
Genetic Diversity and Resistome Analysis of Campylobacter lari Isolated from Gulls in Croatia
 , , , , and
, , , , and
Abstract
:1. Introduction
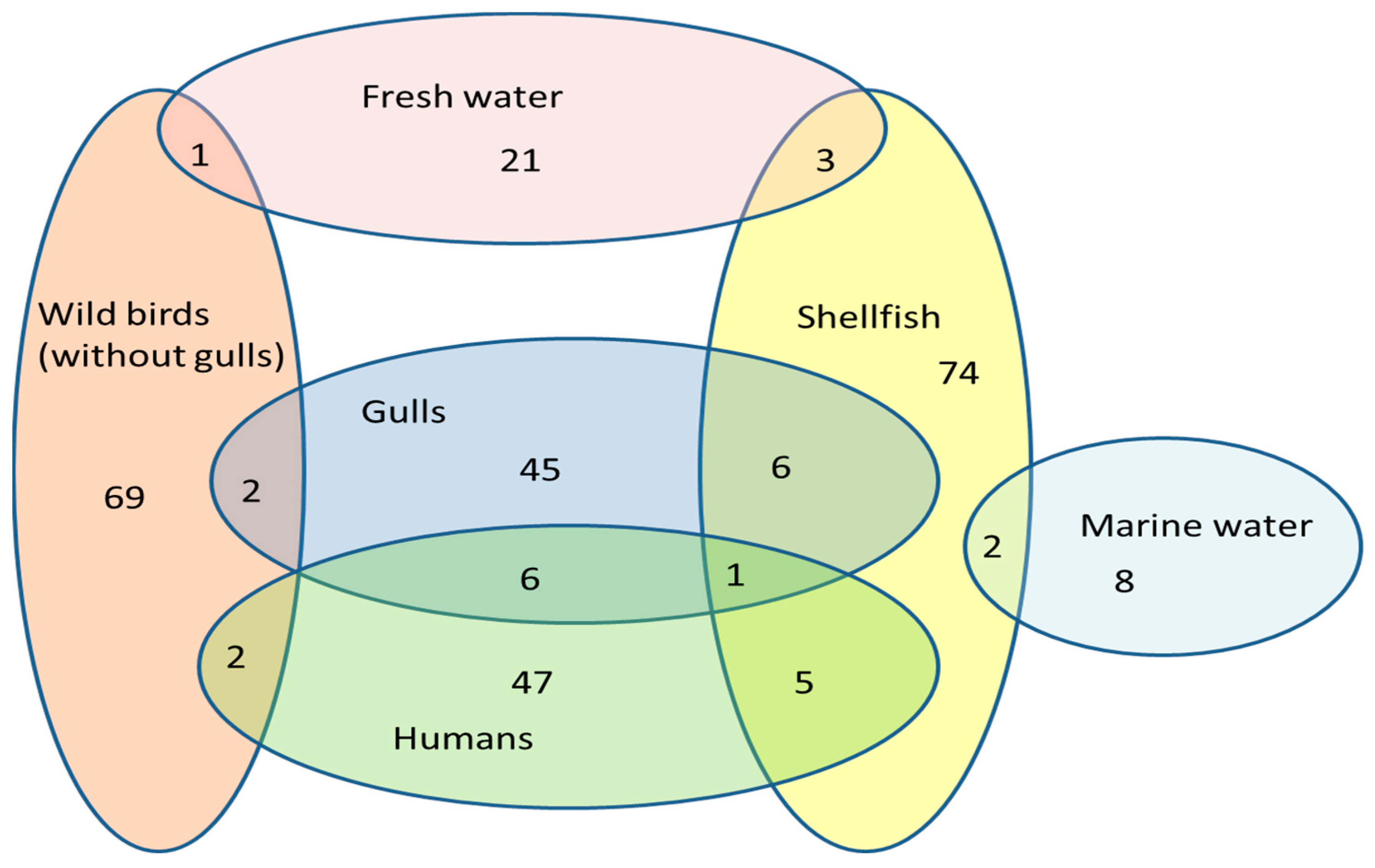
2. Results
2.1. Genotypic Determination of Antimicrobial Resistance (AMR)
2.2. Comparison between Phenotypic and Genotypic AMRs
3. Discussion
4. Materials and Methods
4.1. Sampling and Campylobacter Isolation
4.2. MLST
4.3. Antimicrobial Susceptibility Testing
4.4. Genomics
4.4.1. Whole-Genome Sequencing Data Analysis
4.4.2. Antimicrobial Resistance
4.4.3. Genotypic–Phenotypic Comparisons
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA; ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 20–31. [Google Scholar]
- Tauxe, R.V.; Patton, C.M.; Edmonds, P.; Barrett, T.J.; Brenner, J.; Blake, P.A. Illness associated with Campylobacter laridis, a newly recognized Campylobacter species. J. Clin. Microbiol. 1985, 21, 222–225. [Google Scholar] [CrossRef]
- Namamkin, I.; Stowell, C.; Skalina, D.; Jones, A.M.; Hoop, R.M.; Smibert, M. Campylobacter laridis causing bacteremia in an immunocompromised host. Ann. Intern. Med. 1984, 101, 55–57. [Google Scholar] [CrossRef]
- Dionisio, D.; Milo, D.; Mazzotta, D.; Cecile, P. Campylobacter laridis bacteremia in an AIDS patient. Boll Lst Sieroter Milan 1989, 68, 199–200. [Google Scholar]
- Werno, A.M.; Klena, J.D.; Shaw, G.M.; Murdoch, D.R. Fatal Case of Campylobacter lari Prosthetic Joint Infection and Bacteremia in an Immunocompetent Patient. J. Clin. Microbiol. 2002, 40, 1053–1055. [Google Scholar] [CrossRef] [Green Version]
- Bezian, M.C.; Ribou, G.; Barberis-Giletti, C.; Megraud, F. Isolation of a urease positive thermophilic variant of Campylobacter Ian from a patient with urinary tract infection. Eur. J. Clin. Microbiol. Infect. Dis. 1990, 9, 895–897. [Google Scholar] [CrossRef]
- Bruneau, B.; Burc, L.; Bizet, C.; Lambert-Zechovsky, N.; Branger, N.C. Purulent pleurisy caused by Campylobacter lari. Eur. J. Clin. Microbiol. Infect. Dis. 1998, 17, 185–188. [Google Scholar] [CrossRef]
- Mori, E.; Hashimoto, T.; Yahiro, T.; Miura, M.; Ishihara, T.; Miyazaki, M.; Komiya, K.; Takahashi, N.; Nishizono, A.; Hiramatsu, K. Campylobacter lari vertebral osteomyelitis. Jpn. J. Infect. Dis. 2022, 75, 322–324. [Google Scholar] [CrossRef]
- Benjamin, J.; Leaper, S.; Owen, R.J.; Skirrow, M.B. Description of Campylobacter laridis, a new species comprising the nalidixic acid resistant thermophilic Campylobacter (NARTC) group. Curr. Microbiol. 1983, 8, 231–238. [Google Scholar] [CrossRef]
