Characterization of Endophytic Streptomyces griseorubens MPT42 and Assessment of Antimicrobial Synergistic Interactions of Its Extract and Essential Oil from Host Plant Litsea cubeba

 and
and
Abstract
:1. Introduction
2. Results
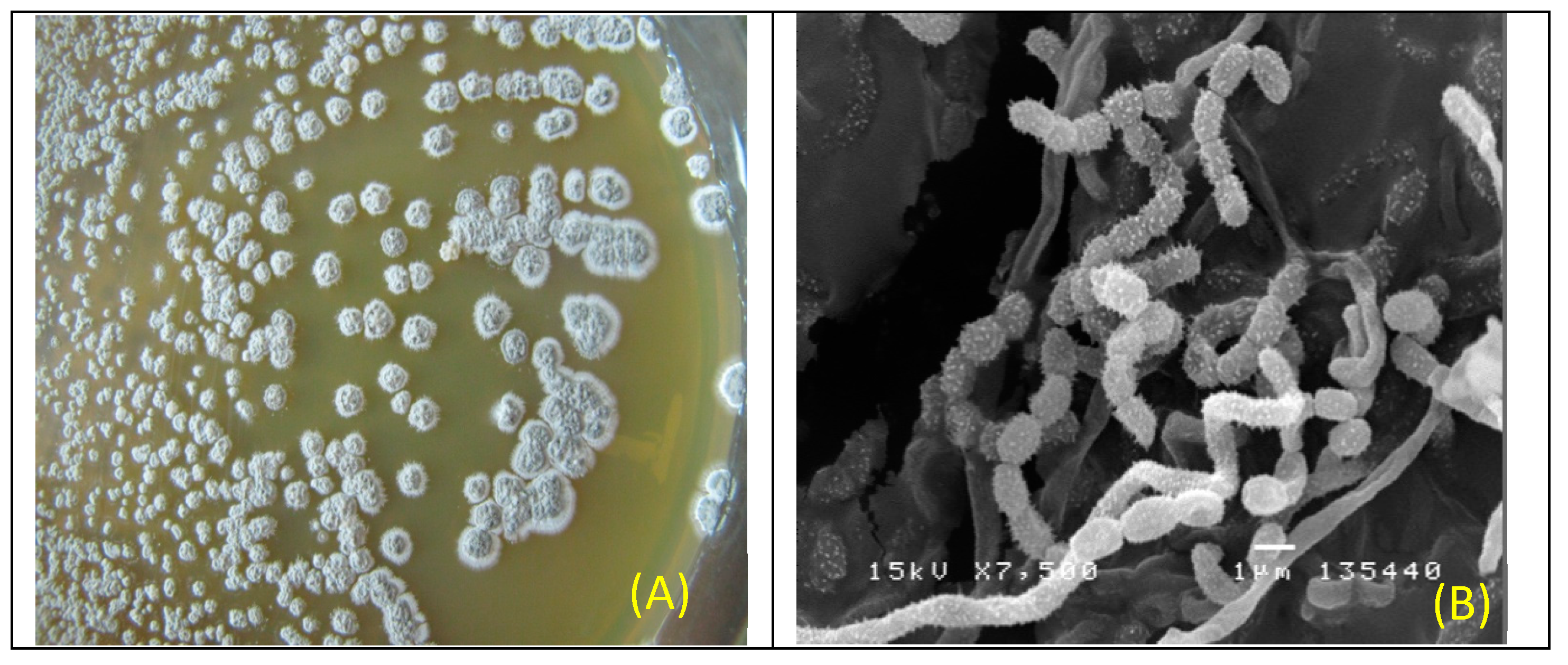
2.1. Identification and Characterization of Antibiotic-Producing Endophytic Actinomycete MPT42
2.2. Antimicrobial Activity of the Crude Ethyl Acetate Extract (CEAE) and Essential Oil (EO)
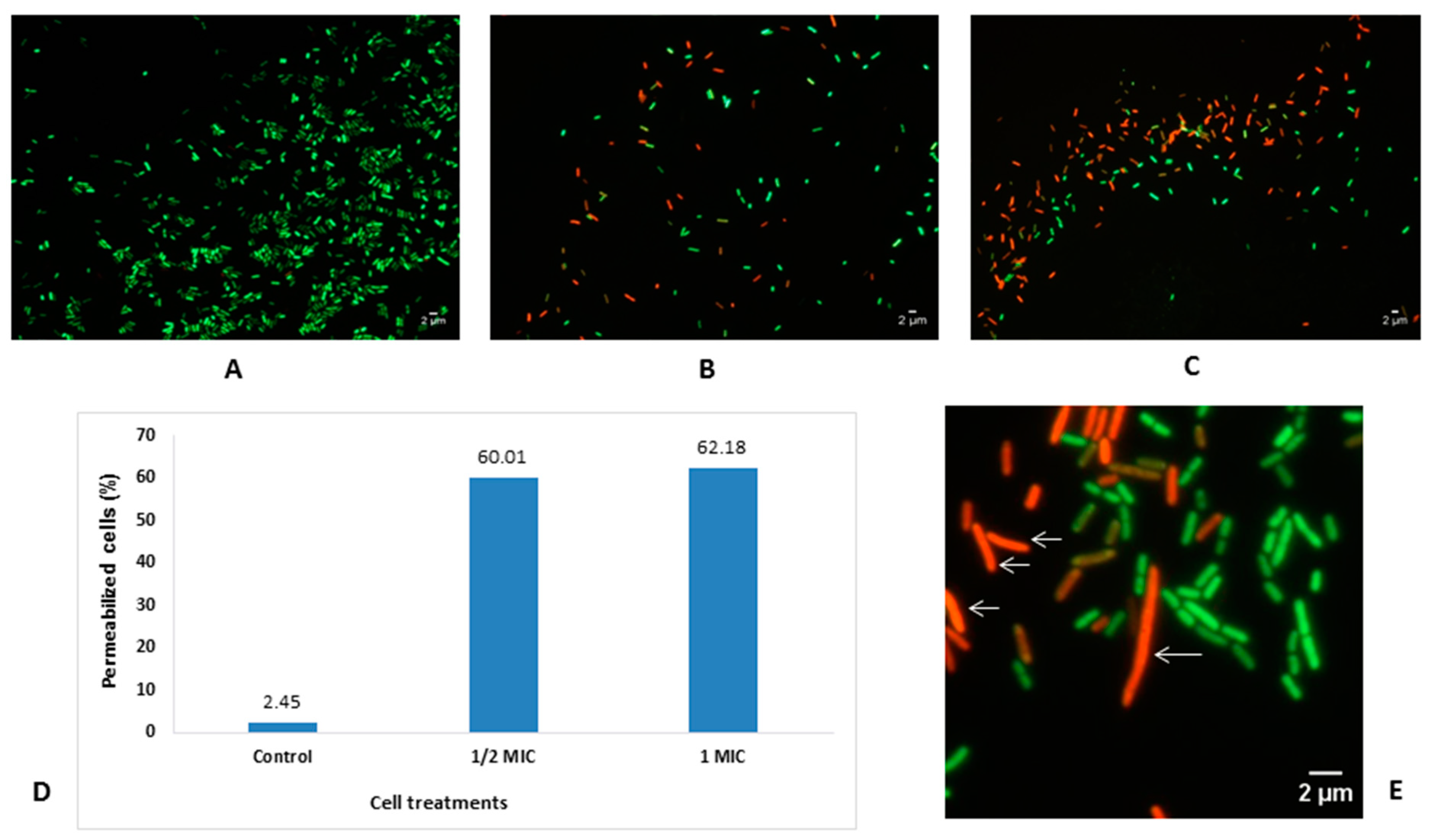
2.3. Effect of the L. cubeba EO on Viability and Cell Morphology of E. coli
2.4. Combined Antimicrobial Effects Against Microbial Strains
3. Discussion
4. Materials and Methods
4.1. Screening for Antibacterial Activity
4.2. Characteristics of the Endophytic Actinomycete MPT42
4.3. Amplification of Secondary Metabolite Biosynthetic Genes and 16S rRNA Encoding Gene
4.4. Preparation of L. cubeba Fruit EO and CEAE From MPT42 Culture
4.5. Determination of Minimum Inhibitory Concentration
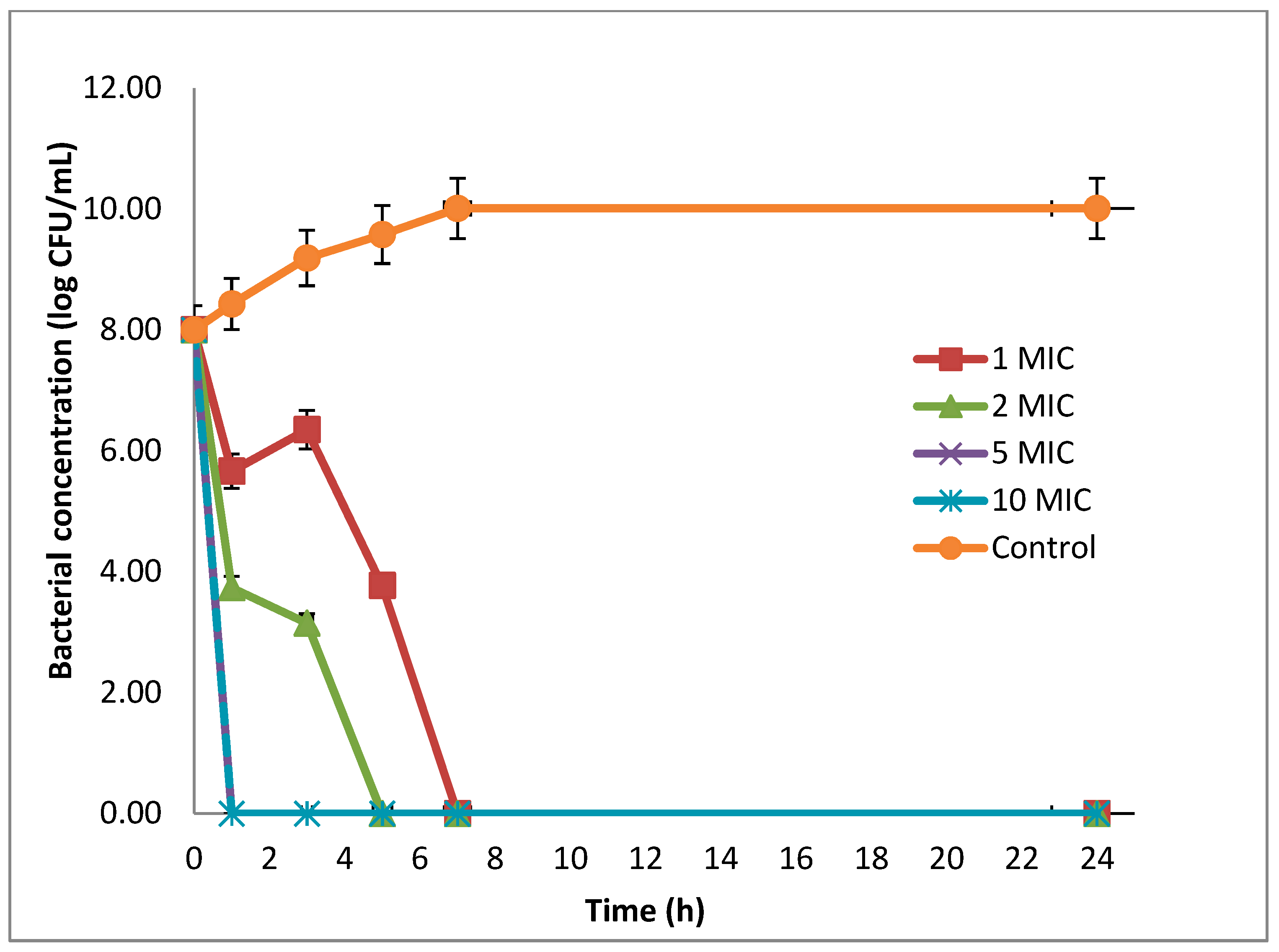
4.6. Time-Killing Assay
4.7. Effects of L. cubeba EO on Bacterial Cell Viability and Morphology
4.8. Microdilution Checkerboard Assays
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.S.; Krishnan, T.; Yiap, B.C.; Hu, C.P.; Chan, K.G.; Lim, S.H. Membrane disruption and anti-quorum sensing effects of synergistic interaction between Lavandula angustifolia (lavender oil) in combination with antibiotic against plasmid-conferred multi-drug-resistant Escherichia coli. J. Appl. Microbiol. 2014, 116, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y. Chemical composition and antibacterial activity of essential oils from different parts of Litsea cubeba. Chem. Biodivers. 2010, 7, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Bhuinya, T.; Singh, P.; Mukherjee, S. Litsea cubeba-Medicinal values-Brief summary. J. Trop. Med. Plants 2010, 11, 179–183. [Google Scholar]
- Chen, Y.; Wang, Y.; Han, X.; Si, L.; Wu, Q.; Lin, L. Biology and chemistry of Litsea cubeba, a promising industrial tree in China. J. Essent. Oil. Res. 2013, 25, 103–111. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Caruso, D.; Lebrun, M.; Nguyen, N.T.; Trinh, T.T.; Meile, J.C.; Chu-Ky, S.; Sarter, S. Antibacterial activity of Litsea cubeba (Lauraceae, May Chang) and its effects on the biological response of common carp Cyprinus carpio challenged with Aeromonas hydrophila. J. Appl. Microbiol. 