Simple Carbohydrate Derivatives Diminish the Formation of Biofilm of the Pathogenic Yeast Candida albicans


Abstract
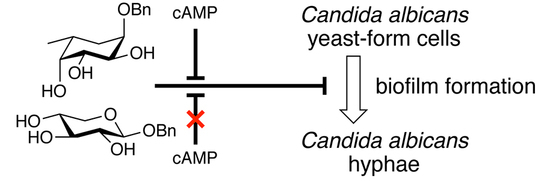
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
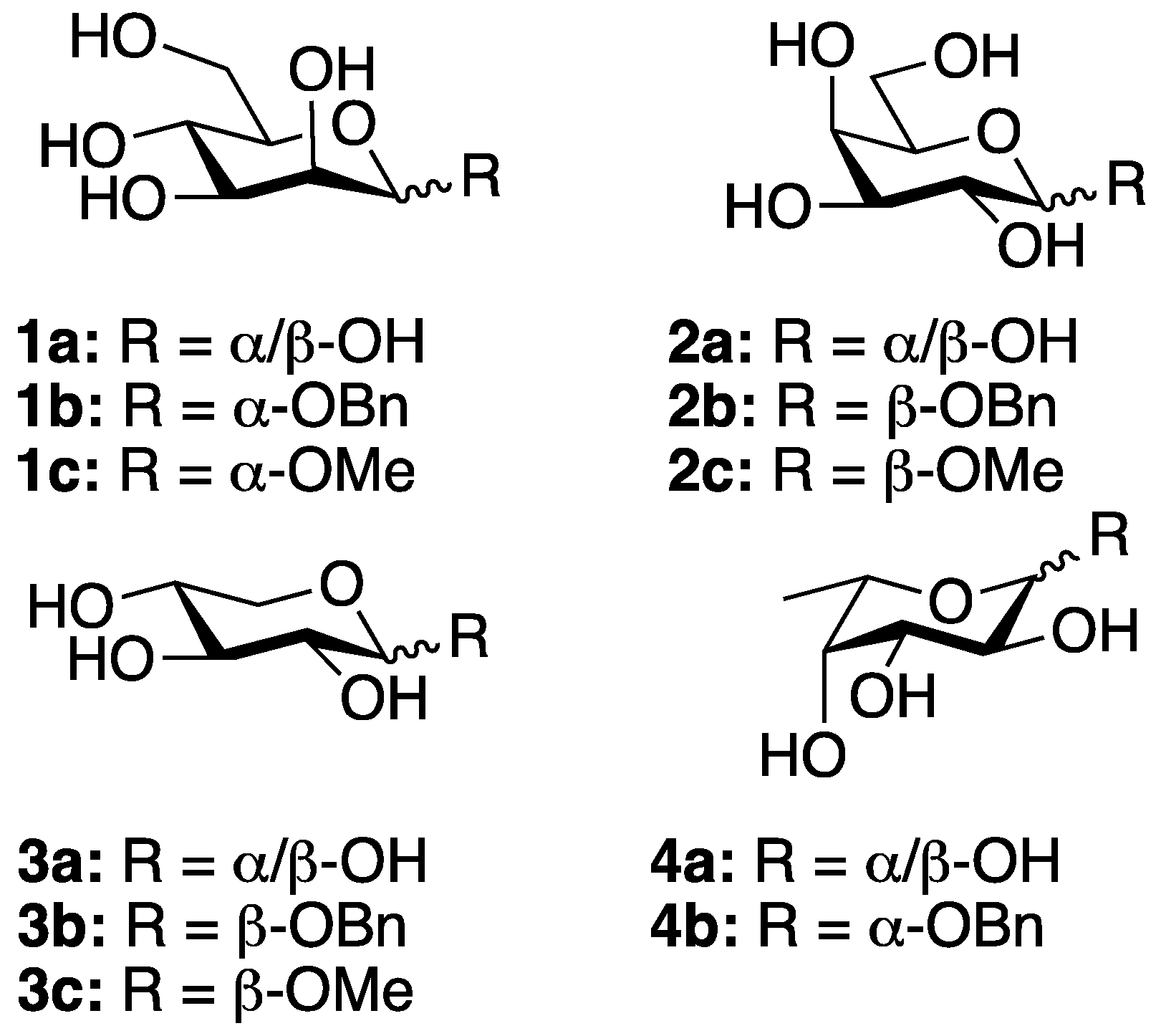
2.1. Carbohydrate Derivatives
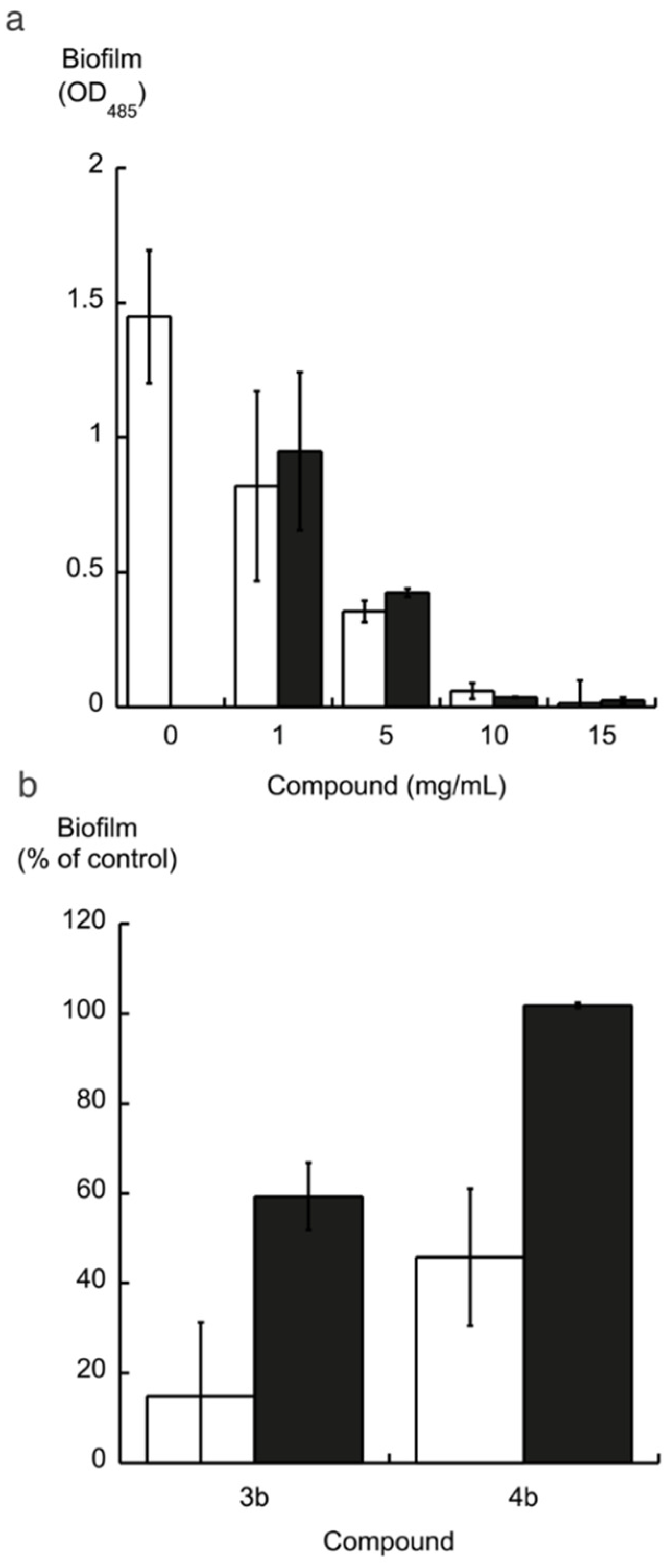
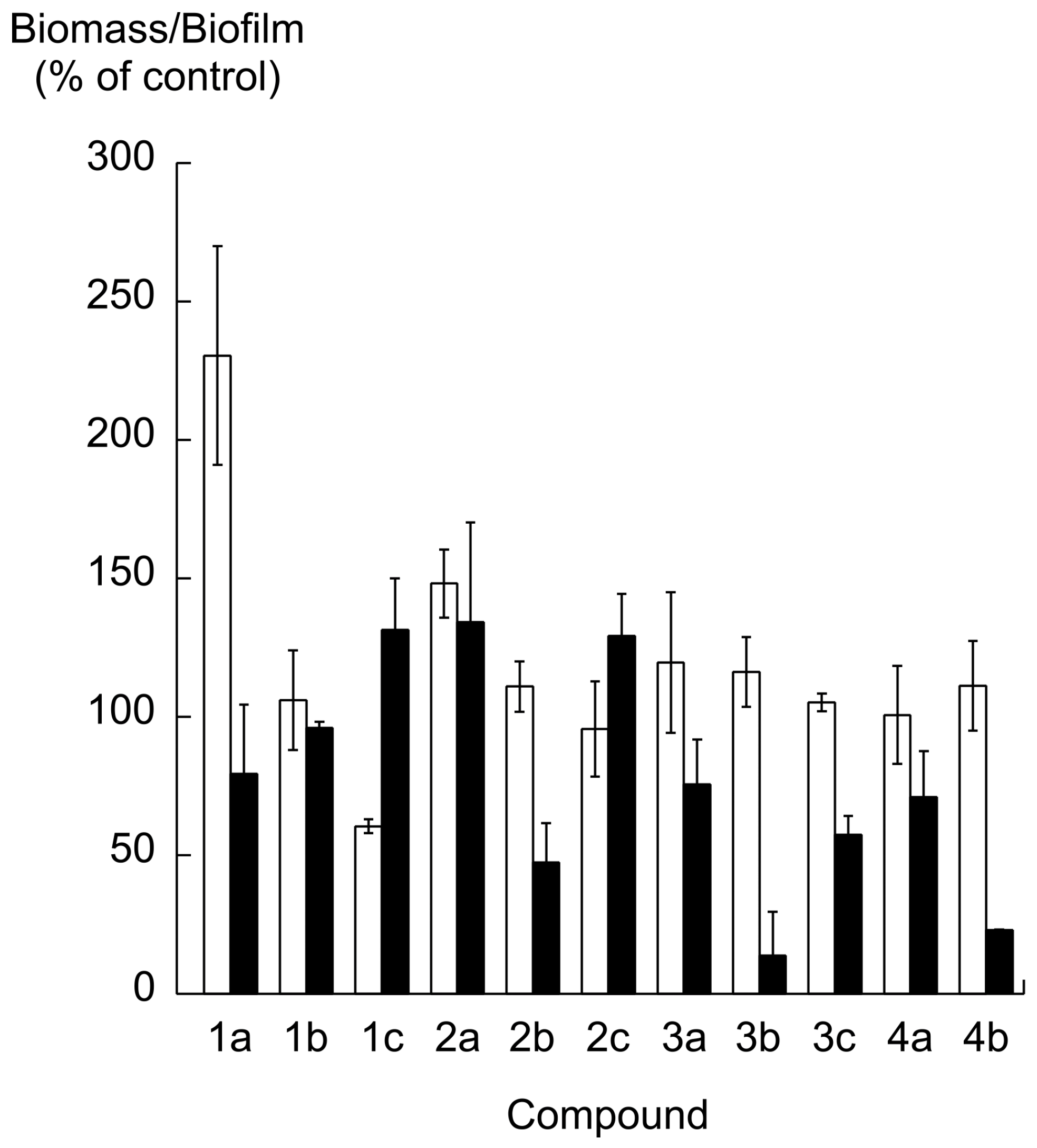
2.2. Biofilm Formation
2.3. Yeast-Hyphae Transition
3. Discussion
4. Materials and Methods
4.1. General Methods, Strains, and Growth Conditions
4.2. Biofilm Assay
4.3. Microfluidics and Microscopy
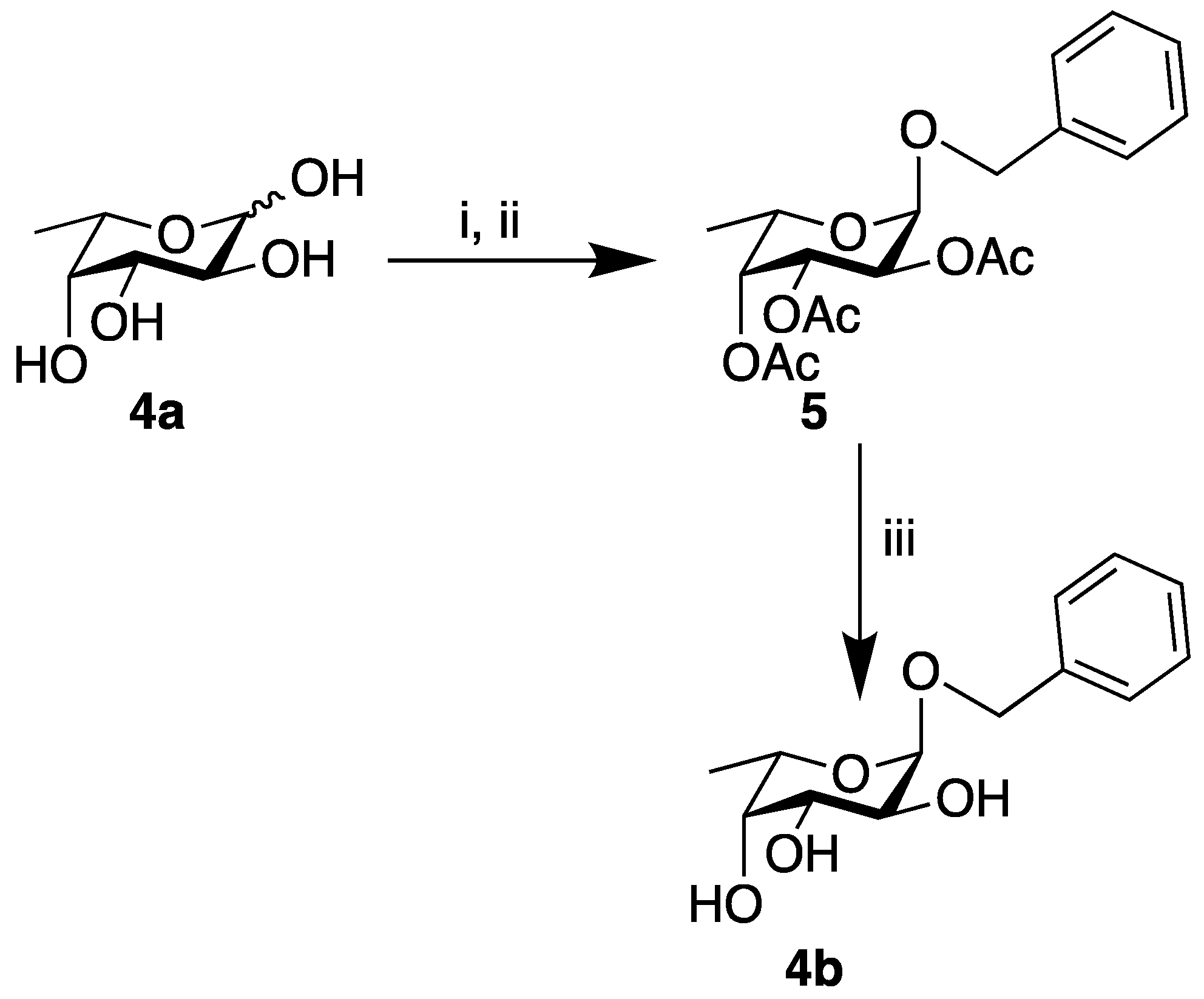
4.4. Synthesis of Benzyl (2,3,4-tri-O-acetyl) α-l-fucopyranoside (5)
4.5. Synthesis of Benzyl α-l-fucopyranoside (4b)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Finkel, J.S.; Mitchell, A.P. Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 2011, 9, 109–118. [Google Scholar] [CrossRef]
- Seneviratne, C.J.; Wang, Y.; Jin, L.; Abiko, Y.; Samaranayake, L.P. Proteomics of drug resistance in Candida glabrata biofilms. Proteomics 2010, 10, 1444–1454. [Google Scholar] [CrossRef]
- Fonseca, E.; Silva, S.; Rodrigues, C.F.; Alves, C.T.; Azeredo, J.; Henriques, M. Effects of fluconazole on Candida glabrata biofilms and its relationship with ABC transporter gene expression. Biofouling 2014, 30, 447–457. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilms: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef] [Green Version]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative role of beta-1,3 glucans in Candida albicans biofilm resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Pradeep Kumar, S.S.; Easwer, H.V.; Maya Nandkumar, A. Multiple drug resistant bacterial biofilms on implanted catheters—A reservoir of infection. J. Assoc. Physicians India 2013, 61, 702–707. [Google Scholar]
