
A Comprehensive Study of Historical Detection Data for Pathogen Isolates from U.S. Cattle
Abstract
:1. Introduction
2. Results
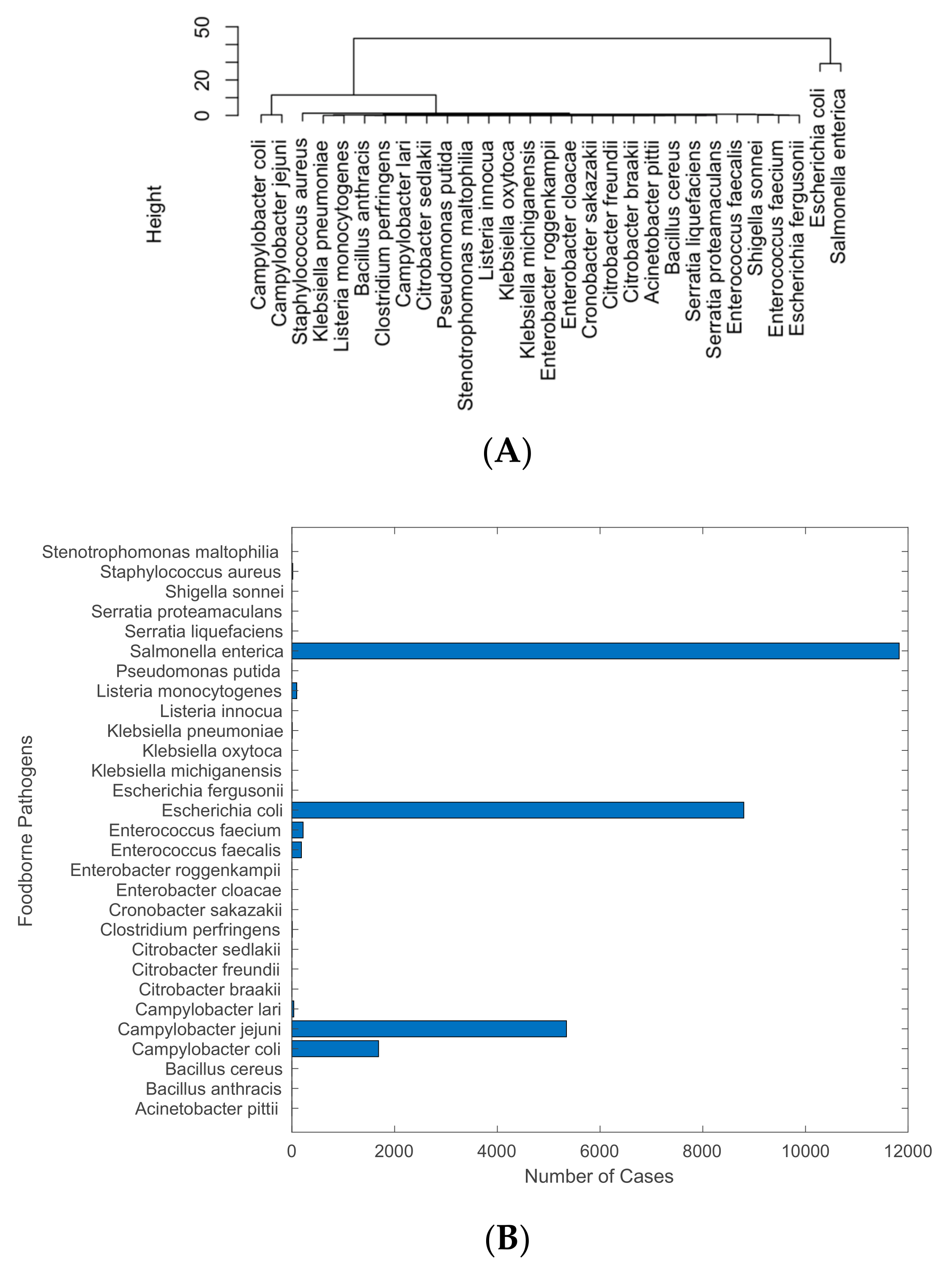
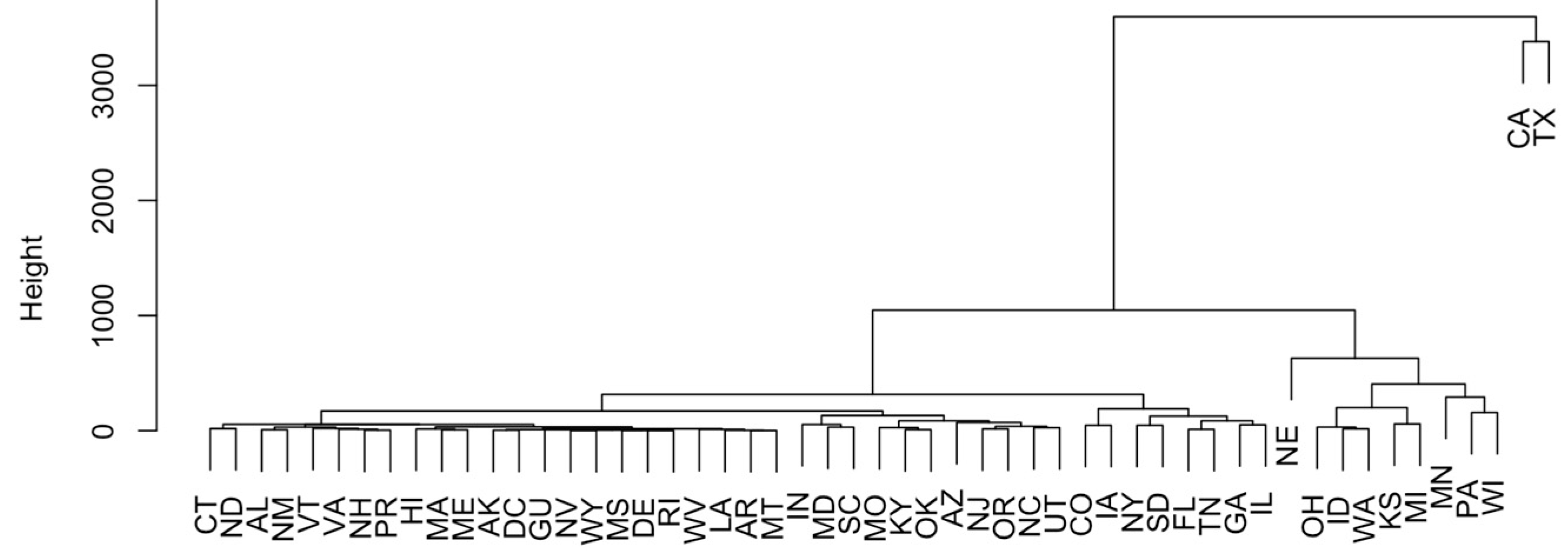
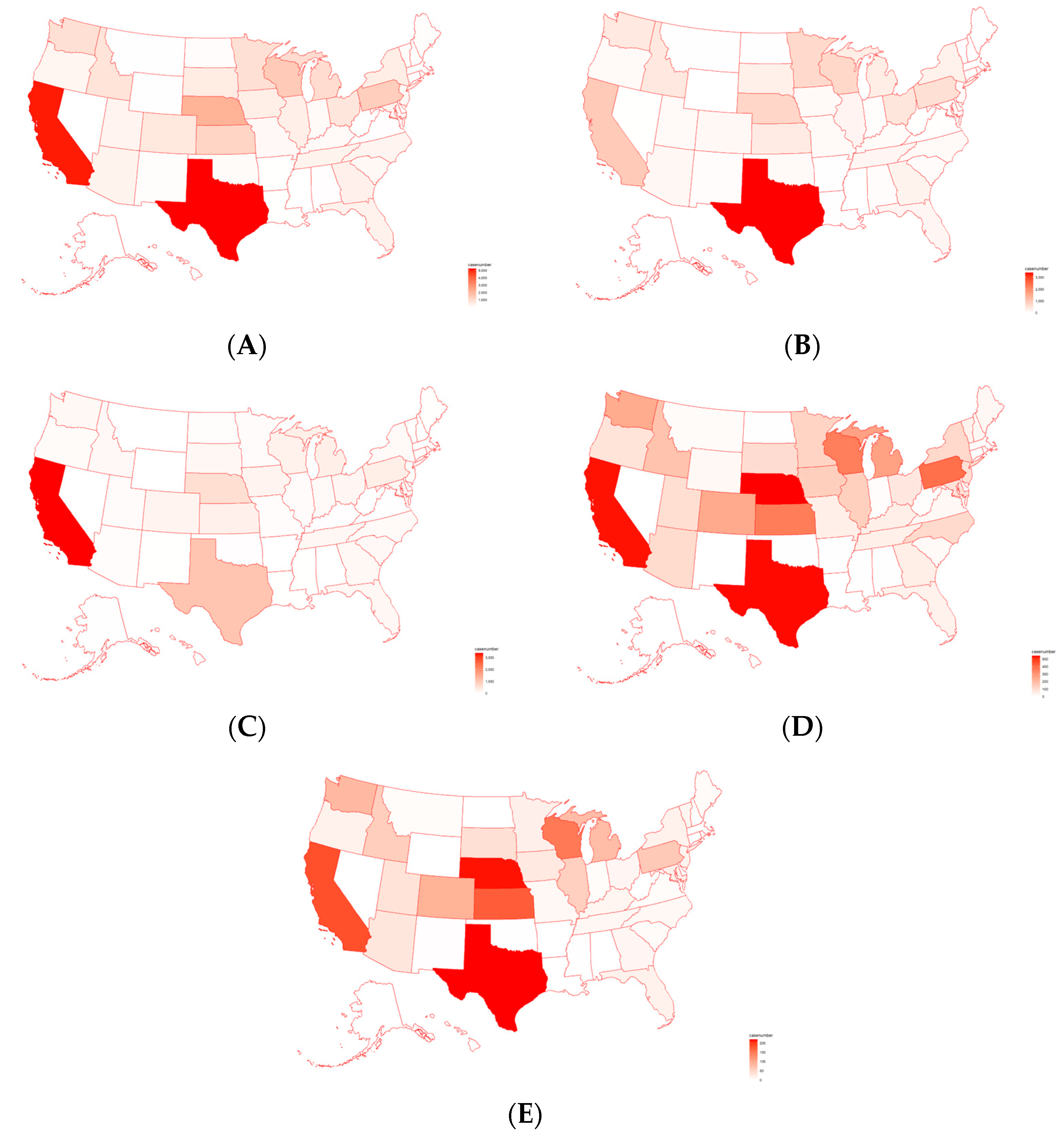
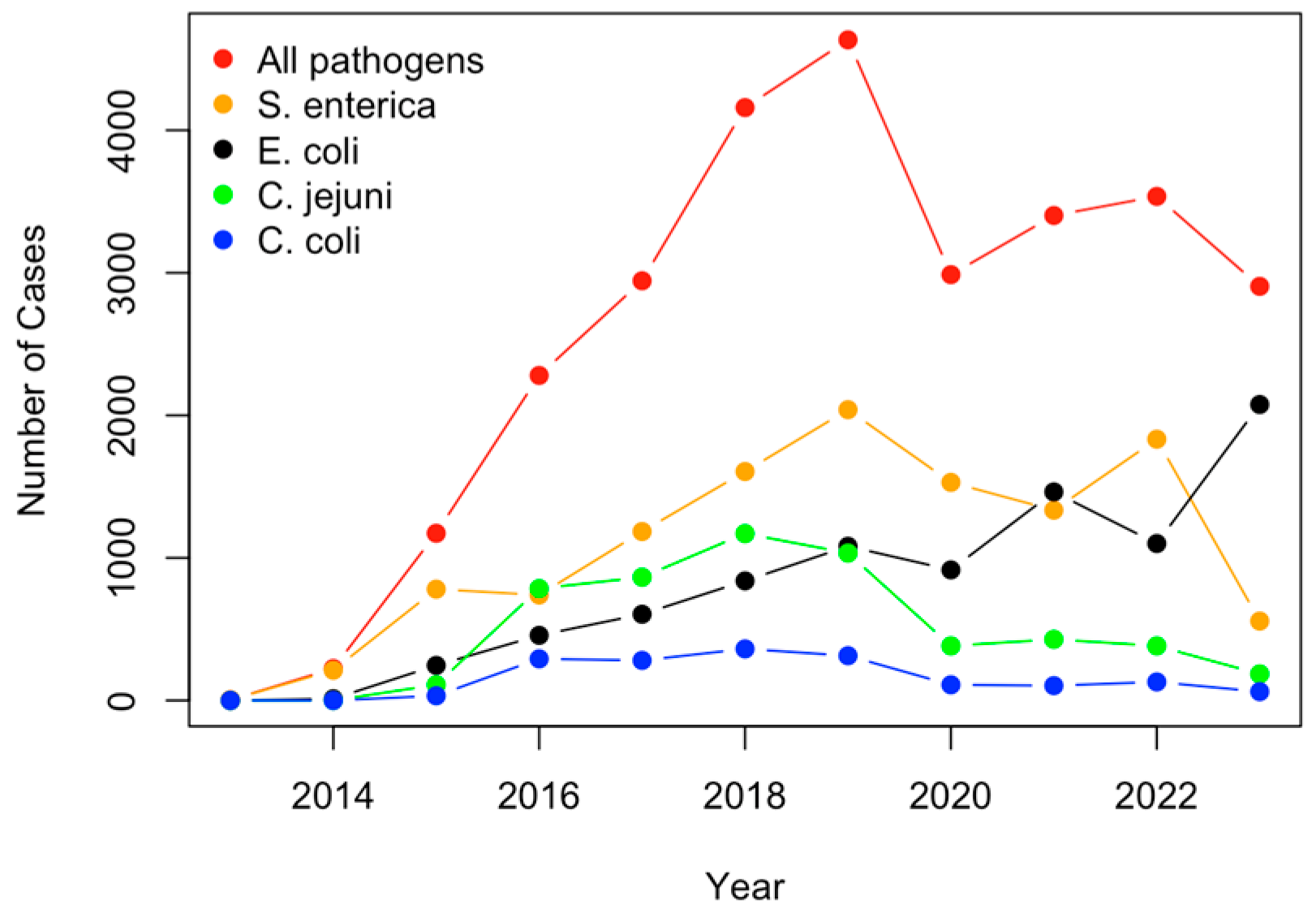
2.1. Analysis of Pathogens Isolated from U.S. Cattle
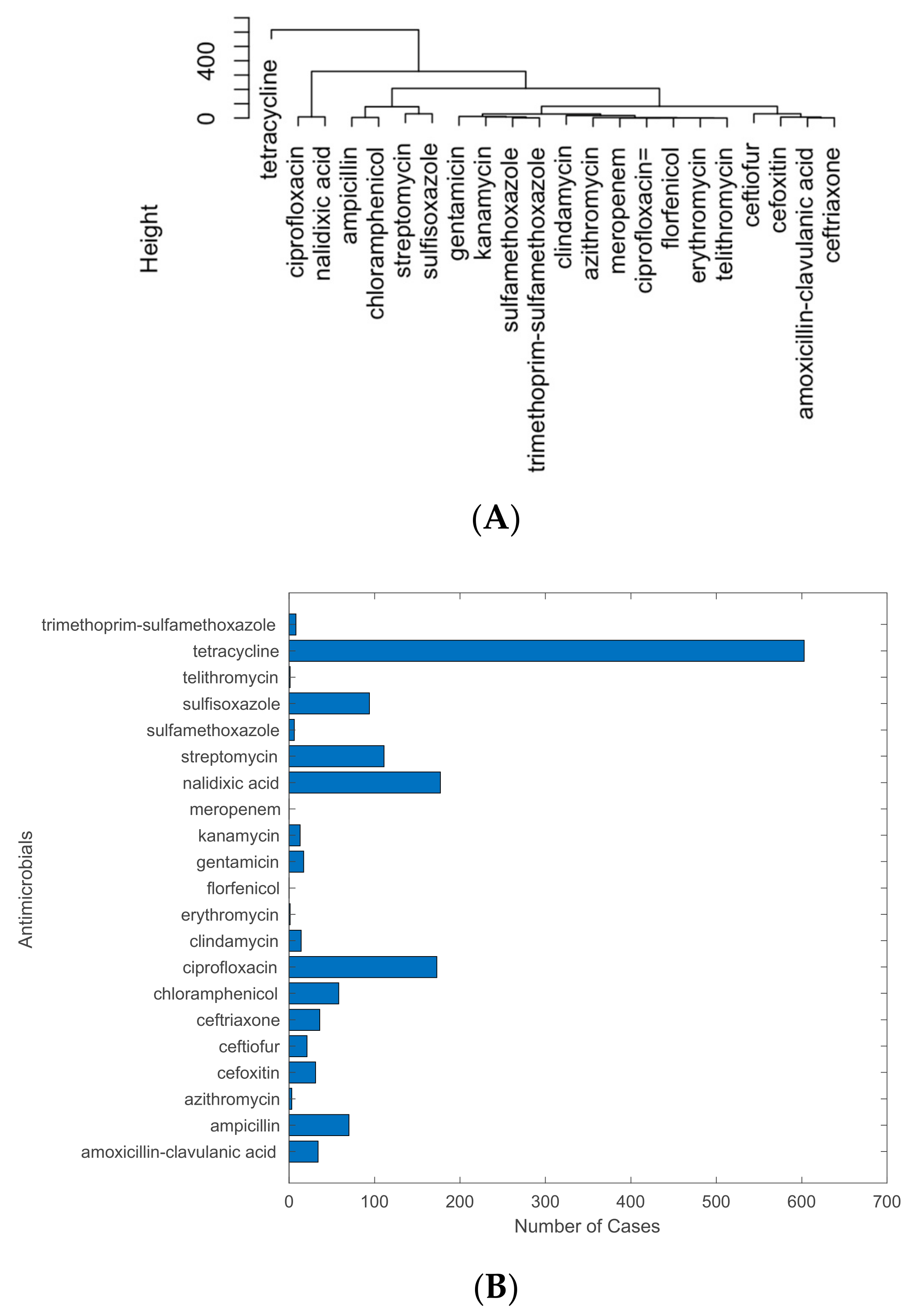
