
Gram-Positive Pneumonia: Possibilities Offered by Phage Therapy

 , ,
, ,  and
and 
Abstract
:1. Introduction
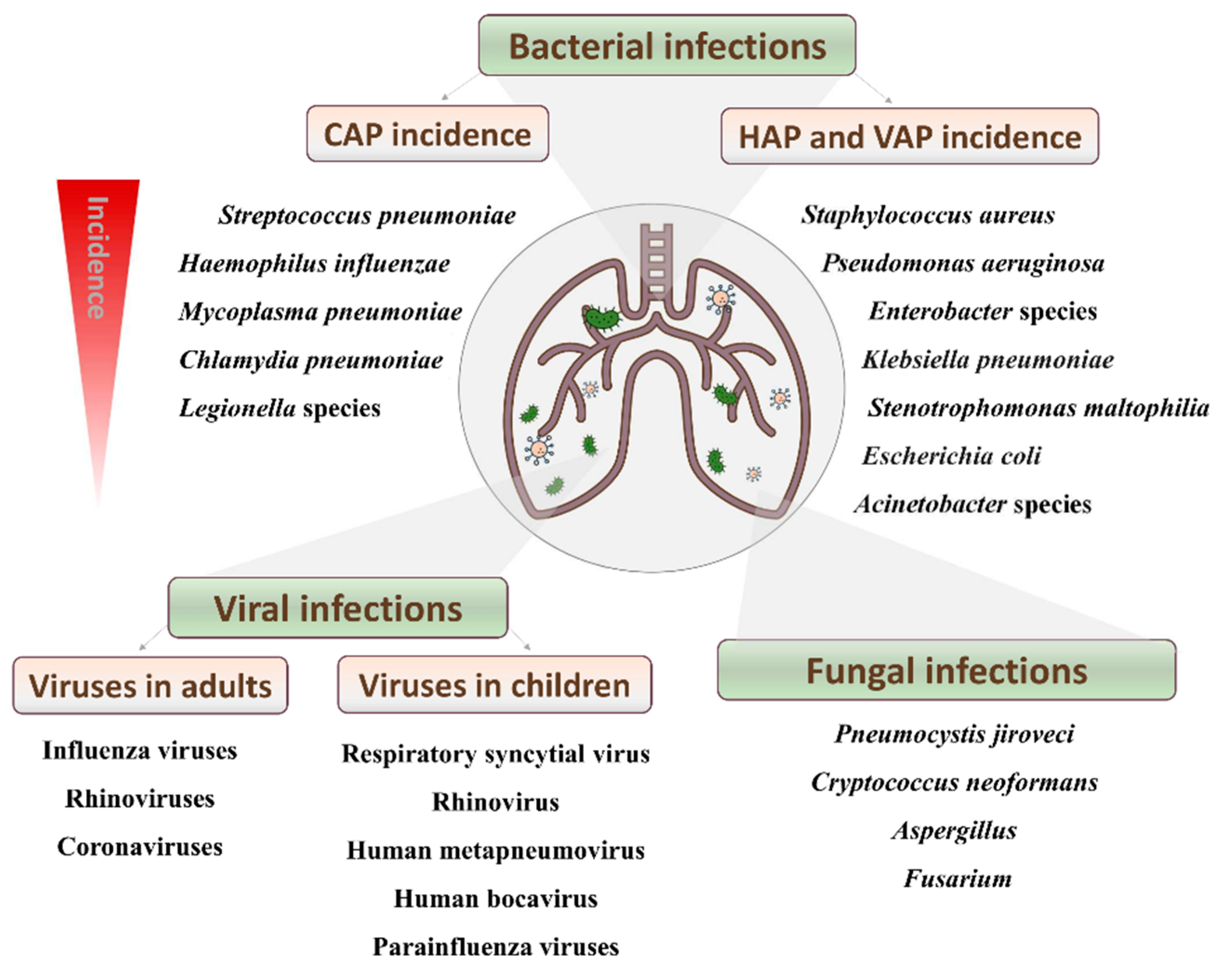
2. Disease Incidence and Mortality
3. Diagnosis of Bacterial Pneumonia
4. Preventive and Therapeutic Strategies against Gram-Positive Pneumonia
4.1. Pneumonia Prevention: Impact of Vaccination
4.2. Present and Future Therapeutic Strategies
5. Phage Therapy
5.1. Bacteriophages as New Weapons against Pneumonia-Causing Bacteria
5.2. Phage-Derived Lytic Proteins
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henig, O.; Kaye, K.S. Bacterial Pneumonia in Older Adults. Infect. Dis. Clin. N. Am. 2017, 31, 689–713. [Google Scholar] [CrossRef]
- Troeger, C.; Blacker, B.; Khalil, I.; Rao, P.C.; Cao, J.; Zimsen, S.R.M.; Albertson, S.B.; Deshpande, A.; Farag, T.; Abebe, Z.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Ruuskanen, O.; Lahti, E.; Jennings, L.C.; Murdoch, D.R. Viral pneumonia. Lancet 2011, 377, 1264–1275. [Google Scholar] [CrossRef]
- Jose, R.J.P.; Brown, J.S. Opportunistic and fungal infections of the lung. Medicine 2012, 40, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Prina, E.; Ranzani, O.; Torres, A. Community-acquired pneumonia. Lancet 2015, 386, 1097–1108. [Google Scholar] [CrossRef]
- Jain, V.; Vashisht, R.; Yilmaz, G.; Bhardwaj, A. Pneumonia Pathology; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Gonzalez-Zorn, B. Antibiotic use in the COVID-19 crisis in Spain. Clin. Microbiol. Infect. 2020, 27, 646–647. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Goodarzi, P.; Asadi, M.; Soltani, A.; Aljanabi, H.A.A.; Jeda, A.S.; Dashtbin, S.; Jalalifar, S.; Mohammadzadeh, R.; Teimoori, A.; et al. Bacterial co-infections with SARS-CoV-2. IUBMB Life 2020, 72, 2097–2111. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Fact Sheet—Pneumonia. 2019. Available online: http://www.who.int/mediacentre/factsheets/fs331/en/ (accessed on 15 January 2021).
- Iannella, H.A.; Luna, C.M. Community-Acquired Pneumonia in Latin America. Semin. Respir. Crit. Care Med. 2016, 37, 868–875. [Google Scholar] [CrossRef]
- Vallés, J.; Pobo, A.; Garcia-Esquirol, O.; Mariscal, D.; Real, J.; Fernandez, R. Excess ICU mortality attributable to ventilator-associated pneumonia: The role of early vs late onset. Intensiv. Care Med. 2007, 33, 1363–1368. [Google Scholar] [CrossRef]
- Feldman, C.; Anderson, R. Epidemiology, virulence factors and management of the pneumococcus. F1000Research 2016, 5, 2320. [Google Scholar] [CrossRef]
- Liese, J.G.; Schoen, C.; Van Der Linden, M.; Lehmann, L.; Goettler, D.; Keller, S.; Maier, A.; Segerer, F.; Rose, M.A.; Streng, A. Changes in the incidence and bacterial aetiology of paediatric parapneumonic pleural effusions/empyema in Germany, 2010–2017: A nationwide surveillance study. Clin. Microbiol. Infect. 2018, 25, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.N. Microbial Etiologies of Hospital-Acquired Bacterial Pneumonia and Ventilator-Associated Bacterial Pneumonia. Clin. Infect. Dis. 2010, 51, S81–S87. [Google Scholar] [CrossRef] [Green Version]