- Murayama, S.; Tanaka, T.; Katsube, Y.; Nakanishi, H.; Nukina, M. Prevalence of thermophilic Campylobacters in crows (Corvus levaillantii, Corvus corone) and serogroups of the isolates. Nippon. Juigaku Zasshi 1990, 52, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Endtz, H.P.; Vliegenthart, J.S.; Vandamme, P.; Weverink, H.W.; van den Braak, N.P.; Verbrugh, H.A.; van Belkum, A. Genotypic diversity of Campylobacter lari isolated from mussels and oysters in The Netherlands. Int. J. Food Microbiol. 1997, 34, 79–88. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Jones, K.; Anderton, S.; Andrew, S. Campylobacters in wild birds: Identification and molecular characterization. In Proceedings of the Campylobacter, Helicobacter and related organisms, Cape Town, South Africa, 15–19 September 1997; Lastovica, A.J., Newell, D.G., Lastovica, E.E., Eds.; Rustica Press: Cape Town, South Africa, 1998; p. 80. [Google Scholar]
- Obiri-Danso, K.; Jones, K. Distribution and seasonality of microbial indicators and thermophilic campylobacters in two freshwater bathing sites on the River Lune in northwest England. J. Appl. Microbiol. 1999, 87, 822–832. [Google Scholar] [CrossRef]
- Waldenstrom, J.; Broman, T.; Carlsson, I.; Hasselquit, D.; Achterberg, R.P.; Wagenaar, J.A.; Olsen, B. Prevalence of Campylobacter jejuni, Campylobacter lari, and Campylobacter coli in different ecological guilds and taxa of migrating birds. Appl. Environ. Microbiol. 2002, 68, 5911–5917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coloe, P.J.; Slattery, J.F.; Cavanaugh, P.; Vaughan, J. The cellular fatty acid composition of Campylobacter species isolated from cases of enteritis in man and animals. J. Hyg. 1986, 96, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duim, B.; Vandamme, P.A.R.; Rigter, A.; Laevens, S.; Dijkstra, J.R.; Wagenaar, J.A. Differentiation of Campylobacter species by AFLP fingerprinting. Microbiology 2001, 147, 2729–2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totten, P.A.; Patton, C.M.; Tenover, F.C.; Barrett, T.J.; Stamm, W.E.; Steigerwalt, A.G.; Lin, J.Y.; Holmes, K.K.; Brenner, D.J. Prevalence and characterization of hippurate-negative Campylobacter jejuni in King County, Washington. J. Clin. Microbiol. 1987, 25, 1747–1752. [Google Scholar] [CrossRef]
- Bolton, F.J.; Holt, A.V.; Hutchinson, D.N. Urease-positive thermophilic campylobacters. Lancet 1985, 325, 1217–1218. [Google Scholar] [CrossRef]
- Fricker, C.R.; Park, R.W.A. A two-year study of the distribution of ‘thermophilic’ campylobacters in human, environmental and food samples from the Reading area with particular reference to toxin production and heat-stable serotype. J. Appl. Bacteriol. 1989, 66, 477–490. [Google Scholar] [CrossRef]
- Mawer, S.L. Campylobacters in man and the environment in Hull and East Yorkshire. Epidemiol. Infect. 1988, 101, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Welcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Jurinović, L.; Sokolović, M.; Šimpraga, B.; Savić, V.; Krstulović, F.; Balenović, M.; Berendika, M. Značaj galebova, Laridae, Aves, kao kliconoša određenih virusnih i bakterijskih zoonoza. In Zaštita Okoliša i Održivo Gospodarenje Resursima; Žunec, N., Špralja Šakić, C., Eds.; Business Media: Zagreb, Croatia, 2014. [Google Scholar]