- Secor, P.R.; James, G.A.; Fleckman, P.; Olerud, J.E.; McInnerney, K.; Stewart, P.S. Staphylococcus aureus Biofilm and Planktonic cultures differentially impact gene expression, mapk phosphorylation, and cytokine production in human keratinocytes. BMC Microbiol. 2011, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Hofs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: Virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef]
- Potera, C. Forging a link between biofilms and disease. Science 1999, 283, 1837–1839. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef]
- da Silva Dantas, A.; Lee, K.K.; Raziunaite, I.; Schaefer, K.; Wagener, J.; Yadav, B.; Gow, N.A. Cell biology of Candida albicans-host interactions. Curr. Opin. Microbiol. 2016, 34, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Mardh, P.A.; Rodrigues, A.G.; Genc, M.; Novikova, N.; Martinez-de-Oliveira, J.; Guaschino, S. Facts and myths on recurrent vulvovaginal candidosis—A review on epidemiology, clinical manifestations, diagnosis, pathogenesis and therapy. Int. J. STD AIDS 2002, 13, 522–539. [Google Scholar] [CrossRef]
- Low, C.-Y.; Rotstein, C. Emerging fungal infections in immunocompromised patients. F1000 Med. Rep. 2011, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [Green Version]
- Baillie, G.S.; Douglas, L.J. Role of dimorphism in the development of Candida albicans biofilms. J. Med. Microbiol. 1999, 48, 671–679. [Google Scholar] [CrossRef]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Andes, D.; Nett, J.; Oschel, P.; Albrecht, R.; Marchillo, K.; Pitula, A. Development and characterization of an in vivo central venous catheter Candida albicans biofilm model. Infect. Immun. 2004, 72, 6023–6031. [Google Scholar] [CrossRef] [Green Version]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Lowman, D.W.; Greene, R.R.; Bearden, D.W.; Kruppa, M.D.; Pottier, M.; Monteiro, M.A.; Soldatov, D.V.; Ensley, H.E.; Cheng, S.-C.; Netea, M.G.; et al. Novel Structural Features in Candida albicans Hyphal Glucan Provide a Basis for Differential Innate Immune Recognition of Hyphae Versus Yeast. J. Biol. Chem. 2014, 289, 3432–3443. [Google Scholar] [CrossRef] [Green Version]
- Machova, E.; Fiacanova, L.; Cizova, A.; Korcova, J. Mannoproteins from yeast and hyphal form of Candida albicans considerably differ in mannan and protein content. Carbohydr. Res. 2015, 408, 12–17. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.J.; Chen, Y.L. Conserved and Divergent Functions of the cAMP/PKA Signaling Pathway in Candida albicans and Candida tropicalis. J. Fungi 2018, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Van Dijck, P.; Datta, A. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants of Candida albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D.; Myers, P.G.; Kaye, D.; Levison, M.E. Adherence of Candida albicans to human vaginal and buccal epithelial cells. J. Infect. Dis. 1981, 143, 76–82. [Google Scholar] [CrossRef]
- Sandin, R.L.; Rogers, A.L.; Patterson, R.J.; Beneke, E.S. Evidence for mannose-mediated adherence of Candida albicans to human buccal cells in vitro. Infect. Immun. 1982, 35, 79–85. [Google Scholar]