- Self, W.H.; Wunderink, R.G.; Williams, D.J.; Zhu, Y.; Anderson, E.J.; Balk, R.A.; Fakhran, S.S.; Chappell, J.D.; Casimir, G.; Courtney, D.M.; et al. Staphylococcus aureus Community-acquired Pneumonia: Prevalence, Clinical Characteristics, and Outcomes. Clin. Infect. Dis. 2016, 63, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Hyams, C.; Williams, O.M.; Williams, P. Urinary antigen testing for pneumococcal pneumonia: Is there evidence to make its use uncommon in clinical practice? ERJ Open Res. 2020, 6, 00223-2019. [Google Scholar] [CrossRef]
- Cima, D.; Méndez, F.J.; Vázquez, F.J.; Aranaz, C.; Álvarez, J.R.R.; García-García, J.-M.; Fleites, A.; González, J.R.D.L.T.; Molinos, L.; De Miguel, D.; et al. Immunodetection of pneumolysin in human urine by ELISA. J. Microbiol. Methods 2003, 54, 47–55. [Google Scholar] [CrossRef]
- García-Suárez, M.D.M.; Cima-Cabal, M.D.; Villaverde, R.; Espinosa, E.; Falguera, M.; Toyos, J.R.D.L.; Vázquez, F.; Méndez, F.J. Performance of a Pneumolysin Enzyme-Linked Immunosorbent Assay for Diagnosis of Pneumococcal Infections. J. Clin. Microbiol. 2007, 45, 3549–3554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murdoch, D.R. Molecular genetic methods in the diagnosis of lower respiratory tract infections. APMIS 2004, 112, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, A.; Lindh, M.; Westin, J.; Andersson, L.-M.; Baldursson, O.; Kristinsson, K.G.; Gottfredsson, M. Utility of oropharyngeal real-time PCR for S. pneumoniae and H. influenzae for diagnosis of pneumonia in adults. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 36, 529–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duke, T. What the PERCH study means for future pneumonia strategies. Lancet 2019, 394, 714–716. [Google Scholar] [CrossRef] [Green Version]
- I A Mustafa, M.; Al-Marzooq, F.; How, S.H.; Kuan, Y.C.; Ng, T.H. The use of multiplex real-time PCR improves the detection of the bacterial etiology of community acquired pneumonia. Trop. Biomed. 2011, 28. [Google Scholar]
- Gowin, E.; Bartkowska-Śniatkowska, A.; Jończyk-Potoczna, K.; Wysocka-Leszczyńska, J.; Bobkowski, W.; Fichna, P.; Sobkowiak, P.; Mazur-Melewska, K.; Bręborowicz, A.; Wysocki, J.; et al. Assessment of the Usefulness of Multiplex Real-Time PCR Tests in the Diagnostic and Therapeutic Process of Pneumonia in Hospitalized Children: A Single-Center Experience. BioMed Res. Int. 2017, 2017, 8037963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Ruan, S.-Y.; Pan, S.-C.; Lee, T.-F.; Chien, J.-Y.; Hsueh, P.-R. Performance of a multiplex PCR pneumonia panel for the identification of respiratory pathogens and the main determinants of resistance from the lower respiratory tract specimens of adult patients in intensive care units. J. Microbiol. Immunol. Infect. 2019, 52, 920–928. [Google Scholar] [CrossRef]
