
Evaluation of the Antifungal Activity of Bacillus amyloliquefaciens and B. velezensis and Characterization of the Bioactive Secondary Metabolites Produced against Plant Pathogenic Fungi

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials Collection
2.2. Plant Segments Sterilization and Endophytes Isolation
2.3. Isolation of Pathogenic Fungi
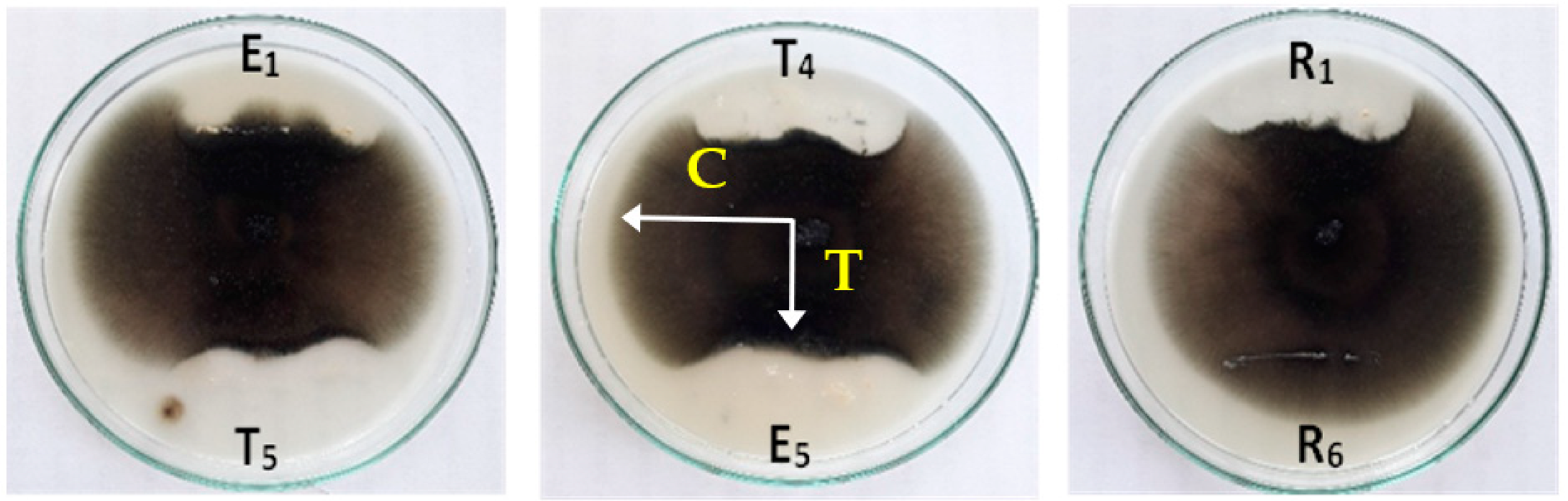
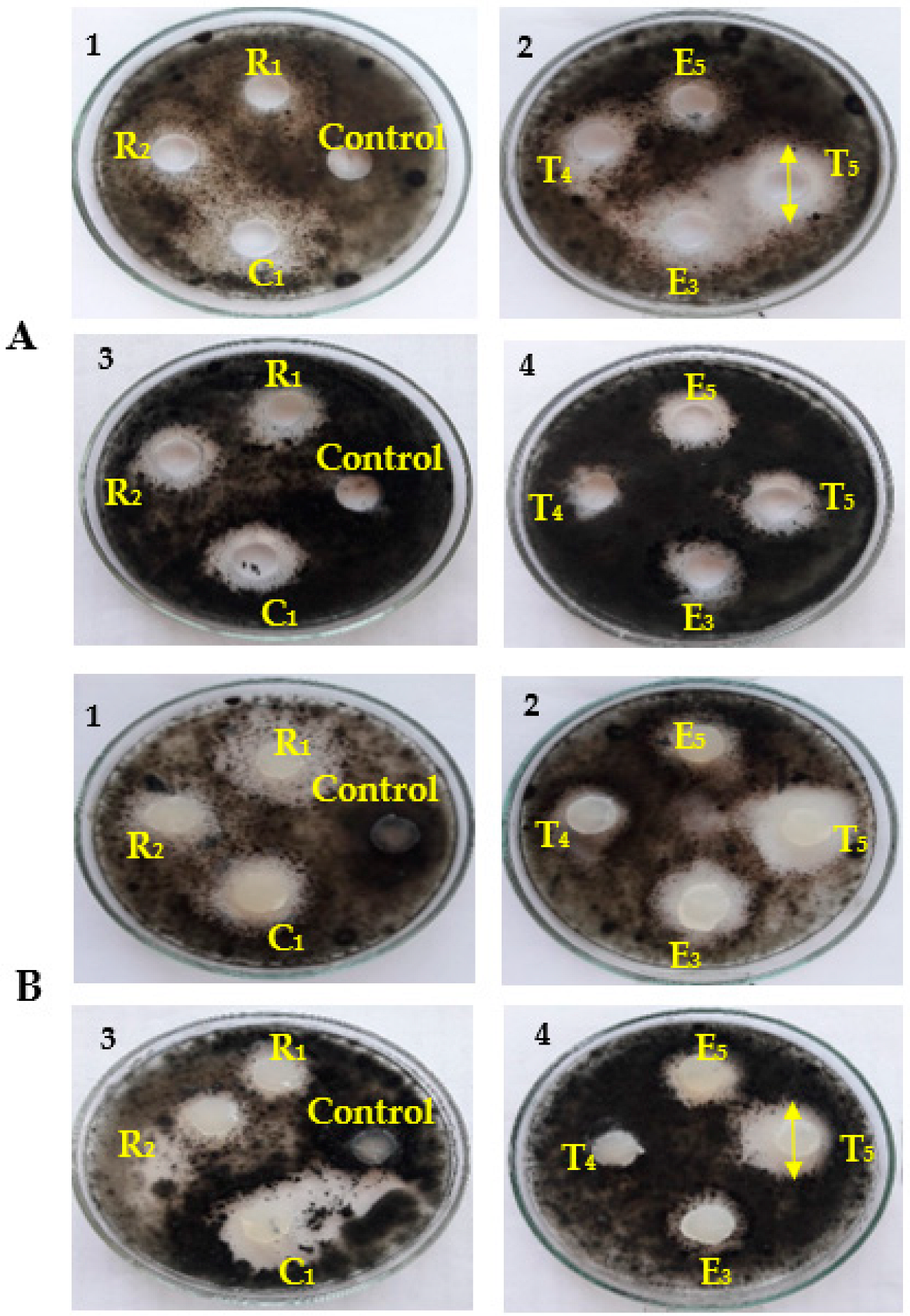
2.4. Primary Screening of the Antagonist Activity of Bacterial Isolates in Vitro
2.5. Evaluation of the Antifungal Activity of Endophytic Bacteria
2.6. Morphological and Biochemical Characteristics of the Antagonistic Bacteria
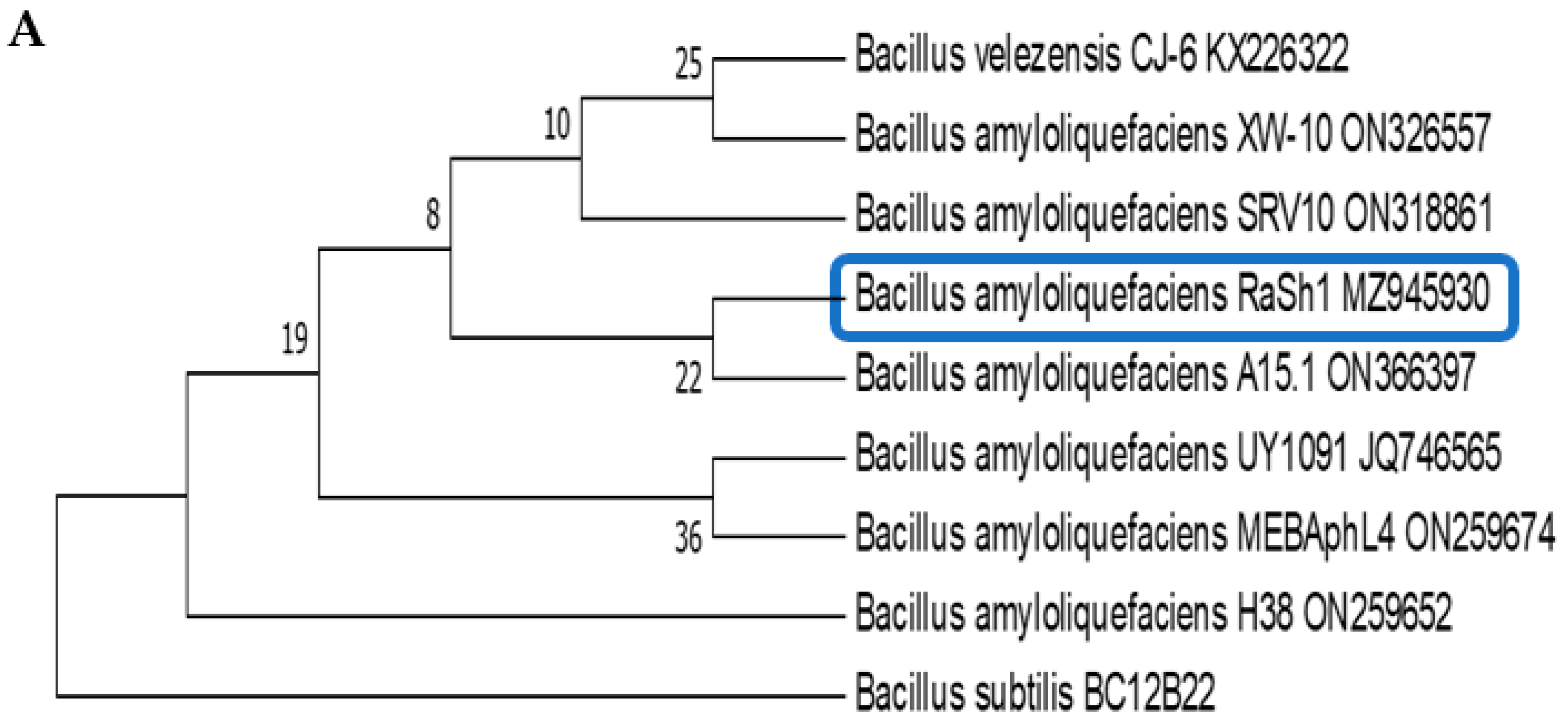
2.7. Molecular Characterization of Bacterial Isolates by Partial Sequencing of 16S rDNA
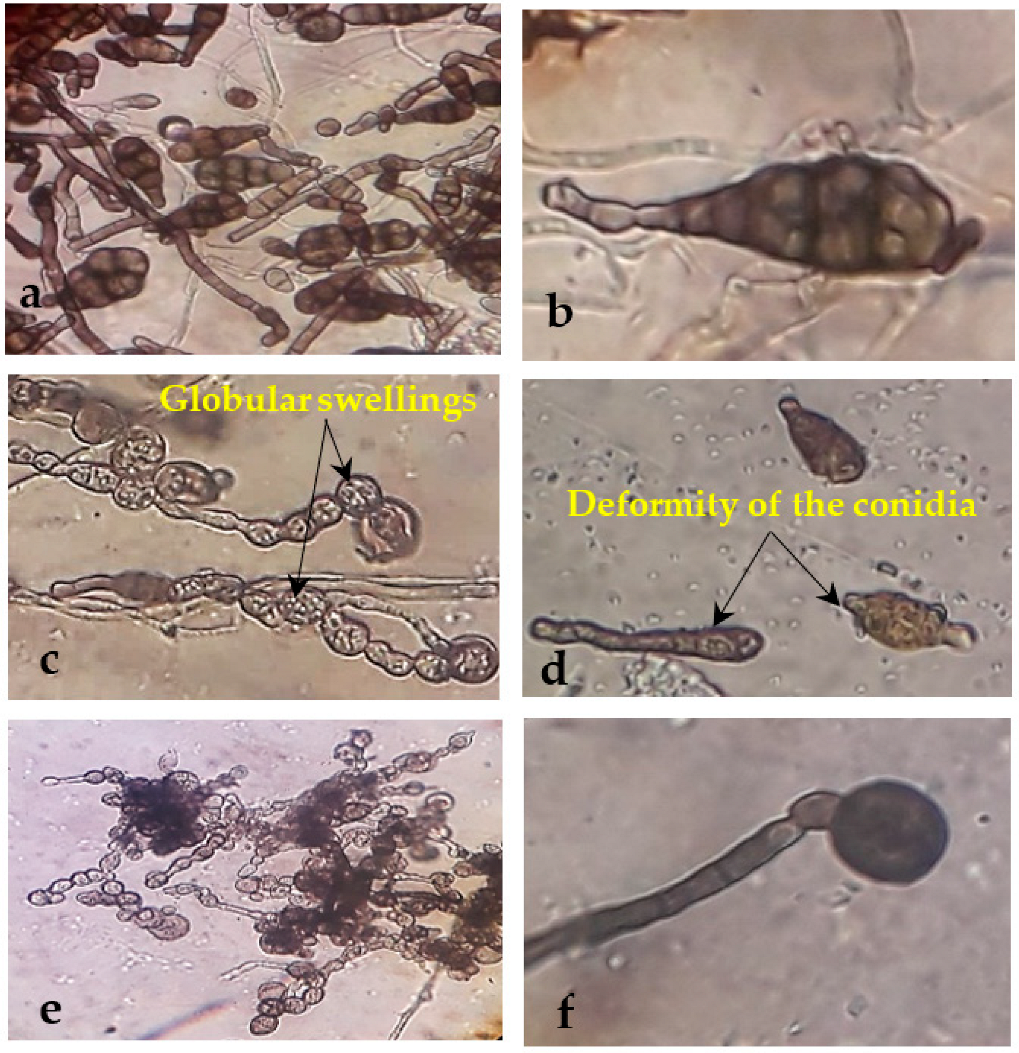
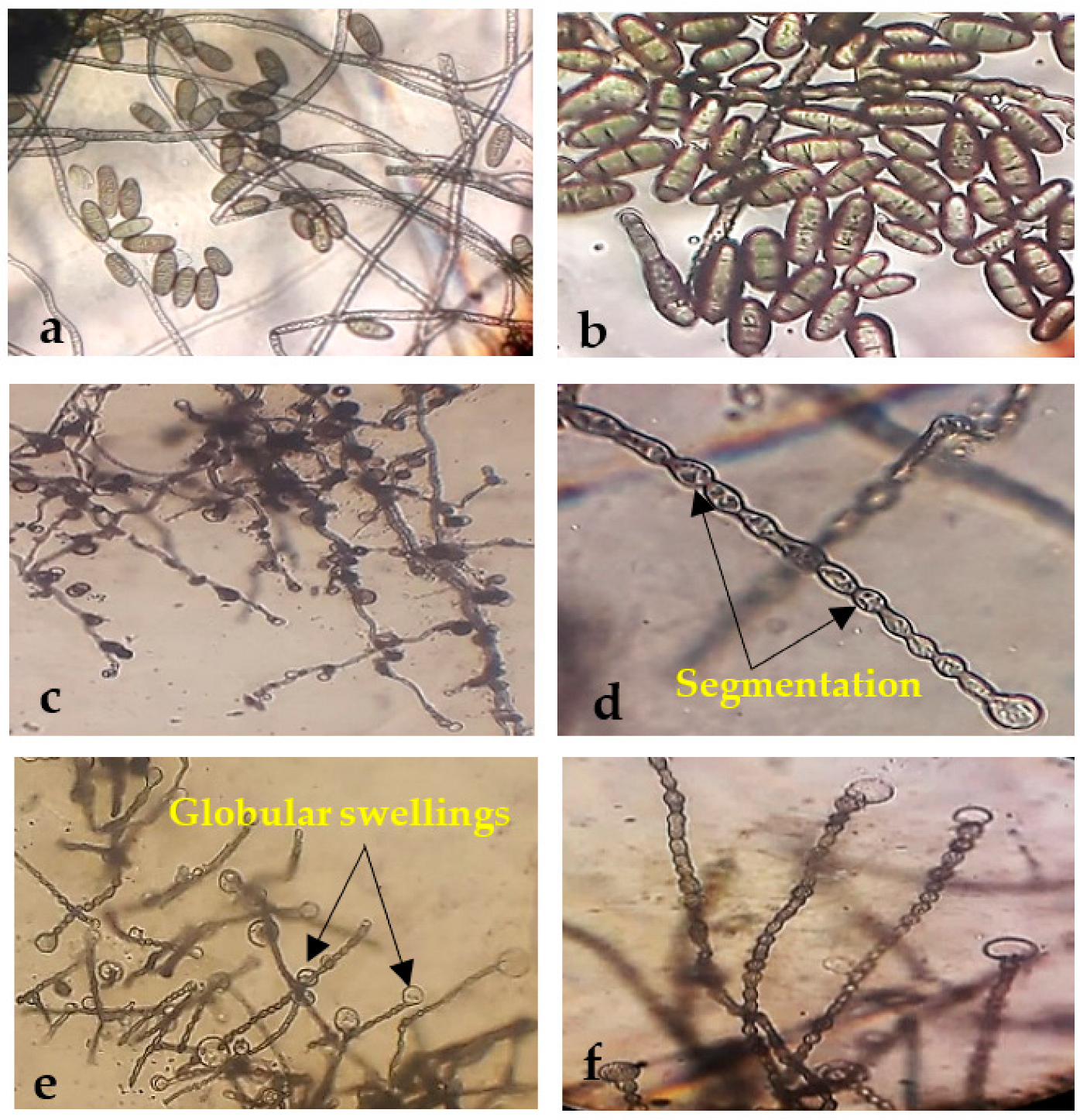
2.8. Morphological Abnormalities in the Alternaria sp. and Helminthosporium sp. Hyphae due to the Antagonistic Effects of Endophytic Bacterial
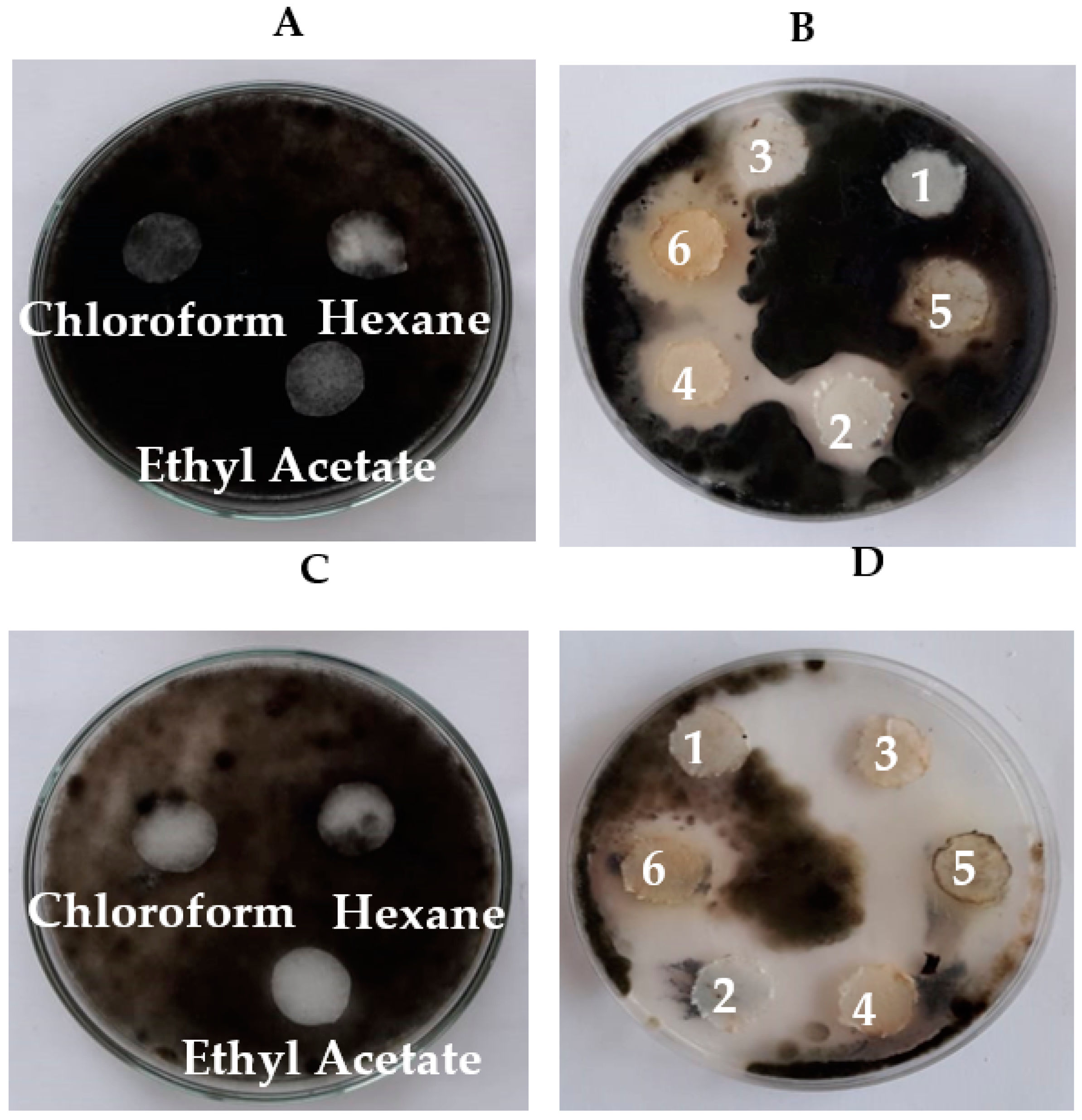
2.9. Preparation of Antifungal Bacterial Crude Extracts using Different Solvents
2.10. Gas Chromatography and Mass Spectrometry (GC–MS)
2.11. In Vivo Evaluation of B. amyloliquefaciens Effects against Alternaria sp.-Infected Pepper Plants under Greenhouse Conditions
2.11.1. B. amyloliquefaciens Inoculum Preparation
2.11.2. Pot Experiment for Cultivation of Pepper Seedlings
2.11.3. Determination of Morphological Parameters
2.11.4. Assessment of Disease Incidence (DI)
2.12. Statistical Analysis
3. Results
3.1. Antifungal Activity of Cell-Free Culture and Discs of Endophytic Bacteria
3.2. Identification of Endophytic Bacterial Isolates
3.3. Morphological Changes under the Light Microscope
3.4. Bioassay and Biological Activity of the Crude Extracts of Endophytic Bacterial Strains
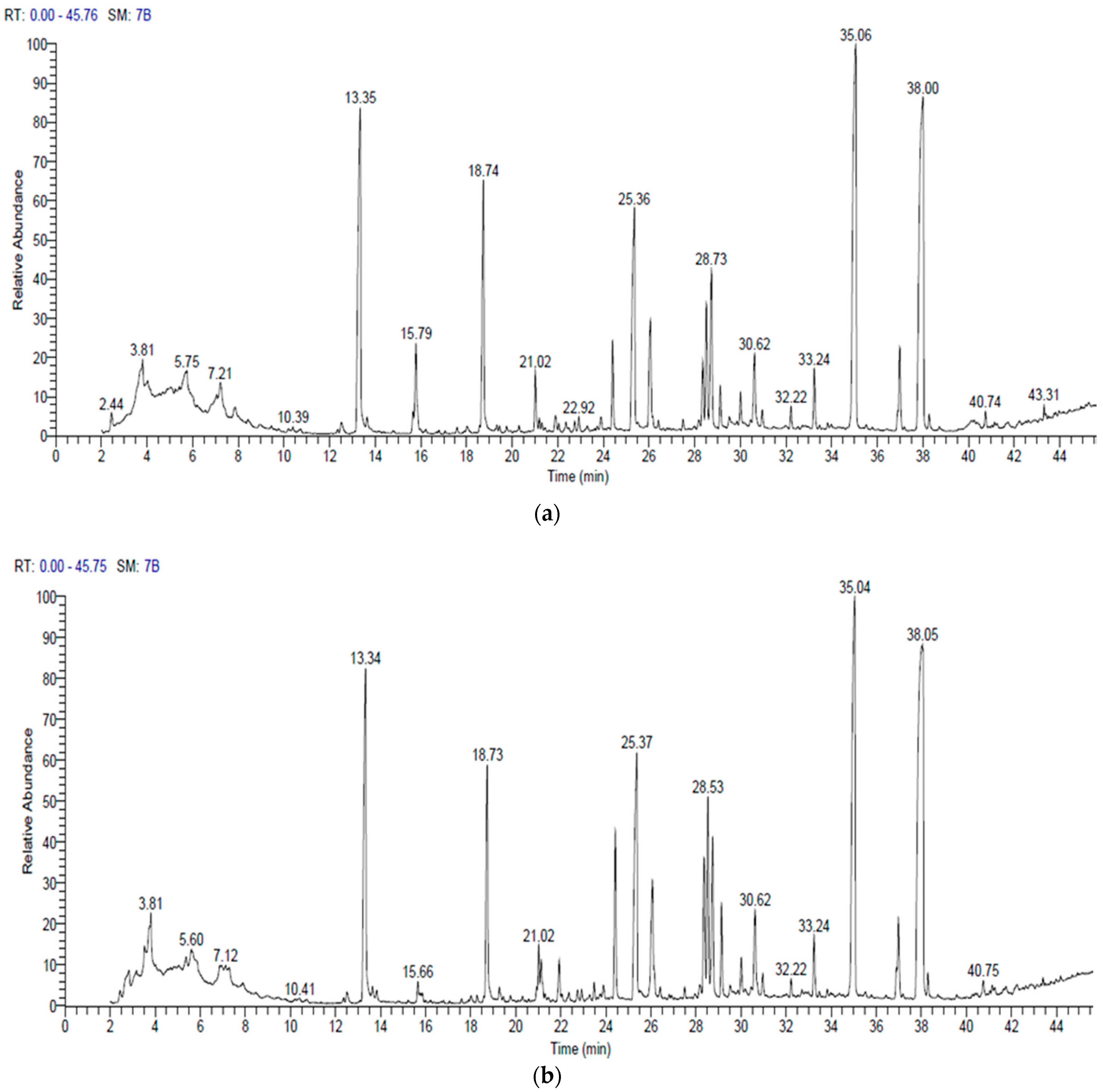
3.5. Gas Chromatography and Mass Spectrometry (GC–MS)
3.6. In Vivo Evaluation of B. amyloliquefaciens Effects against Alternaria sp. Infected Pepper Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal plant ecosystem, health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Dadrasnia, M.; Usman, M.; Omar, R.; Ismail, S.; Abdullah, R. Potential use of Bacillus genus to control of bananas diseases: Approaches toward high yield production and sustainable management. J. King Saud Univ. 2020, 32, 2336–2342. [Google Scholar] [CrossRef]
- Reda, F.M.; Hassanein, W.A.; Sherief, E.A.H.; Moabed, S. Toxicological studies and field applications of a new Bacillus thuringiensis isolate (Bt1) and two chemical pesticides on Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Egypt. J. Biol. Pest Control 2016, 26, 229–236. [Google Scholar]
- Deguine, J.P.; Aubertot, J.N.; Flor, R.J.; Lescourret, F.; Wyckhuys, K.A.G.; Ratnadass, A. Integrated pest management: Good intentions, hard realities. A review. Agron. Sustain. Dev. 2021, 41, 38. [Google Scholar] [CrossRef]
- Rivera-Méndez, W.; Obregón, M.; Morán-Diez, M.E.; Hermosa, R.; Monte, E. Trichoderma asperellum biocontrol activity and induction of systemic defenses against Sclerotium cepivorum in onion plants under tropical climate conditions. Biol. Control 2020, 141, 104145. [Google Scholar] [CrossRef]
- Al-Nadabi, H.H.; Al-Buraiki, N.S.; Al-Nabhani, A.A.; Maharachchikumbura, S.N.; Velazhahan, R.; Al-Sadi, A.M. In vitro antifungal activity of endophytic bacteria isolated from date palm (Phoenix doctylifera L.) against fungal pathogens causing leaf spot of date palm. Egypt. J. Biol. Pest Control 2021, 31, 65. [Google Scholar] [CrossRef]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef]