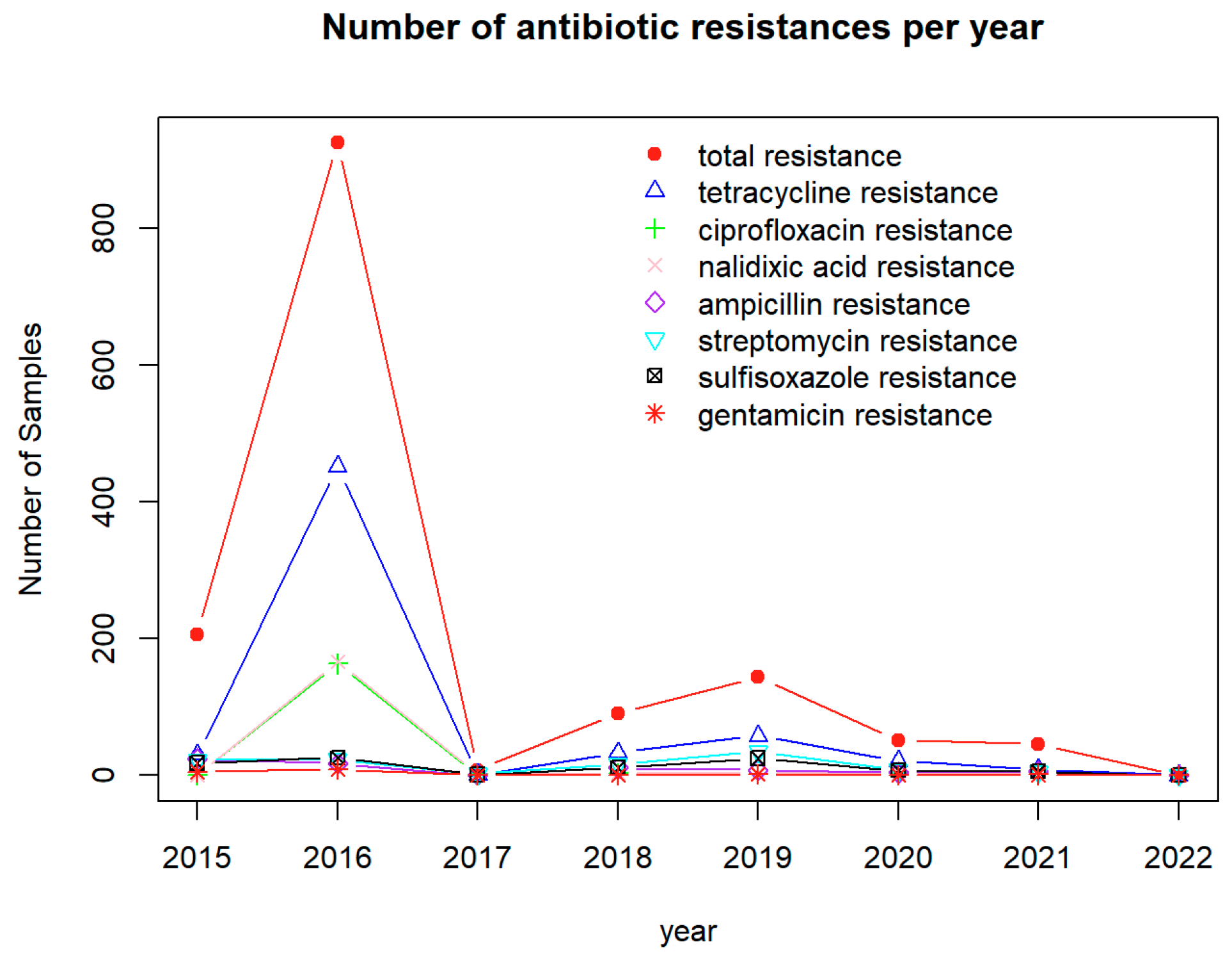
2.2. Analysis of Antimicrobials with Detected Resistance in Pathogen Isolates from U.S. Cattle
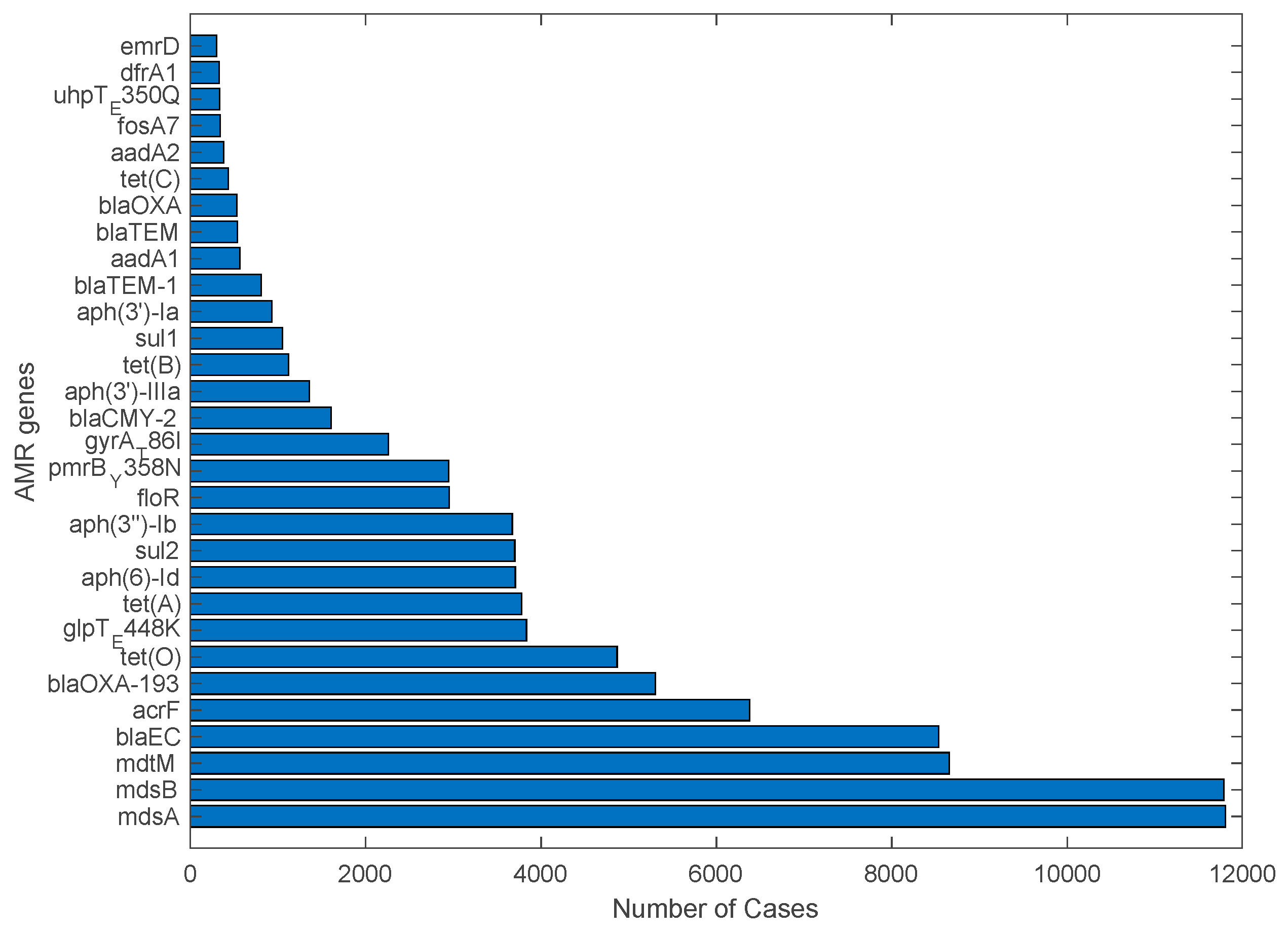
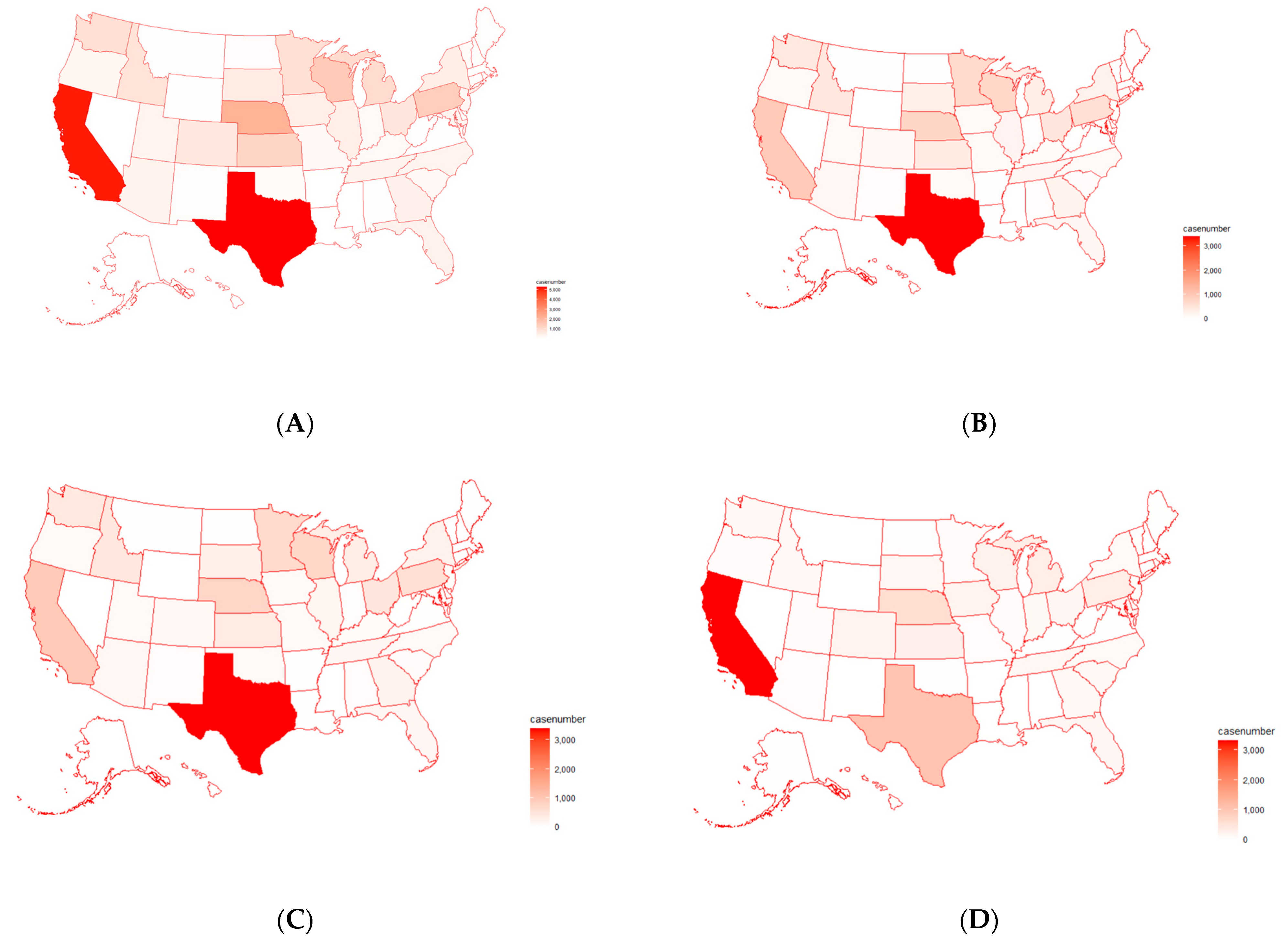
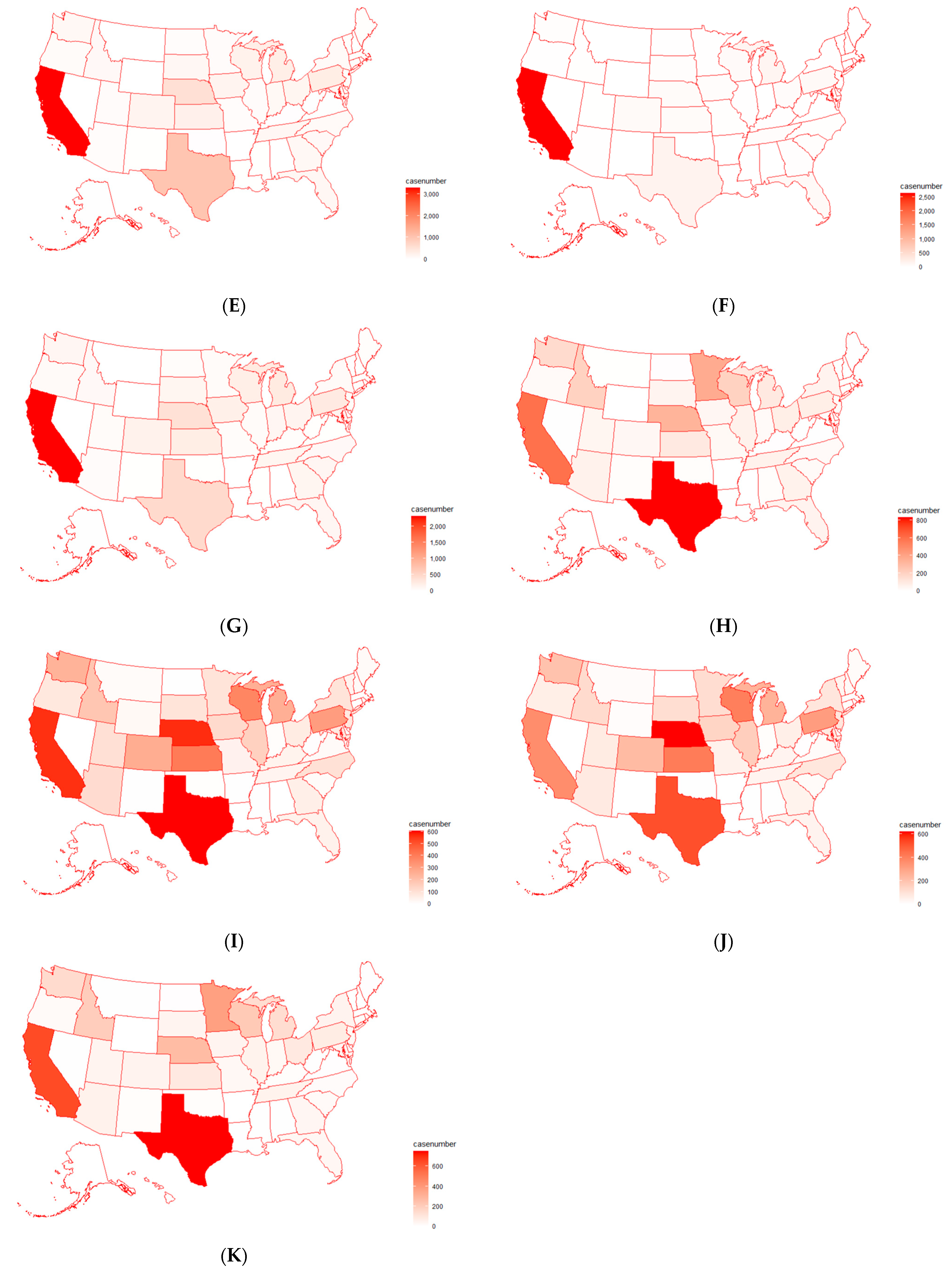
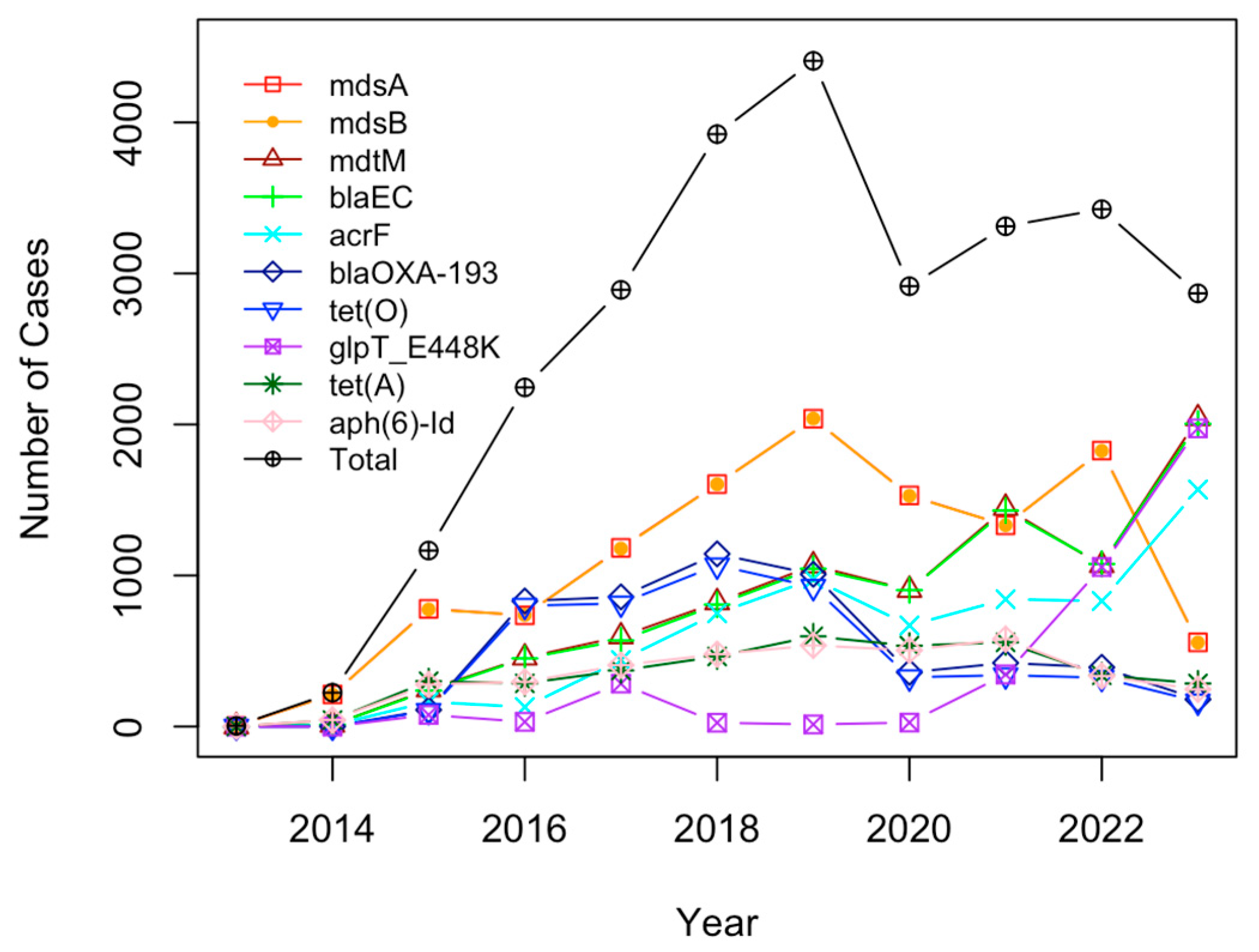
2.3. Analysis of Antimicrobial Resistance Genes in Pathogens Isolated from U.S. Cattle
3. Materials and Methods