- Tomita, N.; Mori, Y.; Kanda, H.; Notomi, T. Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nat. Protoc. 2008, 3, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.-H.; Dai, Y.-Y.; Huang, L.-Y.; Zhang, W.-S.; Guo, X.-G. Diagnosis of mycoplasma pneumoniae by loop-mediated isothermal amplification: Systematic review and meta-analysis. BMC Infect. Dis. 2019, 19, 173. [Google Scholar] [CrossRef] [Green Version]
- Cima-Cabal, M.D.; Vázquez-Espinosa, E.; Vazquez, F.; García-Suárez, M.D.M. Detection of Streptococcus pneumoniae in Urine by Loop-Mediated Isothermal Amplification. J. Pediatr. Infect. Dis. 2020, 16, 018–025. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Z.; Bao, Y.; Yang, Y.; De Groot, R.; Dai, W.; De Jonge, M.I.; Zheng, Y. Clinical diagnostic application of metagenomic next-generation sequencing in children with severe nonresponding pneumonia. PLoS ONE 2020, 15, e0232610. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Cima-Cabal, M.D.; Duarte, A.C.; Rodriguez, A.; García, P.; García-Suárez, M.D.M. Developing Diagnostic and Therapeutic Approaches to Bacterial Infections for a New Era: Implications of Globalization. Antibiotics 2020, 9, 916. [Google Scholar] [CrossRef]
- Wang, H.; Ma, Z.; Qin, J.; Shen, Z.; Liu, Q.; Chen, X.; Wang, H.; An, Z.; Liu, W.; Li, M. A versatile loop-mediated isothermal amplification microchip platform for Streptococcus pneumoniae and Mycoplasma pneumoniae testing at the point of care. Biosens. Bioelectron. 2018, 126, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.P.; Zhang, W.; Lin, S.; Thurston, S.W.; Hopke, P.K.; Masiol, M.; Squizzato, S.; Van Wijngaarden, E.; Utell, M.J.; Rich, D.Q. The Association between Respiratory Infection and Air Pollution in the Setting of Air Quality Policy and Economic Change. Ann. Am. Thorac. Soc. 2018, 16, 321–330. [Google Scholar] [CrossRef]
- Grabenstein, J.D.; Klugman, K.P. A century of pneumococcal vaccination research in humans. Clin. Microbiol. Infect. 2012, 18, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Redi, D.; Raffaelli, C.S.; Rossetti, B.; De Luca, A.; Montagnani, F. Staphylococcus aureus vaccine preclinical and clinical development: Current state of the art. New Microbiol. 2018, 41, 208–213. [Google Scholar]
- Woodhead, M.; Blasi, F.; Ewig, S.; Garau, J.; Huchon, G.; Ieven, M.; Ortqvist, A.; Schaberg, T.; Torres, A.; van der Heijden, G.; et al. Guidelines for the management of adult lower respiratory tract infections—Full version. Clin. Microbiol. Infect. 2011, 17, E1–E59. [Google Scholar] [CrossRef] [Green Version]
- Pränting, M.; Negrea, A.; Rhen, M.; Andersson, D.I. Mechanism and Fitness Costs of PR-39 Resistance in Salmonella enterica Serovar Typhimurium LT2. Antimicrob. Agents Chemother. 2008, 52, 2734–2741. [Google Scholar] [CrossRef] [Green Version]