- Mohamad, O.; Li, L.; Ma, J.; Hatab, S.; Xu, L.; Guo, J.W.; Rasulov, B.A.; Liu, Y.; Hedlund, B.P.; Li, W.J. Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tao, Y.; Zhao, S.; Yin, X.; Chen, J.; Wang, M.; Cai, Y.; Niu, Q. A novel peroxiredoxin from the antagonistic endophytic Bacterium Enterobacter Sp. V1 contributes to cotton resistance against Verticillium Dahliae. Plant Soil 2020, 454, 395–409. [Google Scholar] [CrossRef]
- Kaul, S.; Sharma, T.K.; Dhar, M. “Omics” Tools for better understanding the plant-endophyte interactions. Front. Plant Sci. 2016, 7, 955. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, D.J.; Goswami, G.; Gautom, T.; Parveen, A.; Das, P.; Barooah, M.; Boro, R.C. Lipopeptide mediated biocontrol activity of endophytic Bacillus subtilis against fungal phytopathogens. BMC Microbiol. 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kaushik, N.; Sharma, A.; Bajaj, A.; Rasane, M.; Shouche, Y.S.; Marzouk, T.; Djébali, N. Screening of tomato seed bacterial endophytes for antifungal activity reveals lipopeptide producing Bacillus siamensis Strain NKIT9 as a potential bio-control agent. Front. Microbiol. 2021, 12, 609482. [Google Scholar] [CrossRef] [PubMed]
- Fadiji, A.E.; Babalola, O.O. Exploring the potentialities of beneficial endophytes for improved plant growth. Saudi J. Biol. Sci. 2020, 27, 3622–3633. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Dey, U. An overview of fungal and bacterial biopesticides to control plant pathogens/diseases. Afr. J. Microbiol. Res. 2014, 8, 1749–1762. [Google Scholar]
- Amaresan, N.; Jayakumar, V.; Krishna, K.; Thajuddin, N. Endophytic bacteria from tomato and chilli, their diversity and antagonistic potential against Ralstonia solanacearum. Arch. Phytopathol. Plant Prot. 2011, 45, 344–355. [Google Scholar] [CrossRef]
- Selim, H.M.M.; Gomaa, N.M.; Essa, A.M.M. Antagonistic Effect of the Endophytic Bacteria and against Some Phytopathogens. Egypt. J. Bot. 2016, 56, 613–626. [Google Scholar]
- Kazerooni, E.A.; Maharachchikumbura, S.S.N.; Al-Sadi, A.M.; Kang, S.-M.; Yun, B.-W.; Lee, I.-J. Biocontrol Potential of Bacillus amyloliquefaciens against Botrytis pelargonii and Alternaria alternata on Capsicum annuum. J. Fungi 2021, 7, 472. [Google Scholar] [CrossRef]
- Rashad, Y.M.; Abbas, M.A.; Soliman, H.M.; Abdel-Fattah, G.G.; Abdel-Fattah, G.M. Synergy between endophytic Bacillus amyloliquefaciens GGA and arbuscular mycorrhizal fungi induces plant defense responses against white rot of garlic and improves host plant growth. Phytopathol. Mediterr. 2020, 59, 169–186. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Li, C.; Ma, Y. Antifungal and plant growth promotion activity of volatile organic compounds produced by Bacillus amyloliquefaciens. Microbiol. Open 2019, 8, e813. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, S.P.J.; Chintagunta, A.D.; Agarwal, D.K.; Pal, G.; Singh, A.N.; Simal-Gandara, J. Biocontrol potential of Pseudomonas stutzeri endophyte from Withania somnifera (Ashwagandha) seed extract against pathogenic Fusarium oxysporum and Rhizoctonia solani. Arch. Phytopathol. Plant Prot. 2021, 55, 1–18. [Google Scholar] [CrossRef]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Samad Tahir, H.A.; Gu, Q.; Wu, H.; Gao, X. Volatile compounds of endophytic Bacillus spp. have biocontrol activity against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reda, F.M.; Hassanein, W.A.; Moabed, S. Potential exploitation of Bacillus flexus biofilm against the cowpea weevil, Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Egypt. J. Biol. Pest Control 2020, 30, 18. [Google Scholar] [CrossRef]