3.1. Materials
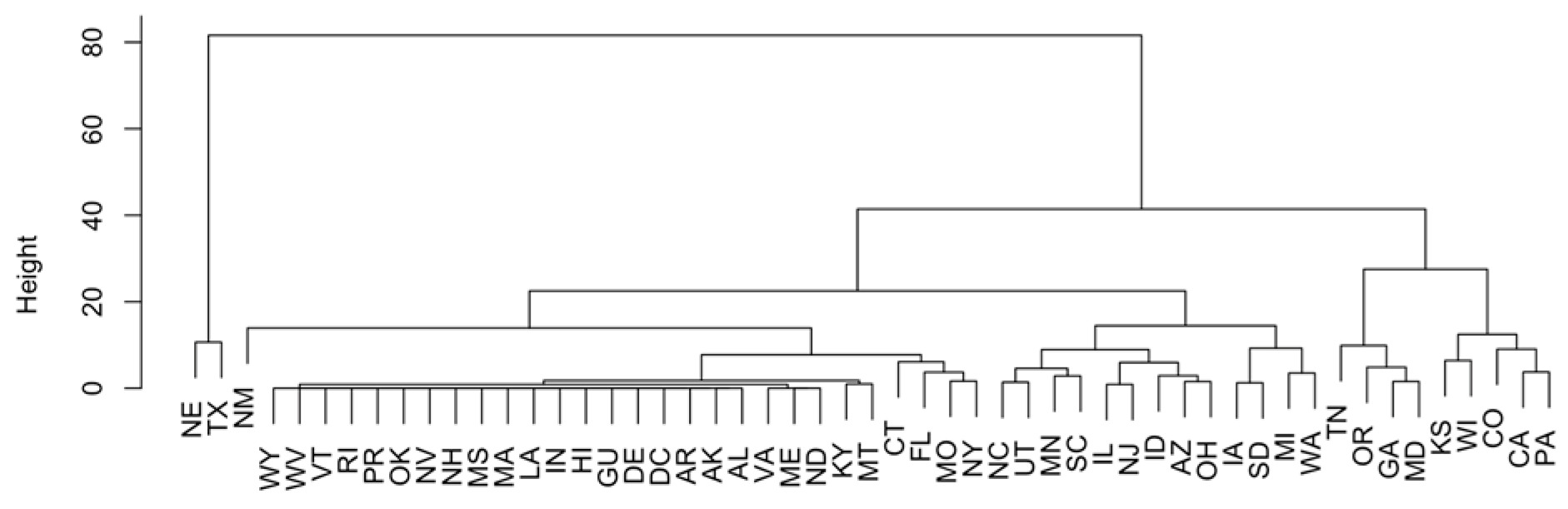
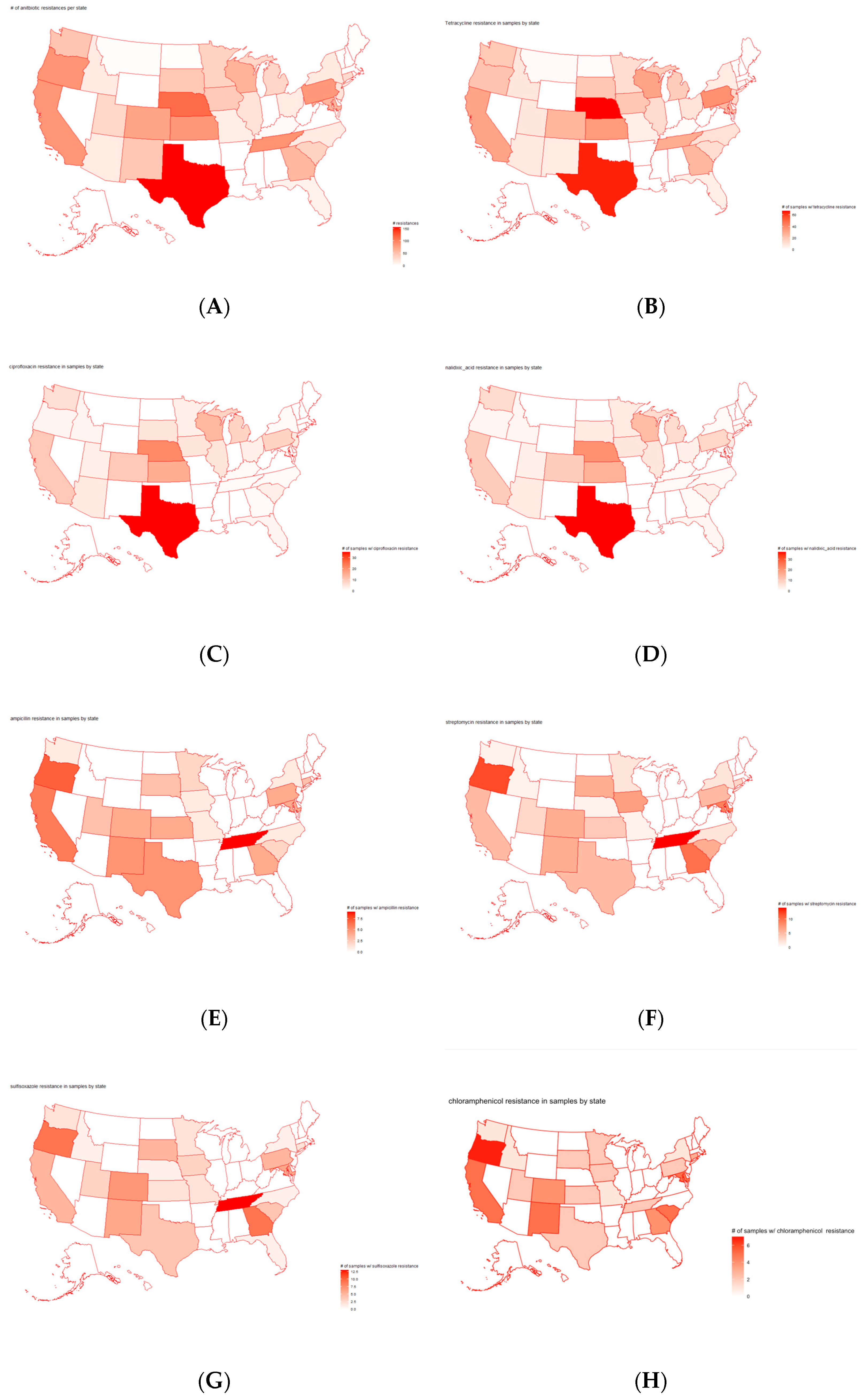
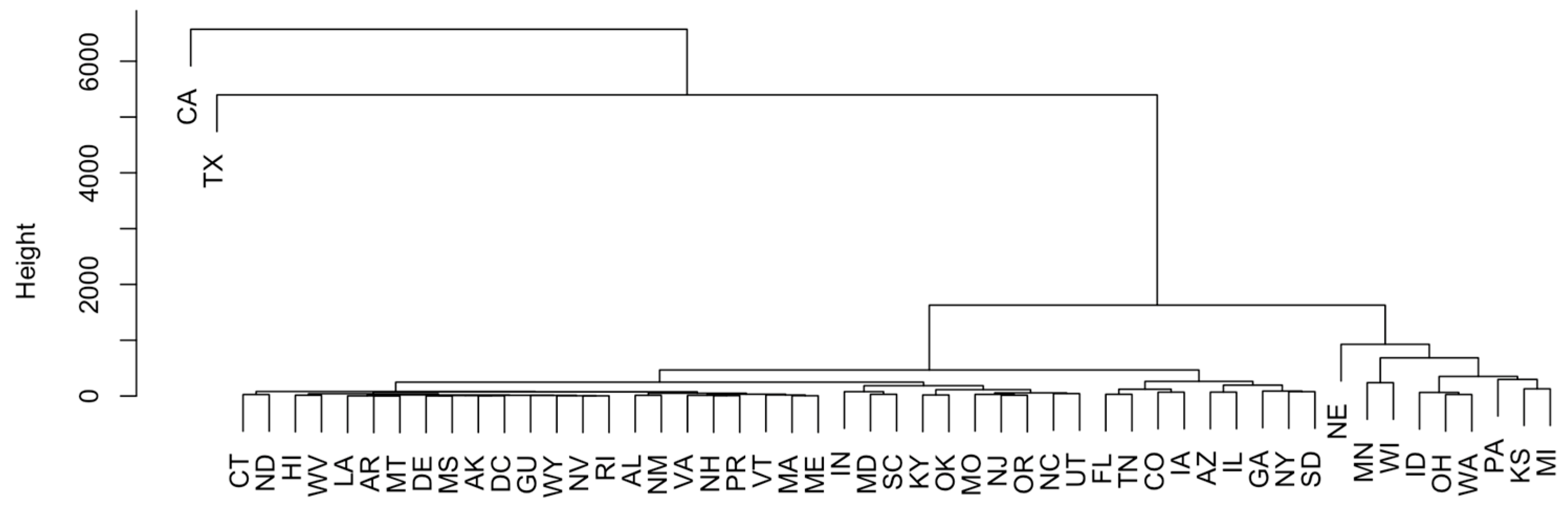
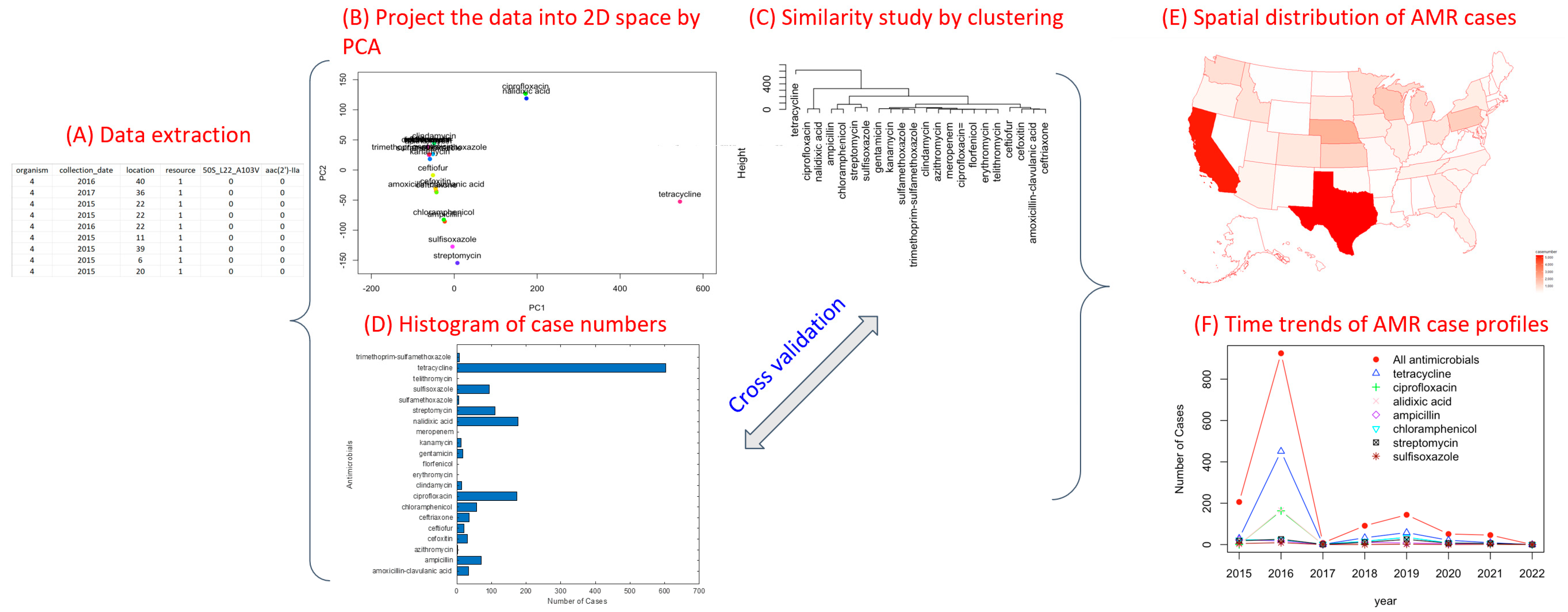
3.2. Methods: PCA, H-Clustering, and Spatiotemporal Trend Analysis
4. Discussion
4.1. Pathogens Mostly Detected from U.S. Cattle
4.2. Antimicrobials with the Most Detected Resistance by Pathogens Isolated from U.S. Cattle