- Sato, M.; Ohshima, T.; Maeda, N.; Ohkubo, C. Inhibitory effect of coated mannan against the adhesion of Candida biofilms to denture base resin. Dent. Mater. J. 2013, 32, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Gharbi, A.; Humblot, V.; Turpin, F.; Pradier, C.M.; Imbert, C.; Berjeaud, J.M. Elaboration of antibiofilm surfaces functionalized with antifungal-cyclodextrin inclusion complexes. FEMS Immunol. Med. Microbiol. 2012, 65, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Enache, E.; Eskandari, T.; Borja, L.; Wadsworth, E.; Hoxter, B.; Calderone, R. Candida albicans adherence to a human oesophageal cell line. Microbiology 1996, 142 Pt 10, 2741–2746. [Google Scholar] [CrossRef] [Green Version]
- Fradin, C.; Jouault, T.; Mallet, A.; Mallet, J.M.; Camus, D.; Sinay, P.; Poulain, D. Beta-1,2-linked oligomannosides inhibit Candida albicans binding to murine macrophage. J. Leukoc. Biol. 1996, 60, 81–87. [Google Scholar] [CrossRef]
- Segal, E.; Kremer, I.; Dayan, D. Inhibition of adherence of Candida albicans to acrylic by a chitin derivative. Eur. J. Epidemiol. 1992, 8, 350–355. [Google Scholar] [CrossRef]
- Gondim, B.L.C.; Castellano, L.R.C.; de Castro, R.D.; Machado, G.; Carlo, H.L.; Valença, A.M.G.; de Carvalho, F.G. Effect of chitosan nanoparticles on the inhibition of Candida spp. biofilm on denture base surface. Arch. Oral Biol. 2018, 94, 99–107. [Google Scholar] [CrossRef]
- Li, Y.; Chang, W.; Zhang, M.; Ying, Z.; Lou, H. Natural product solasodine-3-O-beta-d-glucopyranoside inhibits the virulence factors of Candida albicans. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Vediyappan, G.; Dumontet, V.; Pelissier, F.; d’Enfert, C. Gymnemic acids inhibit hyphal growth and virulence in Candida albicans. PLoS ONE 2013, 8, e74189. [Google Scholar] [CrossRef] [Green Version]