- Jurinović, L.; Kralj, J.; Ječmenica, B.; Brajdić, N. Kretanje galebova koji se hrane na odlagalištu otpada Prudinec u Jakuševcu. In Proceedings of the XV International Symposium of Waste Management; Anić Vučinić, A., Ed.; Faculty of Geotechnical Engineering, University of Zagreb: Zagreb, Croatia, 2018. [Google Scholar]
- Jurinović, L.; Ječmenica, B.; Džafić, N.; Brlek Gorski, D.; Šimpraga, B.; Krstulović, F.; Amšel Zelenika, T.; Humski, A. First Data on Campylobacter spp. Presence in Shellfish in Croatia. Pathogens 2022, 11, 943. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority); Amore, G.; Beloeil, P.-A.; Garcia Fierro, R.; Guerra, B.; Papanikolaou, A.; Rizzi, V.; Stoicescu, A.-V. Manual for reporting 2022 antimicrobial resistance data within the framework of Directive 2003/99/EC and Decision 2020/1729/EU. EFSA Support. Publ. 2023, 20, EN-7826. [Google Scholar] [CrossRef]
- Olsen, K.M.; Larsson, H. Gulls of North America, Europe and Asia; Princeton University Press: London, UK, 2004. [Google Scholar]
- Glünder, G. NaCl-tolerance of Campylobacter isolates from birds and Campylobacter type strains and variation of their serological behaviour. Zentralbl Vet. B 1993, 40, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Obiri-Danso, K.; Paul, N.; Jones, K. The effects of UVB and temperature on the survival of natural populations and pure cultures of Campylobacter jejuni, Camp. coli, Camp. lari and urease-positive thermophilic campylobacters (UPTC) in surface waters. J. Appl. Microbiol. 2001, 90, 256–267. [Google Scholar] [CrossRef]
- Keller, J.I.; Shriver, W.G. Prevalence of three Campylobacter species, C. jejuni, C. coli, and C. lari, using multi locus sequence typing in wild birds of the mid-atlantic region, USA. J. Wildl. Dis. 2014, 50, 31–41. [Google Scholar] [CrossRef]
- Migura-Garcia, L.; Ramos, R.; Cerdà-Cuéllar, M. Antimicrobial resistance of Salmonella serovars and Campylobacter spp. isolated from an opportunistic gull species, yellow-legged gull (Larus michahellis). J. Wildl. Dis. 2017, 53, 148–152. [Google Scholar] [CrossRef]
- Broman, T.; Palmgren, H.; Bergström, S.; Sellin, M.; Waldenström, J.; Danielsson-Tham, M.L.; Olsen, B. Campylobacter jejuni in Black-Headed Gulls (Larus ridibundus): Prevalence, Genotypes, and Influence on C. jejuni Epidemiology. J. Clin. Microbiol. 2002, 40, 4594–4602. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J.; Ricci, V.; Pumbwe, L.; Everett, M.J.; Griggs, D.J. Fluoroquinolone resistance in Campylobacter species from man and animals: Detection of mutations in topoisomerase genes. J. Antimicrob. Chemother. 