- Banaschewski, B.J.H.; Veldhuizen, E.J.A.; Keating, E.; Haagsman, H.P.; Zuo, Y.Y.; Yamashita, C.; Veldhuizen, R.A.W. Antimicrobial and Biophysical Properties of Surfactant Supplemented with an Antimicrobial Peptide for Treatment of Bacterial Pneumonia. Antimicrob. Agents Chemother. 2015, 59, 3075–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, R.; Surendran, N.; Ochs, M.; Pichichero, M.E. Human Antibodies to PhtD, PcpA, and Ply Reduce Adherence to Human Lung Epithelial Cells and Murine Nasopharyngeal Colonization by Streptococcus pneumoniae. Infect. Immun. 2014, 82, 5069–5075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, B.; Mercier, E.; Gonzalez, C.; Asehnoune, K.; Nseir, S.; Fiancette, M.; Desachy, A.; Plantefève, G.; Meziani, F.; de Lame, P.-A.; et al. Safety and tolerability of a single administration of AR-301, a human monoclonal antibody, in ICU patients with severe pneumonia caused by Staphylococcus aureus: First-in-human trial. Intensiv. Care Med. 2018, 44, 1787–1796. [Google Scholar] [CrossRef]
- Hua, L.; Hilliard, J.J.; Shi, Y.; Tkaczyk, C.; Cheng, L.I.; Yu, X.; Datta, V.; Ren, S.; Feng, H.; Zinsou, R.; et al. Assessment of an Anti-Alpha-Toxin Monoclonal Antibody for Prevention and Treatment of Staphylococcus aureus-Induced Pneumonia. Antimicrob. Agents Chemother. 2013, 58, 1108–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foletti, D.; Strop, P.; Shaughnessy, L.; Hasa-Moreno, A.; Casas, M.G.; Russell, M.; Bee, C.; Wu, S.; Pham, A.; Zeng, Z.; et al. Mechanism of Action and In Vivo Efficacy of a Human-Derived Antibody against Staphylococcus aureus α-Hemolysin. J. Mol. Biol. 2013, 425, 1641–1654. [Google Scholar] [CrossRef]
- Rouha, H.; Weber, S.; Janesch, P.; Maierhofer, B.; Gross, K.; Dolezilkova, I.; Mirkina, I.; Visram, Z.C.; Malafa, S.; Stulik, L.; et al. Disarming Staphylococcus aureus from destroying human cells by simultaneously neutralizing six cytotoxins with two human monoclonal antibodies. Virulence 2017, 9, 231–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, B.; Yeh, A.J.; Cheung, G.Y.C.; Otto, M. Investigational therapies targeting quorum-sensing for the treatment of Staphylococcus aureus infections. Expert Opin. Investig. Drugs 2015, 24, 689–704. [Google Scholar] [CrossRef]
- Henry, B.D.; Neill, D.; Becker, K.A.; Gore, S.; Bricio-Moreno, L.; Ziobro, R.; Edwards, M.J.; Mühlemann, K.; Steinmann, J.; Kleuser, B.; et al. Engineered liposomes sequester bacterial exotoxins and protect from severe invasive infections in mice. Nat. Biotechnol. 2014, 33, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, X.; Liu, S.; Li, G.; Shi, L.; Dong, J.; Li, W.; Deng, X.; Niu, X. Morin hydrate attenuates Staphylococcus aureus virulence by inhibiting the self-assembly of α-hemolysin. J. Appl. Microbiol. 2015, 118, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, G.R.; DeDent, A.C.; Becker, R.E.N.; Berube, B.J.; Gebhardt, M.J.; Cao, H.; Wardenburg, J.B. Targeting Staphylococcus aureus α-toxin as a novel approach to reduce severity of recurrent skin and soft-tissue infections. J. Infect. Dis. 2014, 210, 1012–1018. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Nijnik, A.; Philpott, D.J. Modulating immunity as a therapy for bacterial infections. Nat. Rev. Genet. 2012, 10, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Gutiérrez, D.; García, P.; Rodríguez, A. The Perfect Bacteriophage for Therapeutic Applications—A Quick Guide. Antibiotics 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, R. Streptococcus pneumoniae and its bacteriophages: One long argument. Int. Microbiol. 2004, 7, 163–171. [Google Scholar] [PubMed]