- Hassanein, W.A.; Reda, F.M.; Moabed, S.; El Shafiey, S.N. Insecticidal impacts of Bacillus flexus S13 biofilm ‘extracellular matrix’ on cowpea weevil, Callosobruchus maculatus. Biocatal. Agric. Biotechnol. 2021, 31, 101898. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Endophytic fungi from Nerium oleander L (Apocynaceae): Main constituents and antioxidant activity. World J. Microbiol. Biotechnol. 2007, 23, 1253–1263. [Google Scholar] [CrossRef]
- Al-Hussini, H.S.; Al-Rawahi, A.Y.; Al-Marhoon, A.A.; Al-Abri, S.A.; Al-Mahmooli, I.H.; Al-Sadi, A.M.; Velazhahan, R. Biological control of damping-off of tomato caused by Pythium aphanidermatum by using native antagonistic rhizobacteria isolated from Omani soil. J. Plant Pathol. 2019, 101, 315–322. [Google Scholar] [CrossRef]
- Choi, Y.W.; Hyde, K.D.; Ho, W.H. Single spore isolation of fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- Ellis, M.B. Dematiaceus Hyphomycetes; Commonwealth Mycological Institute: Surrey, UK, 1971; 608p. [Google Scholar]
- Archana, T.; Rajendran, L.; Manoranjitham, S.K. Culture-dependent analysis of seed bacterial endophyte, Pseudomonas spp. EGN 1 against the stem rot disease (Sclerotium rolfsii Sacc.) in groundnut. Egypt. J. Biol. Pest Control 2020, 30, 119. [Google Scholar] [CrossRef]
- Pandey, D.K.; Tripathi, N.N.; Tripathi, R.D. Fungitoxic and phytotoxic properties of essential oil of Hyptis suaveolens. Zeitschrift fuer Pflanzenkrankheiten und Pflanzenschutz 1982, 89, 344–349. [Google Scholar]
- Paul, N.C.; Kim, W.K.; Woo, S.K.; Park, M.S.; Yu, S.H. The fungal endophytes in roots of Aralia species and their antifungal activity. Plant Pathol. J. 2007, 23, 287–294. [Google Scholar] [CrossRef]
- O’Connor, E.B.; O’Riordan, B.; Morgan, S.M.; Whelton, H.; O’Mullane, D.M.; Ross, R.P. A lacticin 3147 enriched food ingredient reduces Streptococcus mutans isolated from the human oral cavity in saliva. J. Appl. Microbiol. 2006, 100, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, P.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods for General and Molecular Bacteriology; ASM: Washington, DC, USA, 1994. [Google Scholar]
- Bergey, D.H.; Holt, J.G. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Bergey’s Manual of Systematic Bacteriology; Williams & Wilkins Co.: Baltimore, MD, USA, 1994. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 2001. [Google Scholar]
- Srivastava, S.; Singh, V.; Kumar, V.; Verma, P.C.; Srivastava, R.; Basu, V.; Gupta, V.; Rawat, A.K. Identification of regulatory elements in 16S rRNA gene of Acinetobacter species isolated from water sample. Bioinformation 2008, 3, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Sariah, M. Potential of Bacillus spp. as biocontrol agent for anthracnose fruit rot of chili malays. Appl. Biol. 1994, 23, 1193–1201. [Google Scholar]
- Muzzamal, H.; Sarwar, R.; Sajid, I.; Hasnain, S. Isolation, identification and screening of endophytic bacteria antagonistic to biofilm formers. Pak. J. Zool. 2012, 44, 249–258. [Google Scholar]
- Nandhini, U.; Kumari, L.; Sangareshwari, S. Gas Chromatography-Mass Spectrometry analysis of bioactive constituents from the marine Streptomyces. Asian J. Pharm. Clin. Res. 2015, 8, 244–246. [Google Scholar]
- Senthilkumar, N.; Murugesan, S.; Babu, D.S.; Rajeshkannan, C. GC-MS analysis of the extract of endophytic fungus, Phomopsis sp. isolated from tropical tree species of India, Tectona grandis L. Int. J. Innov. Res. Sci. Eng. Technol. 2014, 3, 10176–10179. [Google Scholar]
- Widnyana, I.K.; Javandira, C. Activities Pseudomonas spp. and Bacillus sp. to stimulate germination and seedling growth of tomato plants. Agric. Agric. Sci. Procedia 2016, 9, 419–423. [Google Scholar] [CrossRef] [Green Version]
- Anjugam, M.; Bharathidasan, R.; Shijila Rani, A.S.; Ambikapathy, V. Evaluation of antimicrobial activities of endophytic fungal metabolites against clinical importance microbes. J. Pharmacogn. Phytochem. 2019, 8, 1004–1007. [Google Scholar]