2003, 51, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Weis, A.M.; Storey, D.B.; Taff, C.C.; Townsend, A.K.; Huang, B.C.; Kong, N.T.; Clothier, K.A.; Spinner, A.; Byrne, B.A.; Weimer, B.C. Genomic comparison of Campylobacter spp. and their potential for zoonotic transmission between birds, primates, and livestock. Appl. Environ. Microbiol. 2016, 82, 7165–7175. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Mendoza, D.; Martínez-Flores, I.; Santamaría, R.I.; Lozano, L.; Bustamante, V.H.; Pérez-Morales, D. Genomic Analysis Reveals the Genetic Determinants Associated With Antibiotic Resistance in the Zoonotic Pathogen Campylobacter spp. Distributed Globally. Front. Microbiol. 2020, 11, 513070. [Google Scholar] [CrossRef]
- Gourmelon, M.; Boukerb, A.M.; Nabi, N.; Banerji, S.; Joensen, K.G.; Serghine, J.; Cormier, A.; Megraud, F.; Lehours, P.; Alter, T.; et al. Genomic Diversity of Campylobacter lari Group Isolates from Europe and Australia in a One Health Context. Appl. Environ. Microbiol. 2022, 88, e0136822. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-Lactamase DataBase (BLDB)—Structure and Function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [Green Version]
- Luo, N.; Sahin, O.; Lin, J.; Michel, L.O.; Zhang, Q. In vivo selection of Campylobacter isolates with high levels of fluoroquinolone resistance associated with gyrA mutations and the function of the CmeABC efflux pump. Antimicrob. Agents. Chemother. 2003, 47, 390–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hoek, A.H.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, Y.; Fu, Q.; Wang, Y.; Li, X.; Wu, C.; Shen, Z.; Zhang, Q.; Qin, P.; Shen, J.; et al. Integrated genomic and proteomic analyses of high-level chloramphenicol resistance in Campylobacter jejuni. Sci. Rep. 2017, 7, 16973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- EN ISO 10271-1:2017; Microbiology of the Food Chain—Horizontal Method for Detection and Enumeration of Campylobacter spp.—Part 1: Detection Method. International Organization for Standardization (ISO): Geneva, Switzerland, 2017.
- Wang, G.; Clark, C.G.; Taylor, T.M.; Pucknell, C.; Barton, C.; Price, L.; Woodward, D.L.; Rodgers, F.G. Colony multiplex PCR assay for identification and differentiation of Campylobacter jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus. J. Clin. Microbiol. 2002, 40, 4744–4747. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.G.; On, S.L.; Wang, G.; Fontanoz, S.; Lastovica, A.J.; Mandrell, R.E. Extended multilocus sequence typing system for Campylobacter coli, C. lari, C. upsaliensis, and C. helveticus. J. Clin. Microbiol. 2005, 43, 2315–2329. [Google Scholar] [CrossRef] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs Zone Diameters. Version 13.1.2023. Available online: http://www.eucast.org (accessed on 27 July 2023).