- Oduor, J.M.O.; Onkoba, N.; Maloba, F.; Nyachieo, A. Experimental phage therapy against haematogenous multi-drug resistant Staphylococcus aureus pneumonia in mice. Afr. J. Lab. Med. 2016, 5, 1–7. [Google Scholar] [CrossRef]
- Prazak, J.; Iten, M.; Cameron, D.; Save, J.; Grandgirard, D.; Resch, G.; Goepfert, C.; Leib, S.L.; Takala, J.; Jakob, S.M.; et al. Bacteriophages Improve Outcomes in Experimental Staphylococcus aureus Ventilator-associated Pneumonia. Am. J. Respir. Crit. Care Med. 2019, 200, 1126–1133. [Google Scholar] [CrossRef]
- Prazak, J.; Valente, L.; Iten, M.; Grandgirard, D.; Leib, S.L.; Jakob, S.M.; Haenggi, M.; Que, Y.-A.; Cameron, D.R. Nebulized bacteriophages for prophylaxis of experimental ventilator-associated pneumonia due to methicillin-resistant Staphylococcus aureus. Crit. Care Med. 2020, 48, 1042–1046. [Google Scholar] [CrossRef]
- Prazak, J.; Valente, L.G.; Iten, M.; Federer, L.; Grandgirard, D.; Soto, S.; Resch, G.; Leib, S.L.; Jakob, S.M.; Haenggi, M.; et al. Benefits of Aerosolized Phages for the Treatment of Pneumonia Due to Methicillin-Resistant Staphylococcus aureus: An Experimental Study in Rats. J. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Lehman, S.M.; Mearns, G.; Rankin, D.; Cole, R.A.; Smrekar, F.; Branston, S.D.; Morales, S. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus aureus Infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Kvachadze, L.; Balarjishvili, N.; Meskhi, T.; Tevdoradze, E.; Skhirtladze, N.; Pataridze, T.; Adamia, R.; Topuria, T.; Kutter, E.; Rohde, C.; et al. Evaluation of lytic activity of staphylococcal bacteriophage Sb-1 against freshly isolated clinical pathogens. Microb. Biotechnol. 2011, 4, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Łusiak-Szelachowska, M.; Żaczek, M.; Weber-Dąbrowska, B.; Międzybrodzki, R.; Kłak, M.; Fortuna, W.; Letkiewicz, S.; Rogóż, P.; Szufnarowski, K.; Jończyk-Matysiak, E.; et al. Phage Neutralization by Sera of Patients Receiving Phage Therapy. Viral Immunol. 2014, 27, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, N.; Delattre, R.; Chevallereau, A.; Ricard, J.-D.; Debarbieux, L. Phage Therapy of Pneumonia Is Not Associated with an Overstimulation of the Inflammatory Response Compared to Antibiotic Treatment in Mice. Antimicrob. Agents Chemother. 2019, 63, e00379-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slopek, S.; Weber-Dabrowska, B.; Dabrowski, M.; Kucharewicz-Krukowska, A. Results of bacteriophage treatment of suppurative bacterial infections in the years 1981-1986. Arch. Immunol. Ther. Exp. 1987, 35, 569–583. [Google Scholar]