- Adeleke, B.S.; Babalola, O.O. Pharmacological Potential of Fungal Endophytes Associated with Medicinal Plants: A Review. J. Fungi 2021, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Al-Badri, B.A.; Al-Maawali, S.S.; AlBalushi, Z.M.; Al-Mahmooli, I.H.; Al-Sadi, A.M.; Velazhahan, R. Cyanide degradation and antagonistic potential of endophytic Bacillus subtilis strain BEB1 from Bougainvillea spectabilis Willd. All Life 2020, 13, 92–98. [Google Scholar] [CrossRef]
- Haruna, A.; Yahaya, S.M. Recent advances in the chemistry of bioactive compounds from plants and soil microbes: A Review. Chem. Africa 2021, 8, 231–345. [Google Scholar] [CrossRef]
- Balderas-Ruíz, K.A.; Bustos, P.; Santamaria, R.I.; González, V.; Cristiano-Fajardo, S.A.; Barrera-Ortíz, S.; Mezo-Villalobos, M.; Aranda-Ocampo, S.; Guevara-García, A.A.; Galindo, E.; et al. Bacillus velezensis 83 a bacterial strain from mango phyllosphere, useful for biological control and plant growth promotion. AMB Express 2020, 10, 163. [Google Scholar] [CrossRef]
- Riera, N.; Handique, U.; Zhang, Y.; Dewdney, M.M.; Wang, N. Characterization of Antimicrobial-Producing Beneficial Bacteria Isolated from Huanglongbing Escape Citrus Trees. Front. Microbiol. 2017, 8, 2415. [Google Scholar] [CrossRef]
- Ali, S.; Hameed, S.; Imran, A.; Iqbal, M.; Lazarovits, G. Genetic, physiological and biochemical characterization of Bacillus sp. strain RMB7 exhibiting plant growth promoting and broad-spectrum antifungal activities. Microb. Cell Factories 2014, 13, 144. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Rooney, A.P. Iturinic lipopeptide diversity in the Bacillus subtilis species group—Important antifungals for plant disease biocontrol applications. Front. Microbiol. 2019, 10, 1794. [Google Scholar] [CrossRef] [PubMed]
- Mülner, P.; Schwarz, E.; Dietel, K.; Junge, H.; Herfort, S.; Weydmann, M.; Lasch, P.; Cernava, T.; Berg, G.; Vater, J. Profiling for bioactive peptides and volatiles of plant growth promoting strains of the Bacillus subtilis complex of industrial relevance. Front. Microbiol. 2020, 11, 1432. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, D.; Qi, G.; Mao, Z.; Hu, X.; Du, B.; Liu, K.; Ding, Y. Effects of Bacillus velezensis FKM10 for Promoting the Growth of Malus hupehensis Rehd. and Inhibiting Fusarium verticillioides. Front. Microbiol. 2020, 10, 2889. [Google Scholar] [CrossRef]
- Kim, S.Y.; Song, H.; Sang, M.K.; Weon, H.Y.; Song, J. The complete genome sequence of Bacillus velezensis strain GH1-13 reveals agriculturally beneficial properties and a unique plasmid. J. Biotechnol. 2017, 259, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.R.; Mouillon, J.M.; Pohl, S.; Arnau, J. Secondary metabolite production and the safety of industrially important members of the Bacillus subtilis group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef]
- Gorai, P.S.; Ghosh, R.; Konra, S.; Mandal, N.C. Biological control of early blight disease of potato caused by Alternaria sp. EBP3 by an endophytic bacterial strain Bacillus velezensis SEB1. Biol. Control 2021, 156, 104551. [Google Scholar] [CrossRef]
- Dunlap, C.; Kim, S.J.; Kwon, S.W.; Rooney, A. Bacillus velezensis is not a later heterotypic synonym of Bacillus amyloliquefaciens, Bacillus methylotrophicus, Bacillus amyloliquefaciens subsp. plantarum and ‘Bacillus oryzicola’ are later heterotypic synonyms of of Bacillus velezensis based on phylogenomics. Int. J. Syst. Evol. Microbiol. 2016, 66, 1212–1217. [Google Scholar] [PubMed]