{kind=link}
| ST | adk | atpA | glnA | glyA | pgi | pgm | tkt | Number of Isolates | Source Species |
|---|---|---|---|---|---|---|---|---|---|
| 8 | 7 | 1 | 1 | 1 | 1 | 3 | 2 | 3 | Larus michahellis (2) Larus ridibundus (1) |
| 68 | 1 | 6 | 1 | 16 | 1 | 1 | 1 | 4 | Larus michahellis (3) Larus ridibundus (1) |
| 69 | 2 | 1 | 1 | 2 | 76 | 3 | 33 | 1 | Larus michahellis |
| 73 | 6 | 5 | 1 | 1 | 69 | 1 | 6 | 1 | Larus michahellis |
| 77 | 7 | 1 | 1 | 53 | 1 | 3 | 2 | 3 | Larus michahellis |
| 127 | 84 | 6 | 1 | 1 | 4 | 1 | 36 | 1 | Larus michahellis |
| 137 | 90 | 5 | 1 | 77 | 3 | 5 | 6 | 2 | Larus michahellis |
| 158 | 103 | 65 | 1 | 1 | 1 | 3 | 44 | 1 | Larus michahellis |
| 159 | 7 | 57 | 1 | 1 | 4 | 1 | 6 | 1 | Larus cachinnans |
| 160 | 6 | 2 | 1 | 1 | 1 | 1 | 6 | 1 | Larus canus |
| 161 | 103 | 2 | 1 | 1 | 1 | 3 | 44 | 2 | Larus michahellis |
| 165 | 4 | 57 | 1 | 2 | 1 | 1 | 2 | 1 | Larus michahellis |
| 168 | 8 | 6 | 1 | 1 | 1 | 1 | 2 | 1 | Larus cachinnans |
| 222 | 8 | 6 | 1 | 1 | 1 | 1 | 6 | 1 | Larus michahellis |
| 223 | 128 | 6 | 1 | 1 | 1 | 1 | 36 | 1 | Larus michahellis |
| 224 | 8 | 6 | 1 | 1 | 1 | 3 | 124 | 1 | Larus michahellis |
| 225 | 6 | 5 | 1 | 1 | 69 | 1 | 123 | 1 | Larus michahellis |
| 226 | 37 | 57 | 1 | 1 | 2 | 147 | 5 | 1 | Larus michahellis |
| 227 | 128 | 121 | 1 | 1 | 1 | 1 | 2 | 1 | Larus ridibundus |
| 228 | 37 | 4 | 104 | 1 | 1 | 3 | 6 | 1 | Larus ridibundus |
| 229 | 6 | 2 | 1 | 1 | 58 | 3 | 127 | 1 | Larus michahellis |
| 238 | 5 | 6 | 1 | 1 | 1 | 1 | 2 | 1 | Larus michahellis |
| 261 | 34 | 32 | 119 | 25 | 33 | 30 | 49 | 1 | Larus ridibundus |
| 262 | 103 | 2 | 1 | 1 | 184 | 3 | 44 | 1 | Larus michahellis |
| 295 | 8 | 6 | 1 | 1 | 58 | 1 | 2 | 2 | Larus ridibundus |
| 296 | 6 | 4 | 1 | 1 | 69 | 1 | 6 | 3 | Larus michahellis |
| 297 | 8 | 152 | 1 | 1 | 1 | 1 | 2 | 2 | Larus michahellis |
| 298 | 8 | 6 | 120 | 1 | 1 | 1 | 3 | 1 | Larus michahellis |
| 299 | 1 | 57 | 121 | 1 | 4 | 172 | 159 | 1 | Larus michahellis |
| 306 | 8 | 6 | 1 | 1 | 1 | 3 | 44 | 1 | Larus ridibundus |
| 312 | 8 | 6 | 1 | 1 | 1 | 3 | 2 | 3 | Larus michahellis |
| 313 | 84 | 6 | 1 | 1 | 1 | 1 | 36 | 1 | Larus michahellis |
| 314 | 8 | 6 | 1 | 1 | 58 | 119 | 36 | 1 | Larus michahellis |
| 315 | 8 | 1 | 1 | 1 | 1 | 1 | 165 | 2 | Larus michahellis |
| 316 | 103 | 62 | 1 | 1 | 1 | 3 | 44 | 1 | Larus michahellis |
| 317 | 7 | 5 | 1 | 1 | 69 | 1 | 2 | 1 | Larus michahellis |
| 318 | 8 | 2 | 1 | 1 | 1 | 3 | 127 | 1 | Larus michahellis |
| 319 | 6 | 5 | 1 | 53 | 2 | 5 | 6 | 1 | Larus michahellis |