- Vázquez, R.; García, E.; García, P. Phage Lysins for Fighting Bacterial Respiratory Infections: A New Generation of Antimicrobials. Front. Immunol. 2018, 9, 2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doehn, J.M.; Fischer, K.; Reppe, K.; Gutbier, B.; Tschernig, T.; Hocke, A.C.; Fischetti, V.A.; Löffler, J.; Suttorp, N.; Hippenstiel, S.; et al. Delivery of the endolysin Cpl-1 by in-halation rescues mice with fatal pneumococcal pneumonia. J. Antimicrob. Chemother. 2013, 68, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Vouillamoz, J.; Entenza, J.M.; Giddey, M.; Fischetti, V.A.; Moreillon, P.; Resch, G. Bactericidal synergism between daptomycin and the phage lysin Cpl-1 in a mouse model of pneumococcal bacteraemia. Int. J. Antimicrob. Agents 2013, 42, 416–421. [Google Scholar] [CrossRef]
- Jado, I.; López, R.; García, E.; Fenoll, A.; Casal, J.; García, P. Phage lytic enzymes as therapy for antibiotic-resistant Streptococcus pneumoniae infection in a murine sepsis model. J. Antimicrob. Chemother. 2003, 52, 967–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, N.; Campillo, N.; García, E.; Gallego, C.; Pera, B.; Diakun, G.P.; Sáiz, J.L.; García, P.; Díaz, J.F.; Menéndez, M. Cpl-7, a Lysozyme Encoded by a Pneumococcal Bacteriophage with a Novel Cell Wall-binding Motif. J. Biol. Chem. 2010, 285, 33184–33196. [Google Scholar] [CrossRef] [Green Version]
- Díez-Martínez, R.; De Paz, H.D.; Bustamante, N.; García, E.; Menéndez, M.; García, P. Improving the Lethal Effect of Cpl-7, a Pneumococcal Phage Lysozyme with Broad Bactericidal Activity, by Inverting the Net Charge of Its Cell Wall-Binding Module. Antimicrob. Agents Chemother. 2013, 57, 5355–5365. [Google Scholar] [CrossRef] [Green Version]
- Corsini, B.; Díez-Martínez, R.; Aguinagalde, L.; González-Camacho, F.; García-Fernández, E.; Letrado, P.; García, P.; Yuste, J. Chemotherapy with Phage Lysins Reduces Pneumococcal Colonization of the Respiratory Tract. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez, R.; García, P. Synergy Between Two Chimeric Lysins to Kill Streptococcus pneumoniae. Front. Microbiol. 2019, 10, 1251. [Google Scholar] [CrossRef] [Green Version]
- Letrado, P.; Corsini, B.; Díez-Martínez, R.; Bustamante, N.; Yuste, J.E.; García, P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Luo, D.; Etobayeva, I.; Li, X.; Gong, Y.; Wang, S.; Li, Q.; Xu, P.; Yin, W.; He, J.; et al. Linker Editing of Pneumococcal Lysin ClyJ Conveys Improved Bactericidal Activity. Antimicrob. Agents Chemother. 2020, 64, e01610-19. [Google Scholar] [CrossRef]
- Luo, D.; Huang, L.; Gondil, V.S.; Zhou, W.; Yang, W.; Jia, M.; Hu, S.; He, J.; Yang, H.; Wei, H. A Choline-Recognizing Monomeric Lysin, ClyJ-3m, Shows Elevated Activity against Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2020, 64, e00311-20. [Google Scholar] [CrossRef]