- Zunk, M.; Kiefel, M.J. An efficient synthesis of selectively functionalized d-rhamnose derivatives. Tetrahedron Lett. 2011, 52, 1296–1299. [Google Scholar] [CrossRef] [Green Version]
- Roslund, M.U.; Aitio, O.; Wärnå, J.; Maaheimo, H.; Murzin, D.Y.; Leino, R. Acyl Group Migration and Cleavage in Selectively Protected β-d-Galactopyranosides as Studied by NMR Spectroscopy and Kinetic Calculations. J. Am. Chem. Soc. 2008, 130, 8769–8772. [Google Scholar] [CrossRef]
- Siegbahn, A.; Manner, S.; Persson, A.; Tykesson, E.; Holmqvist, K.; Ochocinska, A.; Roennols, J.; Sundin, A.; Mani, K.; Westergren-Thorsson, G.; et al. Rules for priming and inhibition of glycosaminoglycan biosynthesis; probing the β4GalT7 active site. Chem. Sci. 2014, 5, 3501–3508. [Google Scholar] [CrossRef] [Green Version]
- Bashyal, B.P.; Fleet, G.W.J.; Gough, M.J.; Smith, P.W. Synthesis of the α-Mannosidase inhibitors swainsonine [(1S, 2R, 8R, 8aR)-1,2,8-trihydroxyoctahydroindolizine] and 1,4-dideoxy-1,4-imino-d-mannitol from mannose. Tetrahedron 1987, 43, 3083–3093. [Google Scholar] [CrossRef]
- Wang, H.-Y.L.; O’Doherty, G.A. De novo synthesis of deoxy sugarvia a Wharton rearrangement. Chem. Commun. 2011, 47, 10251–10253. [Google Scholar] [CrossRef]
- Andersen, K.S.; Bojsen, R.; Sørensen, L.G.R.; Nielsen, M.W.; Lisby, M.; Folkesson, A.; Regenberg, B. Genetic basis for Saccharomyces cerevisiae biofilm in liquid medium. G3 2014, 4, 1671–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, W.J.; Seneviratne, J.; Parahitiyawa, N.; Rosa, E.A.; Samaranayake, L.P.; Del Bel Cury, A.A. Improvement of XTT assay