| 320 | 8 | 6 | 1 | 1 | 4 | 1 | 36 | 2 | Larus michahellis |
| 322 | 6 | 5 | 1 | 1 | 190 | 1 | 6 | 1 | Larus michahellis |
| 325 | 8 | 6 | 1 | 53 | 58 | 1 | 2 | 1 | Larus michahellis |
| 326 | 103 | 6 | 1 | 1 | 4 | 1 | 2 | 1 | Larus michahellis |
| 327 | 7 | 1 | 1 | 53 | 1 | 127 | 2 | 1 | Larus michahellis |
| 6 | 5 | 1 | 1 | 69 | 1 | - | 1 | Larus michahellis | |
| - | 6 | 1 | 1 | 1 | 1 | 36 | 1 | Larus michahellis | |
| - | 57 | 1 | 2 | 1 | 1 | 2 | 1 | Larus michahellis | |
| 128 | - | 1 | 1 | 1 | - | 36 | 1 | Larus michahellis |
| mg/L | CHL | ERY | GEN | CIP | TET | ETP | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 512 | 0 | |||||||||||
| 256 | 0 | |||||||||||
| 128 | 0 | |||||||||||
| 64 | 0 | 0 | 1 | |||||||||
| 32 | 0 | 0 | 0 | 0 | ||||||||
| 16 | 1 | 0 | 0 | 0 | 0 | |||||||
| 8 | 0 | 0 | 0 | 5 | 0 | |||||||
| 4 | 1 | 0 | 0 | 27 | 0 | 1 | ||||||
| 2 | 0 | 2 | 11 | 0 | 0 | |||||||
| ≤2 | 41 | |||||||||||
| 1 | 27 | 0 | 1 | 0 | ||||||||
| ≤1 | 43 | |||||||||||
| 0.5 | 12 | 0 | 1 | |||||||||
| ≤0.5 | 41 | |||||||||||
| 0.25 | 0 | 0 | ||||||||||
| ≤0.25 | 2 | |||||||||||
| 0.12 | ||||||||||||
| ≤0.12 | 0 | 41 | ||||||||||
| Antimicrobial Agent | MIC50 (mg/L) | MIC90 (mg/L) | (n/%) S | (n/%) R | EUCAST Epidemiological Cut-Off Value (ECCOF) |
|---|---|---|---|---|---|
| R> | |||||
| CHL | ≤2 | ≤2 | 43/100 | 0/0 | 16 |
| ERY | ≤1 | ≤1 | 43/100 | 0/0 | 8 |
| GEN | 1 | 1 | 43/100 | 0/0 | 2 * |
| CIP | 4 | 8 | 0/0 | 43/100 | 0.5 |
| TET | ≤0.5 | ≤0.5 | 42/97.7 | 1/2.3 | 2 |
| ERTA | ≤0.12 | ≤0.12 | 42/97.7 | 1/2.3 | 0.5 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurinović, L.; Duvnjak, S.; Humski, A.; Ječmenica, B.; Taylor, L.T.; Šimpraga, B.; Krstulović, F.; Zelenika, T.A.; Kompes, G. Genetic Diversity and Resistome Analysis of Campylobacter lari Isolated from Gulls in Croatia. Antibiotics 2023, 12, 1310. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12081310
Jurinović L, Duvnjak S, Humski A, Ječmenica B, Taylor LT, Šimpraga B, Krstulović F, Zelenika TA, Kompes G. Genetic Diversity and Resistome Analysis of Campylobacter lari Isolated from Gulls in Croatia. Antibiotics. 2023; 12(8):1310. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12081310
Chicago/Turabian StyleJurinović, Luka, Sanja Duvnjak, Andrea Humski, Biljana Ječmenica, Louie Thomas Taylor, Borka Šimpraga, Fani Krstulović, Tajana Amšel Zelenika, and Gordan Kompes. 2023. "Genetic Diversity and Resistome Analysis of Campylobacter lari Isolated from Gulls in Croatia" Antibiotics 12, no. 8: 1310. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12081310