- Xu, T.; Zhu, T.; Li, S. β-1,3-1,4-glucanase gene from Bacillus velezensis ZJ20 exerts antifungal effect on plant pathogenic fungi. World J. Microbiol. Biotechnol. 2016, 32, 26. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.H. Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, Y.; Lai, X.H.; Shan, C.; Deng, Z.; Ji, Y. Screening and characterization of endophytic Bacillus and Paenibacillus strains from medicinal plant Lonicera japonica for use as potential plant growth promoters. Braz. J. Microbiol. 2015, 46, 977–989. [Google Scholar] [CrossRef]
- Chen, L.; Heng, J.; Qin, S.; Bian, K. A comprehensive understanding of the biocontrol potential of Bacillus velezensis LM2303 against Fusarium head blight. PLoS ONE 2018, 13, e0198560. [Google Scholar] [CrossRef]
- Moon, J.H.; Won, S.J.; Maung, C.E.H.; Choi, J.H.; Choi, S.I.; Ajuna, H.B.; Ahn, Y.S. Bacillus velezensis CE 100 inhibits root rot diseases (Phytophthora spp.) and promotes growth of Japanese cypress (Chamaecyparis obtuse Endlicher) seedlings. Microorganisms 2021, 9, 821. [Google Scholar] [CrossRef]
- Habib, M.R.; Karim, M.R. Antimicrobial and cytotoxic activity of di-(2-ethylhexyl) phthalate and anhydrosophoradiol-3-acetate isolated from Calotropis gigantea (Linn.) flower. Mycobiology 2009, 37, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; Mellouli, L.; Lebrihi, A.; Coppel, Y.; Fguira, L.F.B.; Mathieu, F. Purification and structure elucidation of three naturally bioactive molecules from the new terrestrial Streptomyces sp. TN17 strain. Nat. Prod. Res. 2011, 25, 806–814. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.H. Di-(2-ethylhexyl) phthalate, a major bioactive metabolite with antimicrobial and cytotoxic activity isolated from the culture filtrate of newly isolated soil streptomyces (Streptomyces mirabilis Strain NSQu-25). World Appl. Sci. J. 2012, 20, 1202–1212. [Google Scholar]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D.Don. Don. 3 Biotech 2016, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhu, X.; Zhou, S.; Cheng, Z.; Shi, K.; Zhang, C.; Shao, H. Phthalic acid esters: Natural sources and biological activities. Toxins 2021, 13, 495. [Google Scholar] [CrossRef]
- Al-Bari, M.A.A.; Sayeed, M.A.; Rahman, M.S.; Mossadik, M.A. Characterization and antimicrobial activities of a phthalic acid derivative produced by Streptomyces bangladeshiensis—A novel species in Bangladesh. Res. J. Med. Sci. 2006, 1, 77–81. [Google Scholar]
- Kanjana, M.; Kanimozhi, G.; Udayakumar, R.; Panneerselvam, A. GCMS analysis of bioactive compounds of endophytic fungi Chaetomium globosum, Cladosporium tenuissimum and Penicillium janthinellum. J. Biomed. Pharm. Sci. 2019, 2, 123. [Google Scholar]
- Roy, R.N.; Laskar, S.; Sen, S.K. Dibutyl phthalate, the bioactive compound produced by Streptomyces albidoflavus 321.2. Microbiol. Res. 2006, 161, 121–126. [Google Scholar] [CrossRef]
- Mohamad, O.; Ma, J.B.; Liu, Y.H.; Zhang, D.; Hua, S.; Bhute, S.; Hedlund, B.P.; Li, W.J.; Li, L. Beneficial endophytic bacterial populations associated with medicinal plant Thymus vulgaris alleviate salt stress and confer resistance to Fusarium oxysporum. Front. Plant Sci. 2020, 11, 47. [Google Scholar] [CrossRef]
- Birnie, C.; Malamud, D.; Schnaare, R. Antimicrobial evaluation of N-Alkyl Betaines and N-Alkyl-N,N-Dimethylamine oxides with variations in chain length. Antimicrob. Agents Chemother. 2000, 44, 2514–2517. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Correia, V.G.; Aguiar-Ricardo, A.; Cunha, Â.; Moutinho, M.G.M. Antimicrobial activity of new green-functionalized oxazoline-based oligomers against clinical isolates. Springer Plus 2015, 4, 382. [Google Scholar] [CrossRef] [PubMed]