2016, 121, 341–351. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Vu, T.H.N.; T.T., V.; Phi, Q.-T.; Khieu, T.N.; Sarter, S.; Chu-Ky, S. Antimicrobial activities and interaction effects of Vietnamese Litsea Cubeba (lour.) pers essential oils and its endophytic actinobacteria. Vietnam. J. Sci. Technol. 2016, 54, 234–241. [Google Scholar] [CrossRef]
- Saikia, A.K.; Chetia, D.; D’Arrigo, M.; Smeriglio, A.; Strano, T.; Ruberto, G. Screening of fruit and leaf essential oils of Litsea cubeba Pers. from north-east India – chemical composition and antimicrobial activity. J. Essent. Oil. Res. 2013, 25, 330–338. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Z.-T.; Li, R. Antioxidant activity, free radical scavenging potential and chemical composition of Litsea cubeba essential oil. J. Essent. Oil. Bear. Plant 2012, 15, 134–143. [Google Scholar] [CrossRef]
- Ho, C.L.; Jie-Pinge, O.; Liu, Y.C.; Hung, C.P.; Tsai, M.C.; Liao, P.C.; Wang, E.I.; Chen, Y.L.; Su, Y.C. Compositions and in vitro anticancer activities of the leaf and fruit oils of Litsea cubeba from Taiwan. Nat. Prod. Commun. 2010, 5, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.H.N.; Nguyen, Q.H.; Dinh, T.M.L.; Quach, N.T.; Khieu, T.N.; Hoang, H.; Chu-Ky, S.; Vu, T.T.; Chu, H.H.; Lee, J.; et al. Endophytic actinomycetes associated with Cinnamomum cassia Presl in Hoa Binh province, Vietnam: Distribution, antimicrobial activity and, genetic features. J. Gen. Appl. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.F.; Vollu, R.E.; Jurelevicius, D.; Alviano, D.S.; Alviano, C.S.; Blank, A.F.; Seldin, L. Does the essential oil of Lippia sidoides Cham. (pepper-rosmarin) affect its endophytic microbial community? BMC Microbiol. 2013, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Kang, T.M.; Yuan, J.; Beppler, C.; Nguyen, C.; Mao, Z.; Nguyen, M.Q.; Yeh, P.; Miller, J.H. Synergistic interactions of vancomycin with different antibiotics against Escherichia coli: Trimethoprim and nitrofurantoin display strong synergies with vancomycin against wild-type E. coli. Antimicrob. Agents Chemother. 2015, 59, 276–281. [Google Scholar] [CrossRef]