- Van Der Kamp, I.; Draper, L.A.; Smith, M.K.; Buttimer, C.; Ross, R.P.; Hill, C. A New Phage Lysin Isolated from the Oral Microbiome Targeting Streptococcus pneumoniae. Pharmaceuticals 2020, 13, 478. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Fernández, L.; Rodríguez, A.; García, P. Are Phage Lytic Proteins the Secret Weapon to Kill Staphylococcus aureus? mBio 2018, 9, e01923-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, M.; Casey, P.G.; Hill, C.; Gahan, C.; McAuliffe, O.; Mahony, J.O.; Maher, F.; Coffey, A. The truncated phage lysin CHAPk eliminates Staphylococcus aureus in the nares of mice. Bioeng. Bugs 2010, 1, 404–407. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.Y.; Jun, K.I.; Kang, C.K.; Song, K.-H.; Choe, P.G.; Bang, J.-H.; Kim, E.S.; Park, S.W.; Bin Kim, H.; Kim, N.-J.; et al. Efficacy of Intranasal Administration of the Recombinant Endolysin SAL200 in a Lethal Murine Staphylococcus aureus Pneumonia Model. Antimicrob. Agents Chemother. 2019, 63, e02009-18. [Google Scholar] [CrossRef] [Green Version]
- Xia, F.; Li, X.; Wang, B.; Gong, P.; Xiao, F.; Yang, M.; Zhang, L.; Song, J.; Hu, L.; Cheng, M.; et al. Combination Therapy of LysGH15 and Apigenin as a New Strategy for Treating Pneumonia Caused by Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Gondil, V.S.; Dube, T.; Panda, J.J.; Yennamalli, R.M.; Harjai, K.; Chhibber, S. Comprehensive evaluation of chitosan nanoparticle based phage lysin delivery system; a novel approach to counter S. pneumoniae infections. Int. J. Pharm. 2019, 573, 118850. [Google Scholar] [CrossRef]
- Gondil, V.S.; Harjai, K.; Chhibber, S. Investigating the potential of endolysin loaded chitosan nanoparticles in the treatment of pneumococcal pneumonia. J. Drug Deliv. Sci. Technol. 2020, 61, 102142. [Google Scholar] [CrossRef]
- Kaur, J.; Kour, A.; Panda, J.J.; Harjai, K.; Chhibber, S. Exploring Endolysin-Loaded Alginate-Chitosan Nanoparticles as Future Remedy for Staphylococcal Infections. AAPS PharmSciTech 2020, 21, 233. [Google Scholar] [CrossRef]
- Wang, Y.; Khanal, D.; Chang, R.Y.K.; Shang, X.; Yang, H.; Britton, W.J.; Nelson, D.; Chan, H.-K. Can bacteriophage endolysins be nebulised for inhalation delivery against Streptococcus pneumoniae? Int. J. Pharm. 2020, 591, 119982. [Google Scholar] [CrossRef] [PubMed]
- Harhala, M.; Nelson, D.C.; Miernikiewicz, P.; Heselpoth, R.D.; Brzezicka, B.; Majewska, J.; Linden, S.B.; Shang, X.; Szymczak, A.; Lecion, D.; et al. Safety Studies of Pneumococcal Endolysins Cpl-1 and Pal. Viruses 2018, 10, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Górski, A.; Międzybrodzki, R.; Żaczek, M.; Borysowski, J. Phages in the fight against COVID-19? Future Microbiol. 2020, 15, 1095–1100. [Google Scholar] [CrossRef]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; Di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the Host Immune System and Bacteriophage Is Essential for Successful Phage Therapy against an Acute Respiratory Pathogen. Cell Host Microbe 2017, 22, 38–47.e4. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K. Interaction of bacteriophages with the immune system: Induction of bacteriophage-specific Antibodies. Methods Mol. Biol. 2017, 1693, 139–150. [Google Scholar] [CrossRef]