- Shobier, A.H.S.; Abdel Ghani, S.A.; Barakat, K.M. GC/MS spectroscopic approach and antifungal potential of bioactive extracts produced by marine macroalgae. Egypt. J. Aquat. Res. 2016, 42, 289–299. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Senthilkumar, A.; Venkatesalu, V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 775–780. [Google Scholar] [PubMed]
- Mensah-Agyei, G.O.; Ayeni, K.I.; Ezeamagu, C.O. GC-MS analysis of bioactive compounds and evaluation of antimicrobial activity of the extracts of Daedalea elegans: A Nigerian mushroom. Afr. J. Microbiol. Res. 2020, 14, 204–210. [Google Scholar]
- Anwar, P.Z.; Sezhian, U.G.; Narasingam, A. Bioactive compound analysis and bioactivities of endophytic bacteria from Cissus quadrangularis. Int. J. Pharm. Sci. Res. 2020, 11, 5553–5560. [Google Scholar]
- Minerdi, D.; Bossi, S.; Gullino, M.L.; Garibaldi, A. Volatile organic compounds: A potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA 35. Environ. Microbiol. 2009, 11, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Tilocca, B.; Cao, A.; Migheli, Q. Scent of a killer: Microbial volatilome and its role in the biological control of plant pathogens. Front. Microbiol. 2020, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Raza, W.; Shen, Q.; Huang, Q. Antifungal activity of Bacillus amyloliquefaciens NJN6 volatile compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.H.; Liao, M.J.; Wang, H.K.; Zheng, M.Z.; Xu, J.J.; Guo, J.H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biol. Control 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Reda, F.M.; Shafi, S.A.; Ismail, M. Efficient inhibition of some multi-drug resistant pathogenic bacteria by bioactive metabolites from Bacillus amyloliquefaciens S5I4 isolated from archaeological soil in Egypt. Appl Biochem. Microbiol. 2016, 52, 593–601. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Asaf, S.; Lee, I. Plant growth-promoting endophytic bacteria versus pathogenic infections: An example of Bacillus amyloliquefaciens RWL-1 and Fusarium oxysporum f. sp. lycopersici in tomato. PeerJ 2017, 5, e3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Organ | Isolate No. | Antagonistic Activity | |
|---|---|---|---|---|
| Alternaria sp. | Helminthosporium sp. | |||
| Solanum melongena | Leaf, stem and Root | E1 | + | + |
| E2 | − | + | ||
| E3 | +++ | + | ||
| E4 | + | + | ||
| E5 | + | ++ | ||
| Allium cepa | Root and leaf | O1 | + | + |
| O2 | + | + | ||
| Portulaca oleracea | leaf | R1 | ++ | ++ |
| R2 | ++ | ++ | ||
| R3 | − | − | ||
| R4 | + | + | ||
| R5 | ++ | + | ||
| R6 | − | − | ||
| R7 | + | ++ | ||
| Coriandrum sativum | leaf | K1 | + | + |
| Pisum sativum | Leaf and stem | P1 | + | + |
| P2 | + | + | ||
| P3 | + | + | ||
| P4 | + | + | ||
| P5 | + | + | ||
| Solanum lycopersicum | leaf | T1 | + | + |
| T2 | + | + | ||
| T4 | + | ++ | ||
| T5 | +++ | +++ | ||
| Brassica oleracea | leaf | C1 | ++ | +++ |
| Isolate Number | Inhibition of Mycelial Growth (cm) | |||
|---|---|---|---|---|
| Alternaria sp. | Helminthosporium sp. | |||
| Bacterial Filtrate (Cell-Free Culture) (150 μL) | Bacterial Disc (15 mm) | Bacterial Filtrate (Cell-Free Culture) (150 μL) | Bacterial Disc (15 mm) | |
| E3 | 3.8 ± 0.201 a | 2.6 ± 0.137 c | 2.5 ± 0.132 a | 2.5 ± 0.132 cd |
| E5 | 3 ± 0.159 b | 2.7 ± 0.143 bc | 2.7 ± 0.143 a | 2.37 ± 0.125 cd |
| R1 | 3.6 ± 0.191 a | 3.1 ± 0.164 b | 2.7 ± 0.143 a | 2.25 ± 0.119 d |
| R2 | 4 ± 0.212 a | 3 ± 0.158 bc | 2.5 ± 0.132 a | 2.8 ± 0.148 c |
| T4 | 3.8 ± 0.201 a | 2.1 ± 0.111 d | 1.8 ± 0.095 b | 0.9 ± 0.064 e |
| T5 | 3.6 ± 0.191 a | 4.0 ± 0.212 a | 2.6 ± 0.138 a | 3.9 ± 0.329 b |
| C1 | 4.2 ± 0.222 a | 3.1 ± 0.164 b | 2.8 ± 0.148 a | 4.77 ± 0.252 a |
| Characteristics | B. velezensis | B. amyloliquefaciens |
|---|---|---|
| Shape | Rod | Rod |
| Gram stain | + | + |
| Spore formation | + | + |