- Lam, P.N.; Dang, T.T.D.; Vu, T.H.N.; Chu-Ky, S.; Vu, T.T.; Phi, Q.T. Distribution and antimicrobial activity of endophytic actinomycetes isolated from Litsea cubeba (Lour.) Pers. in Northern provinces of Vietnam. Vietnam J. Sci. Technol. 2017, 55, 160. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Medica 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Liu, T.T.; Yang, T.S. Antimicrobial impact of the components of essential oil of Litsea cubeba from Taiwan and antimicrobial activity of the oil in food systems. Int. J. Food. Microbiol. 2012, 156, 68–75. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Meile, J.C.; Lebrun, M.; Caruso, D.; Chu-Ky, S.; Sarter, S. Litsea cubeba leaf essential oil from Vietnam: Chemical diversity and its impacts on antibacterial activity. Lett. Appl. Microbiol. 2018, 66, 207–214. [Google Scholar] [CrossRef]
- Van Vuuren, S.; Suliman, S.; Viljoen, A. The antimicrobial activity of four commercial essential oils in combination with conventional antimicrobials. Lett. Appl. Microbiol. 2009, 48, 440–446. [Google Scholar] [CrossRef]
- Gutierrez, J.; Barry-Ryan, C.; Bourke, P. Antimicrobial activity of plant essential oils using food model media: Efficacy, synergistic potential and interactions with food components. Food Microbiol. 2009, 26, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Christina, A.; Christapher, V.; Bhore, S.J. Endophytic bacteria as a source of novel antibiotics: An overview. Pharmacogn. Rev. 2013, 7, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Li, J.; Chen, H.H.; Zhao, G.Z.; Zhu, W.Y.; Jiang, C.L.; Xu, L.H.; Li, W.J. Isolation, diversity, and antimicrobial activity of rare actinobacteria from medicinal plants of tropical rain forests in Xishuangbanna, China. Appl. Environ. Microbiol. 2009, 75, 6176–6186. [Google Scholar] [CrossRef] [PubMed]
- Al-Askar, A.A.; Rashad, Y.M.; Hafez, E.E.; Abdulkhair, W.M.; Baka, Z.A.; Ghoneem, K.M. Characterization of alkaline protease produced by Streptomyces griseorubens E44G and its possibility for controlling Rhizoctonia root rot disease of corn. Biotechnol. Equip. 2015, 29, 457–462. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nakashima, T. Actinomycetes, an inexhaustible source of naturally occurring antibiotics. Antibiotics 2018, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Bieber, B.; Nuske, J.; Ritzau, M.; Grafe, U. Alnumycin a new naphthoquinone antibiotic produced by an endophytic Streptomyces sp. J. Antibiot. (Tokyo) 1998, 51, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Castillo, U.F.; Strobel, G.A.; Ford, E.J.; Hess, W.M.; Porter, H.; Jensen, J.B.; Albert, H.; Robison, R.; Condron, M.A.M.; Teplow, D.B.; et al. Munumbicins, wide-spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscansa. Microbiology 2002, 148, 2675–2685. [Google Scholar] [CrossRef]