- Bretaudeau, L.; Tremblais, K.; Aubrit, F.; Meichenin, M.; Arnaud, I. Good Manufacturing Practice (GMP) compliance for phage therapy medicinal products. Front. Microbiol. 2020, 11, 1161. [Google Scholar] [CrossRef]
- Starkevič, U.; Bortesi, L.; Virgailis, M.; Ruzauskas, M.; Giritch, A.; Ražanskienė, A. High-yield production of a functional bacteriophage lysin with antipneumococcal activity using a plant virus-based expression system. J. Biotechnol. 2015, 200, 10–16. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Strategies | Effect | Examples | |
|---|---|---|---|
| Antibiotics | Bacterial killing or growth inhibition | Fluoroquinolones | Levofloxacin |
| Moxifloxacin | |||
| β-lactams | Amoxicillin | ||
| Ampicillin | |||
| Amoxicillin-clavulanate | |||
| Ampicillin-sulbactam | |||
| Cefotaxime | |||
| Ceftriaxone | |||
| Oxacillin | |||
| Fluocloxacillin | |||
| Ceftaroline fosamil | |||
| Macrolides | Azithromycin | ||
| Clarithromyxin | |||
| Oxazolidinones | Linezolid | ||
| Glucopeptides | Vancomycin | ||
| Pleuromutilins | Lefamulin | ||
| Antibodies | Toxin neutralization | Ab pneumolysin | |
| Ab choline-binding PspA | |||
| Monoclonal Ab | Human mAb IgG1 AR-301 | ||
| mAb MED14893 | |||
| 2A3 and its variant LC10 | |||
| mAb LTM14 | |||
| Ab leukocidins | mAb ASN-1 | ||
| mAb ASN-2 | |||
| Natural compounds | Toxin prevention | Flavonoids | Baicalin |
| Morin hydrate | |||
| Liposomes | |||
| Hydroxamate inhibitor | GI254023X | ||
| Antimicrobial peptides | Bacterial killing | AMPs | CATH-1 |
| CATH-2 | |||
| CRAMP | |||
| LL-37 | |||
| Immune modulators | Reduction inflammatory response | Toll-like | |
| NOD-like | |||
| Phages and phage proteins | Bacterial killing | Phages AB-SA01 | |
| Pyophage | |||
| Phage Sb-1 | |||
| Lysin Cpl-1 | |||
| Lysin Pal | |||
| Lysin Cpl-7 | |||
| Lysin Cpl-711 | |||
| Lysin PL3 | |||
| Lysin ClyJ | |||
| Lysin 23TH_48 | |||
| Lysin SA01_53 | |||
| Lysin SAL200 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, L.; Cima-Cabal, M.D.; Duarte, A.C.; Rodríguez, A.; García-Suárez, M.d.M.; García, P. Gram-Positive Pneumonia: Possibilities Offered by Phage Therapy. Antibiotics 2021, 10, 1000. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081000
Fernández L, Cima-Cabal MD, Duarte AC, Rodríguez A, García-Suárez MdM, García P. Gram-Positive Pneumonia: Possibilities Offered by Phage Therapy. Antibiotics. 2021; 10(8):1000. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081000
Chicago/Turabian StyleFernández, Lucía, María Dolores Cima-Cabal, Ana Catarina Duarte, Ana Rodríguez, María del Mar García-Suárez, and Pilar García. 2021. "Gram-Positive Pneumonia: Possibilities Offered by Phage Therapy" Antibiotics 10, no. 8: 1000. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081000