| Oxidase | + | + |
| Indole test | - | - |
| Hydrogen sulphide | - | - |
| Catalase reaction | + | + |
| Methyl red test | - | - |
| Nitrate (reduction) | + | + |
| Voges-proskauer test | + | + |
| Urease | - | - |
| Hydrolyzed Starch | + | + |
| Gelatin liquefaction | + | + |
| Growth in 10% NaCl | - | + |
| No. | Compound Name and Class | Molecular Formula | MW | Area% | RT (min) | Base Peak (100%) |
|---|---|---|---|---|---|---|
| 1 | 6,6-Dimethyl-1,3-heptadien-5-ol (Alchols) | C9H16O | 140 | 0.46 | 2.44 | 57.0 |
| 2 | Benzaldehyde, 3-benzyloxy-2-fluoro-4-methoxy benzaldehyde (Aldehyde) | C15H13FO3 | 260 | 2.91 | 3.69 | 91.0 |
| 3 | Chloromethyl benzene (Halobenzene) | C7H7Cl | 126 | 0.73 | 3.80 | 91.0 |
| 4 | decyloxy anime (Amines) | C10H23NO | 173 | 0.98 | 5.75 | 43.0 |
| 5 | Naphthalene, 1,2,3,4-tetrahydro-5-methyl-(poly nuclear aromatic cpds) | C11H14 | 146 | 0.61 | 7.22 | 131.0 |
| 6 | 1,3,5-Triazine-2,4-diamine, 6-chloro-N-ethyl- (Heterocyclic cpds) | C5H8ClN5 | 173 | 0.34 | 7.84 | 43.0 |
| 7 | N,N-Dimethyldodecylamine (Tertiary amine) | CH3(CH2)11N(CH3)2 | 213 | 11.84 | 13.34 | 58.0 |
| 8 | 5-Octadecene (Alkene) | C18H36 | 252 | 0.21 | 15.65 | 55.0 |
| 9 | Cetene (Alkene) | C16H32 | 224 | 0.21 | 15.65 | 41.0 |
| 10 | Diethyl phthalate (Esters) | C12H14O4 | 222 | 2.05 | 15.79 | 149.0 |
| 11 | N,N-Dimethyltetradecylamine (Tertiary amine) | C16H35N | 241 | 6.57 | 18.74 | 58.0 |
| 12 | 1-Docosene (Alkene) | C22H44 | 308 | 1.38 | 21.02 | 55.0 |
| 13 | 9-Nonadecene (Alkene) | C19H38 | 266 | 1.38 | 21.02 | 41.0 |
| 14 | 9-Eicosene, (E)- (Alkene) | C20H40 | 280 | 1.38 | 21.02 | 57.0 |
| 15 | Octadecane (Alkane) | CH3(CH2)16CH3 | 254 | 0.19 | 21.19 | 57.0 |
| 16 | 4-Phenyleicosane (Alkyl benzene) | C26H46 | 358 | 0.28 | 22.36 | 91.0 |
| 17 | Methyl palmitate (fatty ester) | C17H34O2 | 270 | 2.11 | 24.40 | 74.0 |
| 18 | Dibutyl phthalate (Esters) | C16H22O4 | 278 | 8.10 | 25.35 | 149.0 |
| 19 | Ethyl hexadecanoate (fatty esters) | C18H36O2 | 284 | 3.25 | 26.06 | 88.0 |
| 20 | Methyl linoleate (un-saturated fatty ester) | C19H34O2 | 294 | 1.47 | 28.35 | 67.0 |
| 21 | Methyl 11-Octadecenoate (unsaturated fatty ester) | C19H36O2 | 296 | 2.86 | 28.51 | 55.0 |
| 22 | 3-(N-Benzyl-N-methylamino)-1,2-propanediol (Amino alchol) | C11H17NO2 | 195 | 4.09 | 28.73 | 91.0 |
| 23 | Methyl stearate (Sat. fatty acids) | C19H38O2 | 298 | 0.94 | 29.12 | 74.0 |
| 24 | 1,3,5(10)-Oestratrien-17α-ol (Chlosterol) | C18H24O | 256 | 0.36 | 29.52 | 43.0 |
| 25 | Ethyl oleate (un-sat. fatty acids) | C20H38O2 | 310 | 0.74 | 30.01 | 55.0 |
| 26 | Ethyl Octadecanoate (Sat.fatty esters) | C20H40O2 | 312 | 1.91 | 30.62 | 88.0 |
| 27 | N-Methyl-N-benzyltetradecanamine (Tertiary amine) | C22H39N | 317 | 1.50 | 33.24 | 134.0 |
| 28 | 1-Phenylacetone (ketone) | C9H10O2 | 134 | 1.50 | 33.24 | 43.00 |
| 29 | Bis (2-ethylhexyl) ester (Esters) | C22H42O4 | 370 | 18.59 | 35.06 | 129.0 |
| 30 | Octyl hexadecanoate (Sat. fatty acids) | C24H48O2 | 368 | 2.33 | 36.99 | 257.0 |
| 31 | Bis(2-ethylhexyl) phthalate (Esters) | C24H38O4 | 390 | 20.36 | 38.00 | 149.0 |
| No. | Compound Name and Class | Molecular Formula | MW | Area % | RT (min) | Base Peak (100%) |
|---|---|---|---|---|---|---|
| 1 | 4-isopropenyl-1-methylcyclohexene (R(+) Limonene) | C10H16 | 136 | 1.42 | 2.84 | 68.0 |
| 2 | Perilla alcohol (Alkaloids) | C10H16O | 152 | 0.40 | 3.17 | 41.0 |
| 3 | Benzaldehyde, 3-benzyloxy-2-fluoro-4-methoxy- (Aldehyde) | C15H13FO3 | 260 | 0.59 | 3.53 | 91.0 |
| 4 | Chloromethyl benzene (Halobenzene) | C7H7Cl | 126 | 0.32 | 3.81 | 91.0 |
| 5 | Dodec-1-ene (Alkane) | C12H24 | 168 | 0.25 | 5.38 | 43.0 |
| 6 | 5-Isopropenyl-2-methyl-2-cyclohexen-1-one (alkaloids) | C10H16O | 150 | 0.36 | 6.86 | 82.0 |