- Inahashi, Y.; Iwatsuki, M.; Ishiyama, A.; Namatame, M.; Nishihara-Tsukashima, A.; Matsumoto, A.; Hirose, T.; Sunazuka, T.; Yamada, H.; Otoguro, K.; et al. Spoxazomicins A-C, novel antitrypanosomal alkaloids produced by an endophytic actinomycete, Streptosporangium oxazolinicum K07-0460(T). J. Antibiot. (Tokyo) 2011, 64, 303–307. [Google Scholar] [CrossRef]
- Li, W.R.; Shi, Q.S.; Liang, Q.; Xie, X.B.; Huang, X.M.; Chen, Y.B. Antibacterial activity and kinetics of Litsea cubeba oil on Escherichia coli. PLoS ONE 2014, 9, e110983. [Google Scholar] [CrossRef]
- Benitez, N.P.; Meléndez León, E.M.; Stashenko, E.E. Eugenol and methyl eugenol chemotypes of essential oil of species Ocimum gratissimum L. and Ocimum campechianum Mill. from Colombia. J. Chromatogr. Sci. 2009, 47, 800–803. [Google Scholar] [CrossRef]
- Ooi, L.S.M.; Li, Y.; Kam, S.-L.; Wang, H.; Wong, E.Y.L.; Ooi, V.E.C. Antimicrobial activities of cinnamon oil and cinnamaldehyde from the Chinese medicinal herb Cinnamomum cassia Blume. Am. J. Chinese Med. 2006, 34, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Trinh, N.T.T.; Dumas, E.; Thanh, M.L.; Degraeve, P.; Amara, C.B.; Gharsallaoui, A.; Oulahal, N. Effect of a Vietnamese Cinnamomum cassia essential oil and its major component trans-cinnamaldehyde on the cell viability, membrane integrity, membrane fluidity, and proton motive force of Listeria innocua. Can. J. Microbiol. 2015, 61, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Bouhdid, S.; Abrini, J.; Amensour, M.; Zhiri, A.; Espuny, M.J.; Manresa, A. Functional and ultrastructural changes in Pseudomonas aeruginosa and Staphylococcus aureus cells induced by Cinnamomum verum essential oil. J. Appl. Microbiol. 2010, 109, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, J.P.; de Azerêdo, G.A.; de Araújo Torres, R.; da Silva Vasconcelos, M.A.; da Conceição, M.L.; de Souza, E.L. Synergies of carvacrol and 1,8-cineole to inhibit bacteria associated with minimally processed vegetables. Int. J. Food. Microbiol. 2012, 154, 145–151. [Google Scholar] [CrossRef]