4.3. Antimicrobial Resistance Genes Mostly Detected in Pathogen Isolates from U.S. Cattle
4.4. Limitation and Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bintsis, T. Patógenos transmitidos por los alimentos. AIMS Microbiol. 2017, 3. [Google Scholar] [CrossRef]
- Doyle, M.P.; Erickson, M.C. Reducing the Carriage of Foodborne Pathogens in Livestock and Poultry. Poult. Sci. 2006, 85, 960–973. [Google Scholar] [CrossRef]
- Oliver, S.P.; Jayarao, B.M.; Almeida, R.A. Foodborne pathogens, mastitis, milk quality, and dairy food safety. Foodborne Pathog. Dis. 2005, 1, 3–27. [Google Scholar]
- Hailu, W.; Helmy, Y.A.; Carney-Knisely, G.; Kauffman, M.; Fraga, D.; Rajashekara, G. Prevalence and Antimicrobial Resistance Profiles of Foodborne Pathogens Isolated from Dairy Cattle and Poultry Manure Amended Farms in Northeastern Ohio, the United States. Antibiotics 2021, 10, 1450. [Google Scholar] [CrossRef]
- National Agricultural Statistics Service West Virginia Field Office. Red Meat Production Cattle Inventory Sheep and Goat Inventories Farm Numbers and Land in Farms; National Agricultural Statistics Service West Virginia Field Office: Charleston, WV, USA, 2023; Volume 36, pp. 1–4.