performance for studies involving Candida albicans biofilms. Braz. Dent. J. 2008, 19, 364–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veses, V.; Richards, A.; Gow, N.A. Vacuole inheritance regulates cell size and branching frequency of Candida albicans hyphae. Mol. Microbiol. 2009, 71, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Barelle, C.J.; Richard, M.L.; Gaillardin, C.; Gow, N.A.R.; Brown, A.J.P. Candida albicans VAC8 Is Required for Vacuolar Inheritance and Normal Hyphal Branching. Eukaryot. Cell 2006, 5, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Zarnowski, R.; Westler, W.M.; Lacmbouh, G.A.; Marita, J.M.; Bothe, J.R.; Bernhardt, J.; Lounes-Hadj Sahraoui, A.; Fontaine, J.; Sanchez, H.; Hatfield, R.D.; et al. Novel entries in a fungal biofilm matrix encyclopedia. MBio 2014, 5, e01333-14. [Google Scholar] [CrossRef] [Green Version]
- Donohue, D.S.; Ielasi, F.S.; Goossens, K.V.; Willaert, R.G. The N-terminal part of Als1 protein from Candida albicans specifically binds fucose-containing glycans. Mol. Microbiol. 2011, 80, 1667–1679. [Google Scholar] [CrossRef]
- Bojsen, R.K.; Andersen, K.S.; Regenberg, B. Saccharomyces cerevisiae—A model to uncover molecular mechanisms for yeast biofilm biology. FEMS Immunol. Med. Microbiol. 2012, 65, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi—A review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes Dev. 2006, 20, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Fujita, M.; Feng, Q.; Clardy, J.; Fink, G.R. Tyrosol is a quorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 5048–5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Saville, S.P.; Wickes, B.L.; Lopez-Ribot, J.L. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl. Environ. Microbiol. 2002, 68, 5459–5463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machida, K.; Tanaka, T.; Yano, Y.; Otani, S.; Taniguchi, M. Farnesol-induced growth inhibition in Saccharomyces cerevisiae by a cell cycle mechanism. Microbiology 1999, 145 Pt 2, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C.; Brown, A.J.P.; Gow, N.A.R. Candida albicans genome sequence: A platform for genomics in the absence of genetics. Genome Biol. 2004, 5, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Fujarte, I.; Lopez-Romero, E.; Reyna-Lopez, G.E.; Martinez-Gamez, M.A.; Vega-Gonzalez, A.; Cuellar-Cruz, M. Influence of culture media on biofilm formation by Candida species and response of sessile cells to antifungals and oxidative stress. Biomed Res. Int. 2015, 2015, 783639. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishchuk, O.P.; Sterner, O.; Ellervik, U.; Manner, S. Simple Carbohydrate Derivatives Diminish the Formation of Biofilm of the Pathogenic Yeast Candida albicans. Antibiotics 2020, 9, 10. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010010
Ishchuk OP, Sterner O, Ellervik U, Manner S. Simple Carbohydrate Derivatives Diminish the Formation of Biofilm of the Pathogenic Yeast Candida albicans. Antibiotics. 2020; 9(1):10. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010010
Chicago/Turabian StyleIshchuk, Olena P., Olov Sterner, Ulf Ellervik, and Sophie Manner. 2020. "Simple Carbohydrate Derivatives Diminish the Formation of Biofilm of the Pathogenic Yeast Candida albicans" Antibiotics 9, no. 1: 10. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9010010