- Mahboubi, M.; Ghazian Bidgoli, F. Antistaphylococcal activity of Zataria multiflora essential oil and its synergy with vancomycin. Phytomedicine 2010, 17, 548–550. [Google Scholar] [CrossRef]
- Van Vuuren, S.; Viljoen, A. Plant-based antimicrobial studies-methods and approaches to study the interaction between natural products. Planta. Med. 2011, 77, 1168–1182. [Google Scholar] [CrossRef]
- Langeveld, W.T.; Veldhuizen, E.J.; Burt, S.A. Synergy between essential oil components and antibiotics: A review. Crit. Rev. Microbiol. 2014, 40, 76–94. [Google Scholar] [CrossRef]
- Ozbek-Celik, B.; Damar-Celik, D.; Mataraci-Kara, E.; Bozkurt-Guzel, C.; Savage, P.B. Comparative In Vitro Activities of First and Second-Generation Ceragenins Alone and in Combination with Antibiotics Against Multidrug-Resistant Klebsiella pneumoniae Strains. Antibiotics 2019, 8, 130. [Google Scholar] [CrossRef]
- Mgbeahuruike, E.E.; Stålnacke, M.; Vuorela, H.; Holm, Y. Antimicrobial and Synergistic Effects of Commercial Piperine and Piperlongumine in Combination with Conventional Antimicrobials. Antibiotics 2019, 8, 55. [Google Scholar] [CrossRef]
- Adwan, G.; Mhanna, M. Synergistic effects of plant extracts and antibiotics on Staphylococcus aureus strains isolated from clinical specimens. Middle-East J. Sci. Res. 2008, 3, 134–139. [Google Scholar]
- Yeh, P.; Tschumi, A.I.; Kishony, R. Functional classification of drugs by properties of their pairwise interactions. Nat. Genet. 2006, 38, 489. [Google Scholar] [CrossRef] [PubMed]
- Holder, I.A.; Boyce, S.T. Agar well diffusion assay testing of bacterial susceptibility to various antimicrobials in concentrations non-toxic for human cells in culture. Burns 1994, 20, 426–429. [Google Scholar] [CrossRef]
- Shirling, E.T.; Gottlieb, D. Methods for characterization of Streptomyces species 1. Int. J. Syst. Evol. Microbiol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Goodfellow, M.; Kumar, Y. Reclassification of Streptomyces hygroscopicus strains as Streptomyces aldersoniae sp. nov., Streptomyces angustmyceticus sp. nov., comb. nov., Streptomyces ascomycinicus sp. nov., Streptomyces decoyicus sp. nov., comb. nov., Streptomyces milbemycinicus sp. nov. and Streptomyces wellingtoniae sp. Int. J. Syst. Evol. Microbiol. 2010, 60, 769–775. [Google Scholar]