- Sargeant, J.; Sanderson, M.; Smith, R.; Griffin, D. Escherichia coli O157 in feedlot cattle feces and water in four major feeder-cattle states in the USA. Prev. Veter- Med. 2003, 61, 127–135. [Google Scholar] [CrossRef]
- Munns, K.D.; Selinger, L.B.; Stanford, K.; Guan, L.; Callaway, T.R.; McAllister, T.A.; Wang, L.; Qu, K.; Li, X.; Cao, Z.; et al. Perspectives on Super-Shedding of Escherichia coli O157:H7 by Cattle. Foodborne Pathog. Dis. 2015, 12, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.E.; Callaway, T.R.; Nagaraja, T.; Josephs-Spaulding, J.; Beeler, E.; Singh, O.V.; Ekong, P.S.; Sanderson, M.W.; Cernicchiaro, N.; Huang, Q.; et al. Dietary Interactions and Interventions Affecting Escherichia coli O157 Colonization and Shedding in Cattle. Foodborne Pathog. Dis. 2009, 6, 785–792. [Google Scholar] [CrossRef]
- Ahmad, A.; Nagaraja, T.; Zurek, L. Transmission of Escherichia coli O157:H7 to cattle by house flies. Prev. Veter-Med. 2007, 80, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.J.; Warnick, L.D.; Elton, M.; Rodriguez-Rivera, L.D.; Siler, J.D.; Wright, E.M.; Gröhn, Y.T.; Wiedmann, M.; Pereira, R.V.; Rodríguez-Prieto, V.; et al. Salmonella enterica Serotype Cerro Among Dairy Cattle in New York: An Emerging Pathogen? Foodborne Pathog. Dis. 2010, 7, 659–665. [Google Scholar] [CrossRef]
- Threlfall, E.; Ward, L.R.; Frost, J.A.; Willshaw, G.A. The emergence and spread of antibiotic resistance in food-borne bacteria. Int. J. Food Microbiol. 2000, 62, 1–5. [Google Scholar] [CrossRef]
- Teuber, M. Spread of antibiotic resistance with food-borne pathogens. Cell. Mol. Life Sci. 1999, 56, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Silvester, R.; Madhavan, A.; Kokkat, A.; Parolla, A.; Adarsh, B.M.; Harikrishnan, M.; Abdulla, M.H. Global surveillance of antimicrobial resistance and hypervirulence in Klebsiella pneumoniae from LMICs: An in-silico approach. Sci. Total. Environ. 2022, 802, 149859. [Google Scholar] [CrossRef]
- DeDonder, K.; Apley, M.D. A literature review of antimicrobial resistance in Pathogens associated with bovine respiratory disease. Anim. Health Res. Rev. 2015, 16, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ahn, J. Emergence and spread of antibiotic-resistant foodborne pathogens from farm to table. Food Sci. Biotechnol. 2022, 31, 1481–1499. [Google Scholar] [CrossRef]
- Rodrigues, I.d.A.; Ferrari, R.G.; Panzenhagen, P.; dos Santos, A.M.P.; Rodrigues, G.L.; Junior, C.A.C.; Mano, S.B. The antibiotic resistome in Escherichia coli isolated from human, food, and animal sources. J. Appl. Microbiol. 2023, 134, lxac059. [Google Scholar] [CrossRef]
- Meroni, G.; Sora, V.M.; Martino, P.A.; Sbernini, A.; Laterza, G.; Zaghen, F.; Soggiu, A.; Zecconi, A. Epidemiology of Antimicrobial Resistance Genes in Streptococcus agalactiae Sequences from a Public Database in a One Health Perspective. Antibiotics 2022, 11, 1236. [Google Scholar] [CrossRef] [PubMed]
- Hull, D.M.; Harrell, E.; van Vliet, A.H.M.; Correa, M.; Thakur, S. Antimicrobial resistance and interspecies gene transfer in Campylobacter coli and Campylobacter jejuni isolated from food animals, poultry processing, and retail meat in North Carolina, 2018–2019. PLoS ONE 2021, 16, e0246571. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Bhattacharjee, A.S.; Phan, D.; Ashworth, D.; Schmidt, M.P.; Murinda, S.E.; Obayiuwana, A.; Murry, M.A.; Schwartz, G.; Lundquist, T.; et al. Potential reservoirs of antimicrobial resistance in livestock waste and treated wastewater that can be disseminated to agricultural land. Sci. Total. Environ. 2023, 872, 162194. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, E.; Tang, A.; Ye, M.C.; Wang, K.; Jia, Q.; Huang, Z. Data-Driven Analysis of Antimicrobial Resistance in Foodborne Pathogens from Six States within the US. Int. J. Environ. Res. Public Health 2019, 16, 1811. [Google Scholar] [CrossRef]
- Guo, X.; Akram, S.; Stedtfeld, R.; Johnson, M.; Chabrelie, A.; Yin, D.; Mitchell, J. Distribution of antimicrobial resistance across the overall environment of dairy farms—A case study. Sci. Total. Environ. 2021, 788, 147489. [Google Scholar] [CrossRef]
- Cui, K.; Gong, I.; Dong, A.; Yan, J.; Wang, M.; Huang, Z. Investigation of Virulence Genes Detected in Antimicrobial-Resistance Pathogens Isolates for Five Countries across the World. Processes 2020, 8, 1589. [Google Scholar] [CrossRef]
- Pei, R.; Zhang, L.; Duan, C.; Gao, M.; Feng, R.; Jia, Q.; Huang, Z. Investigation of Stress Response Genes in Antimicrobial Resistant Pathogens Sampled from Five Countries. Processes 2021, 9, 927. [Google Scholar] [CrossRef]
- Li, M.; Wang, K.; Tang, A.; Tang, A.; Chen, A.; Huang, Z. Investigation of the Genes Involved in the Outbreaks of Escherichia coli and Salmonella spp. in the United States. Antibiotics 2021, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zheng, J.; Deng, T.; Peng, J.; Daniel, D.; Jia, Q.; Huang, Z. An Analysis of Antimicrobial Resistance of Clinical Pathogens from Historical Samples for Six Countries. Processes 2019, 7, 964. [Google Scholar] [CrossRef]
- Hanes, R.; Huang, Z. Investigation of Antimicrobial Resistance in Listeria monocytogenes from Historical Isolate Data. Int. J. Environ. Res. Public Health 2022, 19, 5506. [Google Scholar] [CrossRef]
- Yang, K.; Wang, A.; Fu, M.; Wang, A.; Chen, K.; Jia, Q.; Huang, Z. Investigation of Incidents and Trends of Antimicrobial Resistance in Foodborne Pathogens in Eight Countries from Historical Sample Data. Int. J. Environ. Res. Public Health 2020, 17, 472. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley interdisciplinary reviews: Computational statistics. In Wiley Interdisplinary Reviews: Computational Statistics; Wiley Online Library: New York, NY, USA, 2010. [Google Scholar]
- Pearson, K. On lines and planes of closest fit to systems of points in space. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1901, 2, 559–572. [Google Scholar] [CrossRef]
- Hotelling, H. Analysis of a complex of statistical variables into principal components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Joliffe, I.; Morgan, B. Principal component analysis and exploratory factor analysis. Stat. Methods Med. Res. 1992, 1, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Sarle, W.S.; Jain, A.K.; Dubes, R.C. Algorithms for Clustering Data. Technometrics 1990, 32, 227–229. [Google Scholar] [CrossRef]
