in Aeromonas hydrophila. Environ. Microbiol. 2010, 4, 18–28. [Google Scholar] [CrossRef]
- Li, J.; Ni, X.D.; Liu, Y.J.; Lu, C.P. Detection of three virulence genes alt, ahp and aerA in Aeromonas hydrophila and their relationship with actual virulence to zebrafish. J. Appl. Microbiol. 2011, 110, 823–830. [Google Scholar] [CrossRef]
- Brackman, G.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob. Agents Chemother. 2011, 55, 2655. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; He, S.; Zhou, Z.; Zhang, M.; Mao, W.; Zhang, H.; Yao, B. Orally Administered Thermostable N-Acyl Homoserine Lactonase from Bacillus sp. Strain AI96 Attenuates Aeromonas hydrophila Infection in Zebrafish. Appl. Environ. Microbiol. 2012, 78, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Zhou, Z.; Cao, Y.; Bai, Y.; Yao, B. High yield expression of an AHL-lactonase from Bacillus sp. B546 in Pichia pastoris and its application to reduce Aeromonas hydrophila mortality in aquaculture. Microb. Cell Factories 2010, 9, 39. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Fragment Size |
|---|---|---|
| B-subumit | B-subumitF:gtgcgtgaaggtctgattgccgt B-subumitR:gcttctcacccatcgcctgttcg | 160 |
| ast | Ast-F:ccccgcctggctgttcttt Ast-R:cggcgaagtcttgcggtga | 166 |
| aerA | AerA-F:cctgagcctgtctgaccaagt AerA-R:gctcgggtcgaagttctcg | 192 |
| hem | hem-F:gtcatgacctgacgctgag hem-R:ctggtaacgaatgctgctc | 137 |
| ep | Ep-F:tcccattgccctgttgct Ep-R:tcgtcactgttgccatcca | 122 |
| ahyB | AhyB-F:gccgctgaatccctcctcctac AhyB-R:tgctgccgacgttgttcttgtag | 156 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, M.; Tong, W.; Zhao, Z.; Xiao, L.; Wang, Z.; Liu, X.; He, X.; Song, Z. Attenuation of Aeromonas hydrophila Infection in Carassius auratus by YtnP, a N-acyl Homoserine Lactonase from Bacillus licheniformis T-1. Antibiotics 2021, 10, 631. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060631
Peng M, Tong W, Zhao Z, Xiao L, Wang Z, Liu X, He X, Song Z. Attenuation of Aeromonas hydrophila Infection in Carassius auratus by YtnP, a N-acyl Homoserine Lactonase from Bacillus licheniformis T-1. Antibiotics. 2021; 10(6):631. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060631
Chicago/Turabian StylePeng, Mengfan, Wentao Tong, Zhen Zhao, Ling Xiao, Zhaoyue Wang, Xuanming Liu, Xuanhao He, and Zengfu Song. 2021. "Attenuation of Aeromonas hydrophila Infection in Carassius auratus by YtnP, a N-acyl Homoserine Lactonase from Bacillus licheniformis T-1" Antibiotics 10, no. 6: 631. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060631