- Vu, H.T.; Nguyen, D.T.; Nguyen, H.Q.; Chu, H.H.; Chu, S.K.; Chau, M.V.; Phi, Q.T. Antimicrobial and Cytotoxic Properties of Bioactive Metabolites Produced by Streptomyces cavourensis YBQ59 Isolated from Cinnamomum cassia Prels in Yen Bai Province of Vietnam. Curr. Microbiol. 2018, 75, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Phi, Q.T.; Park, Y.M.; Seul, K.J.; Ryu, C.M.; Park, S.H.; Kim, J.G.; Ghim, S.Y. Assessment of root-associated paenibacillus polymyxa groups on growth promotion and induced systemic resistance in pepper. J. Microbiol. Biotechnol. 2010, 20, 1605–1613. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Metsa-Ketela, M.; Salo, V.; Halo, L.; Hautala, A.; Hakala, J.; Mantsala, P.; Ylihonko, K. An efficient approach for screening minimal PKS genes from Streptomyces. FEMS Microbiol. Lett. 1999, 180, 1–6. [Google Scholar] [CrossRef]
- Ayuso-Sacido, A.; Genilloud, O. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: Detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 2005, 49, 10–24. [Google Scholar] [CrossRef]
- Salam, N.; Khieu, T.N.; Liu, M.J.; Vu, T.T.; Chu-Ky, S.; Quach, N.T.; Phi, Q.T.; Narsing Rao, M.P.; Fontana, A.; Sarter, S.; et al. Endophytic Actinobacteria Associated with Dracaena cochinchinensis Lour.: Isolation, Diversity, and Their Cytotoxic Activities. Biomed. Res. Int. 2017, 2017, 1308563. [Google Scholar] [CrossRef]
- Visvalingam, J.; Holley, R.A. Temperature-dependent effect of sublethal levels of cinnamaldehyde on viability and morphology of Escherichia coli. J. Appl. Microbiol. 2012, 113, 591–600. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Zone of Inhibition (D ± SD, mm) # |
|---|---|
| Gram-positive bacteria | |
| Listeria innocua ATCC 33090 | 27.5d ± 0.5 |
| Staphylococcus aureus ATCC 25923 | 44.0a ± 2.0 |
| Bacillus cereus ATCC 13061 | 18.0f ± 2.0 |
| Bacillus subtilis ATCC 11778 | 36.3b ± 1.1 |
| MRSE | 28.2d ± 0.5 |
| Gram-negative bacteria | |
| Escherichia coli ATCC 25922 | 32.3c ± 0.4 |
| Salmonella Typhimurium ATCC 14028 | 24.8e ± 0.9 |
| Proteus vulgaris ATCC 49132 | 11.4h ± 0.4 |
| Pseudomonas aeruginosa ATCC 9027 | 17.2g ± 0.9 |
| Enterobacter aerogenes ATCC 13048 | 18.3f ± 0.4 |
| Characteristic | Result | Characteristic | Result |
|---|---|---|---|
| Morphological Characteristics | Fructose | + | |