- Rafsanjani, M.K.; Varzaneh, Z.A.; Chukanlo, N.E. A Survey of Hierarchical Clustering Algorithms. J. Math. Comput. Sci. 2012, 5, 229–240. [Google Scholar] [CrossRef]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Kalunke, R.M.; Grasso, G.; D’Ovidio, R.; Dragone, R.; Frazzoli, C. Detection of ciprofloxacin residues in cow milk: A novel and rapid optical β-galactosidase-based screening assay. Microchem. J. 2018, 136, 128–132. [Google Scholar] [CrossRef]
- Song, S.; Hwang, S.; Lee, S.; Ha, N.-C.; Lee, K. Interaction Mediated by the Putative Tip Regions of MdsA and MdsC in the Formation of a Salmonella-Specific Tripartite Efflux Pump. PLoS ONE 2014, 9, e100881. [Google Scholar] [CrossRef] [PubMed]
- Law, C.J.; Alegre, K.O. Clamping down on drugs: The Escherichia coli multidrug efflux protein MdtM. Res. Microbiol. 2018, 169, 461–467. [Google Scholar] [CrossRef]
- Adator, E.H.; Walker, M.; Narvaez-Bravo, C.; Zaheer, R.; Goji, N.; Cook, S.R.; Tymensen, L.; Hannon, S.J.; Church, D.; Booker, C.W.; et al. Whole Genome Sequencing Differentiates Presumptive Extended Spectrum Beta-Lactamase Producing Escherichia coli along Segments of the One Health Continuum. Microorganisms 2020, 8, 448. [Google Scholar] [CrossRef]
- Blair, J.M.; Piddock, L.J. Structure, function and inhibition of RND efflux pumps in Gram-negative bacteria: An update. Curr. Opin. Microbiol. 2009, 12, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Habib, I.; Mohamed, M.-Y.I.; Ghazawi, A.; Lakshmi, G.B.; Khan, M.; Li, D.; Sahibzada, S. Genomic characterization of molecular markers associated with antimicrobial resistance and virulence of the prevalent Campylobacter coli isolated from retail chicken meat in the United Arab Emirates. Curr. Res. Food Sci. 2023, 6, 100434. [Google Scholar] [CrossRef]
- Altayb, H.N.; Elbadawi, H.S.; Alzahrani, F.A.; Baothman, O.; Kazmi, I.; Nadeem, M.S.; Hosawi, S.; Chaieb, K. Co-Occurrence of β-Lactam and Aminoglycoside Resistance Determinants among Clinical and Environmental Isolates of Klebsiella pneumoniae and Escherichia coli: A Genomic Approach. Pharmaceuticals 2022, 15, 1011. [Google Scholar] [CrossRef]
- Fang, Y.; Tran, F.; Stanford, K.; Yang, X. Stress Resistance and Virulence Gene Profiles Associated with Phylogeny and Phenotypes of Escherichia coli from Cattle. J. Food Prot. 2023, 86, 100122. [Google Scholar] [CrossRef]
- Hedayatianfard, K.; Akhlaghi, M.; Sharifiyazdi, H. Detection of tetracycline resistance genes in bacteria isolated from fish farms using polymerase chain reaction. Veter-Res. Forum Int. Q. J. 2014, 5, 269–275. [Google Scholar]
- Ashenafi, M.; Ammosova, T.; Nekhai, S.; Byrnes, W.M. Purification and characterization of aminoglycoside phosphotransferase APH(6)-Id, a streptomycin-inactivating enzyme. Mol. Cell. Biochem. 2013, 387, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L. ggplot2: Elegant Graphics for Data Analysis by WICKHAM, H. Biometrics 2011, 67, 678–679. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Fitzgerald, A.; Edrington, T.; Looper, M.; Callaway, T.; Genovese, K.; Bischoff, K.; McReynolds, J.; Thomas, J.; Anderson, R.; Nisbet, D. Antimicrobial susceptibility and factors affecting the shedding of E. coli O157:H7 and Salmonella in dairy cattle. Lett. Appl. Microbiol. 2003, 37, 392–398. [Google Scholar] [CrossRef]
- Flach, M.G.; Dogan, O.B.; Kreikemeier, W.M.; Nightingale, K.K.; Brashears, M.M. Prevalence of Foodborne Pathogens in Pacific Northwest Beef Feedlot Cattle Fed Two Different Direct-Fed Microbials. J. Food Prot. 2023, 86, 100139. [Google Scholar] [CrossRef] [PubMed]
- Gelalcha, B.D.; Ensermu, D.B.; Agga, G.E.; Vancuren, M.; Gillespie, B.E.; D’Souza, D.H.; Okafor, C.C.; Dego, O.K. Prevalence of Antimicrobial Resistant and Extended-Spectrum Beta-Lactamase-producing Escherichia coli in Dairy Cattle Farms in East Tennessee. Foodborne Pathog. Dis. 2022, 19, 408–416. [Google Scholar] [CrossRef]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in Food and Feedingstuffs: From Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. J. Anal. Methods Chem. 2017, 2017, 1315497. [Google Scholar] [CrossRef]
- Wallinga, D. Better Burgers: Why It’s High Time the U.S. Beef Industry Kicked Its Antibiotics Habit; NRDC: New York, NY, USA, 2020. [Google Scholar]
- Wallinga, D.; Smit, L.A.M.; Davis, M.F.; Casey, J.A.; Nachman, K.E. A Review of the Effectiveness of Current US Policies on Antimicrobial Use in Meat and Poultry Production. Curr. Environ. Health Rep. 2022, 9, 339–354. [Google Scholar] [CrossRef]
- FDA. Summary Report on Antimicrobials Sold or Distributed for Use in Food-Producing Animals; FDA: Silver Spring, MD, USA, 2020.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Collection Date | Location | Resource | 50S_L22_A103V | aac(3)-Via | aac(3)-IId | Streptomycin | Sulfamethoxazole | Sulfisoxazole | Telithromycin | Tetracycline | Trimethoprim-sulfamethoxazole | Sample |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 2016 | 40 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 447 |
| 4 | 2017 | 36 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 450 |
| 4 | 2015 | 22 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 451 |
| 4 | 2015 | 22 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 452 |
| 4 | 2016 | 22 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 453 |
| 4 | 2015 | 11 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 535 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, G.; Pei, H.; Zhou, A.; Fan, B.; Zhou, H.; Choi, A.; Huang, Z. A Comprehensive Study of Historical Detection Data for Pathogen Isolates from U.S. Cattle. Antibiotics 2023, 12, 1509. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12101509
Gu G, Pei H, Zhou A, Fan B, Zhou H, Choi A, Huang Z. A Comprehensive Study of Historical Detection Data for Pathogen Isolates from U.S. Cattle. Antibiotics. 2023; 12(10):1509. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12101509
Chicago/Turabian StyleGu, George, Henry Pei, Alan Zhou, Brianna Fan, Hanlin Zhou, Austin Choi, and Zuyi Huang. 2023. "A Comprehensive Study of Historical Detection Data for Pathogen Isolates from U.S. Cattle" Antibiotics 12, no. 10: 1509. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12101509