| 7 | 1,3,5-Triazine-2,4-diamine, 6-chloro-N-ethyl-(Heterocyclic cpds) | C5H8ClN5 | 173 | 0.70 | 7.29 | 43.0 |
| 8 | 4,4,6-Trimethyl-6-phenyl-1,3-oxazinane-2-thione (Heterocycliccpds) | C13H17NOS | 235 | 0.34 | 7.29 | 118.0 |
| 9 | N,N-Dimethyldodecylamine (Tertiary amine) | CH3(CH2)11N(CH3)2 | 213 | 9.08 | 13.34 | 58.0 |
| 10 | Methyl 10-methylundecanoate (saturated Fatty ester) | C13H26O2 | 214 | 0.17 | 13.83 | 74.0 |
| 11 | Diethyl phthalate (Esters) | C12H14O4 | 222 | 0.17 | 15.77 | 149.0 |
| 12 | Methyl tetradecanoate (Sat. fatty acids) | C15H30O2 | 242 | 0.29 | 19.27 | 74.0 |
| 13 | N,N-Dimethyltetradecylamine (Tertiary amine) | C16H35N | 241 | 5.21 | 18.73 | 58.0 |
| 14 | Methyl 12-methyltetradecanoate (Sat. fatty acids) | C16H32O2 | 256 | 1.10 | 20.92 | 74.0 |
| 15 | 9-Eicosene, (E)-(Alkene) | C20H40 | 280 | 1.06 | 21.02 | 57.0 |
| 16 | Methyl 9-oxodecanoate (Esters) | C11H20O3 | 200 | 0.87 | 21.93 | 43.0 |
| 17 | Methyl 14-methylpentadecanoate (Sat. fatty esters) | C17H34O2 | 270 | 3.88 | 23.48 | 74.0 |
| 18 | Dibutyl phthalate (Esters) | C16H22O4 | 278 | 8.18 | 25.37 | 149.0 |
| 19 | Ethyl hexadecanoate (Fatty esters) | C18H36O2 | 284 | 3.24 | 26.06 | 88.0 |
| 20 | Methyl linoleate (un-saturated fatty ester) | C19H34O2 | 294 | 2.85 | 28.36 | 67.0 |
| 21 | Methyl elaidate (un-saturated fatty ester) | C19H36O2 | 296 | 4.03 | 28.53 | 55.0 |
| 22 | Methyl stearate (Sat. fatty acids) | C19H38O2 | 298 | 1.87 | 29.13 | 74.0 |
| 23 | Ethyl 9-octadecenoate (unsat. fatty ester) | C20H38O2 | 310 | 0.68 | 30.01 | 55.0 |
| 24 | Ethyl Octadecenoate (Sat. fatty ester) | C20H20O2 | 312 | 1.88 | 30.62 | 88.0 |
| 25 | N-Methyl-N-benzyltetradecanamine (Tertiary amine) | C22H39N | 317 | 1.30 | 33.24 | 134.0 |
| 26 | 1-Phenyl acetone (ketone) | C9H10O | 134 | 1.30 | 33.24 | 43.0 |
| 27 | Bis (2-ethylhexyl) ester (Esters) | C22H42O4 | 370 | 14.98 | 35.04 | 129.0 |
| 28 | Octyl hexadecanoate (Sat. fatty acids) | C24H48O2 | 368 | 2.15 | 36.98 | 257.0 |
| 29 | Bis(2-ethylhexyl) phthalate (Esters) | C24H38O4 | 390 | 24.39 | 38.04 | 149.0 |
| 30 | Octadecanoic acid (Sat. fatty acids) | C18H36O2 | 284 | 0.36 | 40.75 | 43.0 |
| 31 | Dinonyl phthalate (Esters) | C26H42O4 | 418 | 0.17 | 41.14 | 149.0 |
| Treatments | Plant Height (cm/Plant) | TFW (g/Plant) | TDW (g/Plant) | Disease Incidence DI (%) |
|---|---|---|---|---|
| Control | 26 ± 0.687 ab | 7.73 ± 0.205 b | 1.339 ± 0.035 a | 0 ± 0.0 c |
| Alternaria sp. | 20.8 ± 0.55 c | 3.81 ± 0.101 d | 0.668 ± 0.018 c | 80 ± 2.07 a |
| B. amyloliquefaciens | 28 ± 0.74 a | 8.36 ± 0.221 a | 1.4299 ± 0.038 a | 0 ± 0.0 c |
| Alternaria sp. + B. amyloliquefaciens | 24.5 ± 0.648 b | 5.88 ± 0.156 c | 0.8024 ± 0.021 b | 40 ± 1.096 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, S.A.; Khaleil, M.M.; Metwally, R.A. Evaluation of the Antifungal Activity of Bacillus amyloliquefaciens and B. velezensis and Characterization of the Bioactive Secondary Metabolites Produced against Plant Pathogenic Fungi. Biology 2022, 11, 1390. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101390
Soliman SA, Khaleil MM, Metwally RA. Evaluation of the Antifungal Activity of Bacillus amyloliquefaciens and B. velezensis and Characterization of the Bioactive Secondary Metabolites Produced against Plant Pathogenic Fungi. Biology. 2022; 11(10):1390. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101390
Chicago/Turabian StyleSoliman, Shereen A., Mona M. Khaleil, and Rabab A. Metwally. 2022. "Evaluation of the Antifungal Activity of Bacillus amyloliquefaciens and B. velezensis and Characterization of the Bioactive Secondary Metabolites Produced against Plant Pathogenic Fungi" Biology 11, no. 10: 1390. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11101390