| Aerial mycelium | Grey and white | Rhamnose | + |
| Substrate mycelium | Faint-brown | Saccharose | − |
| Diffusible pigment | − | Sorbitol | + |
| Spore chain | Spiral | Trehalose | + |
| Spore surface | Spiny | Asparagin | + |
| Spore shape | Oval-shaped | Histidine | − |
| Physiological Properties | Phenylalanin | − | |
| Temperature range for growth | 25–37 °C | Leucin | + |
| Optimum temperature | 35 °C | Tryptophan | + |
| pH range for growth | 6–10 | Arginin | + |
| Optimum pH | 8 | Isoleucin | + |
| NaCl range for growth | 0.5–5% | Valin | + |
| Optimum NaCl | 1% | Methionin | + |
| Biochemical Properties | Lysin | + | |
| Glucose | + | Threonin | + |
| Galactose | + | Cystein | + |
| Mantose | + | Manitol | + |
| Lactose | − | Biosynthetic Genes | |
| Arabinose | + | pks-I | + |
| Glucosamine | + | pks-II | + |
| Myo-inositol | + | nrps | + |
| Bacteria | MIC of Antimicrobial Agents (mean ± SD, µg/mL) | |||
|---|---|---|---|---|
| EO | CEAE | Erythromycin | Vancomycin | |
| Gram-positive bacteria | ||||
| Bacillus cereus ATCC 13061 | 2800 ± 0 | 300 ± 0 | 6.7 ± 2.3 | 1.0 ± 0 |
| Bacillus subtilis ATCC 11778 | 2800 ± 0 | 200 ± 0 | 10.7 ± 4.6 | 2.0 ± 0 |
| Staphylococcus aureus ATCC 25923 | 2800 ± 0 | 80 ± 0 | 0.7 ± 0.3 | 1.0 ± 0 |
| MRSA | 3700 ± 1.6 | 1500 ± 0 | 4.0 ± 0 | 2.0 ± 0 |
| MRSE | 3700 ± 1.6 | 400 ± 0.1 | 8.0 ± 0 | 3.3 ± 1.2 |
| Listeria innocua ATCC 33090 | 1200 ± 0.4 | 600 ± 0 | 16.0 ± 0 | 16.0 ± 0.6 |
| Gram-negative bacteria | ||||
| Aeromonas hydrophila ATCC 35654 | 2800 ± 0 | 600 ± 0 | 128.0 ± 0 | 128.0 ± 0 |
| Escherichia coli ATCC 25922 | 5500 ± 0 | 100 ± 0 | 32.0 ± 0 | 64.0 ± 0 |
| Proteus vulgaris ATCC 49132 | 700 ± 0 | 600 ± 0 | 16.0 ± 0 | 64.0 ± 0 |
| Pseudomonas aeruginosa ATCC 9027 | 2300 ± 0.8 | 400 ± 0.1 | 256.0 ± 0 | 32.0 ± 0 |
| Salmonella Typhimurium ATCC 14028 | 5500 ± 0 | 100 ± 0 | 64.0 ± 0 | 64.0 ± 0 |
| Vibrio parahaemolyticus ATCC 17802 | 5500 ± 0 | 200 ± 0 | 42.7 ± 18.5 | 128.0 ± 0 |
| Bacterial Species | Combined EO and Antimicrobial Agents | |||||
|---|---|---|---|---|---|---|
| CEAE | E | VA | ||||
| MIC (μg/mL) | FIC Index* (mean ± SD) | MIC (μg/mL) | FIC Index * (mean ± SD) | MIC (µg/mL) | FIC Index* (mean ± SD) | |
| Gram-positive Bacteria | ||||||
| B. cereus ATCC 13061 | 20 | 0.31 ± 0.0 (S) | 0.56 | 0.25 ± 0.06 (S) | 1.0 | 1.1 ± 0.04 (I) |
| B. subtilis ATCC 11778 | 50 | 0.31 ± 0.0 (S) | 0.89 | 0.21 ± 0.04 (S) | 0.21 | 0.35 ± 0.04 (S) |
| S. aureus ATCC 25923 | 10 | 0.27 ± 0.04 (S) | 0.04 | 0.19 ± 0.00 (S) | 0.08 | 0.21 ± 0.04 (S) |
| MRSA ATCC 33591 | 380 | 0.58 ± 0.14 (A) | 0.67 | 0.23 ± 0.07 (S) | 0.05 | 0.50 ± 0.0 (A) |
| MRSE ATCC 35984 | 50 | 0.19 ± 0.0 (S) | 2.67 | 0.42 ± 0.13 (S) | 1.65 | 0.58 ± 0.04 (A) |
| L. innocua ATCC 35984 | 40 | 0.19 ± 0.0 (S) | 2.0 | 0.63 ± 0.11 (A) | 3.33 | 0.38 ± 0.0 (S) |
| Gram-negative Bacteria | ||||||
| A. hydrophila ATCC 35654 | 60 | 0.35 ± 0.04 (S) | 42.7 | 0.54 ± 0.07 (A) | 8.0 | 0.31 ± 0.0 (S) |
| E. coli ATCC 25922 | 20 | 0.27 ± 0.04 (S) | 4.0 | 0.23 ± 0.04 (S) | 16.0 | 0.56 ± 0.0 (A) |
| P. vulgaris ATCC 49132 | 60 | 0.60 ± 0.04 (A) | 1.33 | 0.33 ± 0.04 (S) | 26.7 | 0.54 ± 0.04 (A) |
| P. aeruginosa ATCC 9027 | 100 | 0.31 ± 0.0 (S) | 21.3 | 1.08 ± 0.04 (I) | 2.67 | 0.33 ± 0.04 (S) |
| S. Typhimurium ATCC 14028 | 6 | 1.06 ± 0.0 (I) | 32.0 | 0.56 ± 0.0 (A) | 53.3 | 1.83 ± 0.29 (I) |
| V. parahaemolyticus ATCC 17802 | 100 | 0.56 ± 0.0 (A) | 14.2 | 0.58 ± 0.14 (A) | 26.7 | 0.54 ± 0.07 (A) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, Q.H.; Nguyen, H.V.; Vu, T.H.-N.; Chu-Ky, S.; Vu, T.T.; Hoang, H.; Quach, N.T.; Bui, T.L.; Chu, H.H.; Khieu, T.N.; et al. Characterization of Endophytic Streptomyces griseorubens MPT42 and Assessment of Antimicrobial Synergistic Interactions of Its Extract and Essential Oil from Host Plant Litsea cubeba. Antibiotics 2019, 8, 197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040197
Nguyen QH, Nguyen HV, Vu TH-N, Chu-Ky S, Vu TT, Hoang H, Quach NT, Bui TL, Chu HH, Khieu TN, et al. Characterization of Endophytic Streptomyces griseorubens MPT42 and Assessment of Antimicrobial Synergistic Interactions of Its Extract and Essential Oil from Host Plant Litsea cubeba. Antibiotics. 2019; 8(4):197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040197
Chicago/Turabian StyleNguyen, Quang Huy, Hai Van Nguyen, Thi Hanh-Nguyen Vu, Son Chu-Ky, Thu Trang Vu, Ha Hoang, Ngoc Tung Quach, Thi Lien Bui, Hoang Ha Chu, Thi Nhan Khieu, and et al. 2019. "Characterization of Endophytic Streptomyces griseorubens MPT42 and Assessment of Antimicrobial Synergistic Interactions of Its Extract and Essential Oil from Host Plant Litsea cubeba" Antibiotics 8, no. 4: 197. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040197