
Pond Assay for the Sensory Systems of Caenorhabditis elegans: A Novel Anesthesia-Free Method Enabling Detection of Responses to Extremely Low Chemical Concentrations


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture
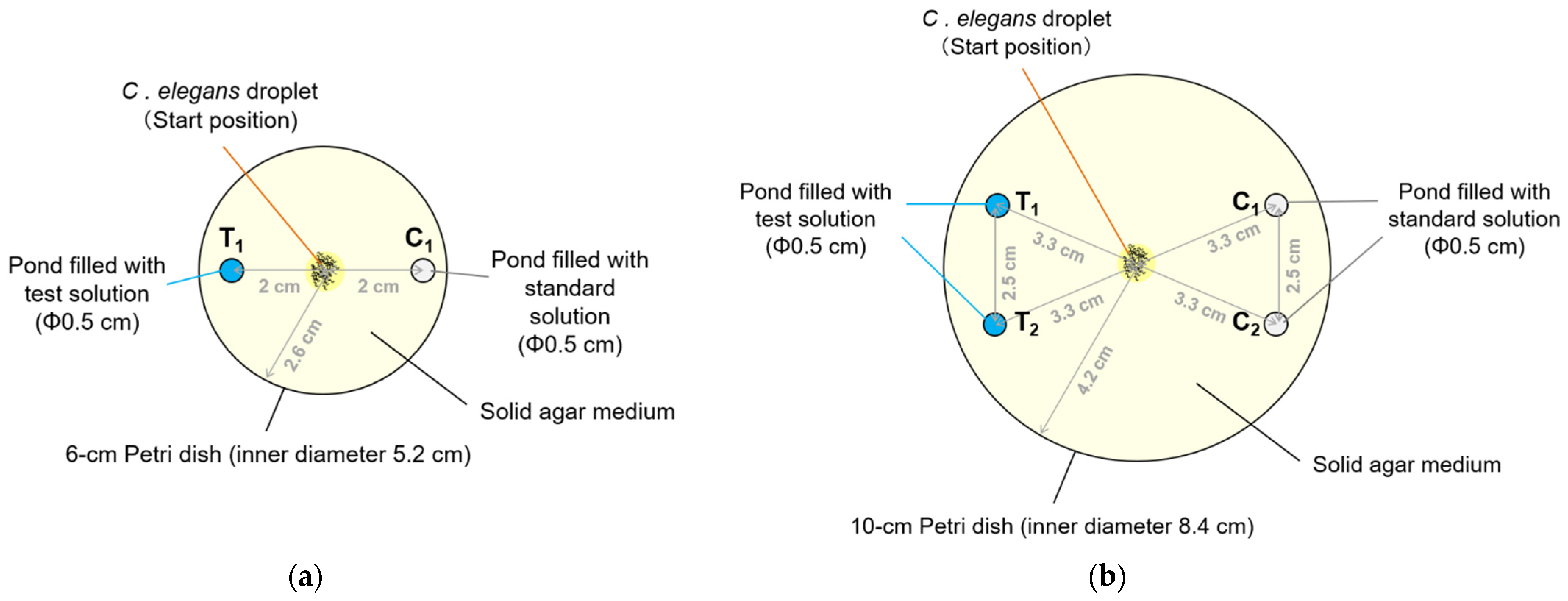
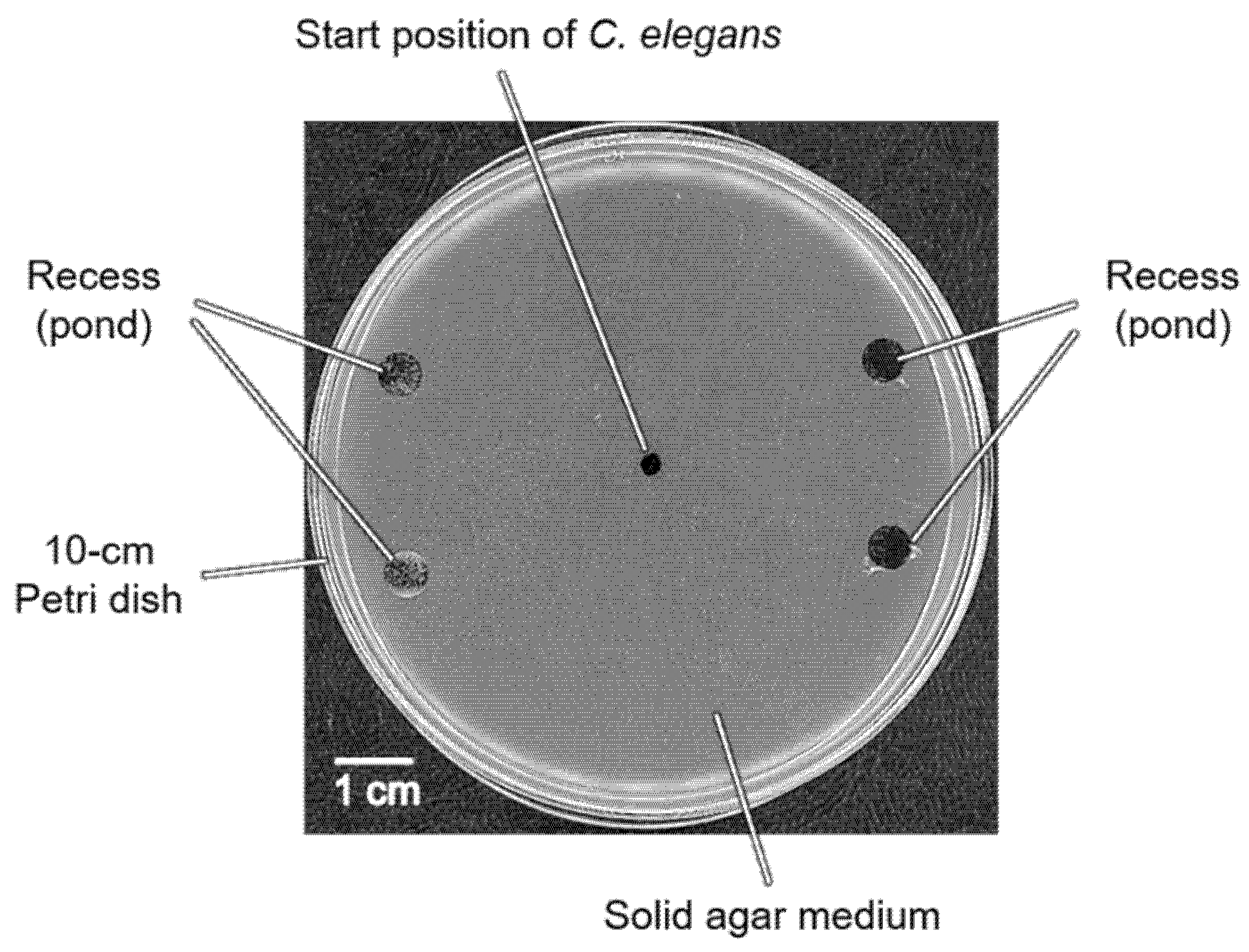
2.2. Basics of Preparation of a Novel Plate for the Pond Assay for the Sensory Systems (PASS Plate)
2.3. Preparation of C. elegans Individuals for the PASS
2.4. Counting the Number of C. elegans Individuals and Its Support Software
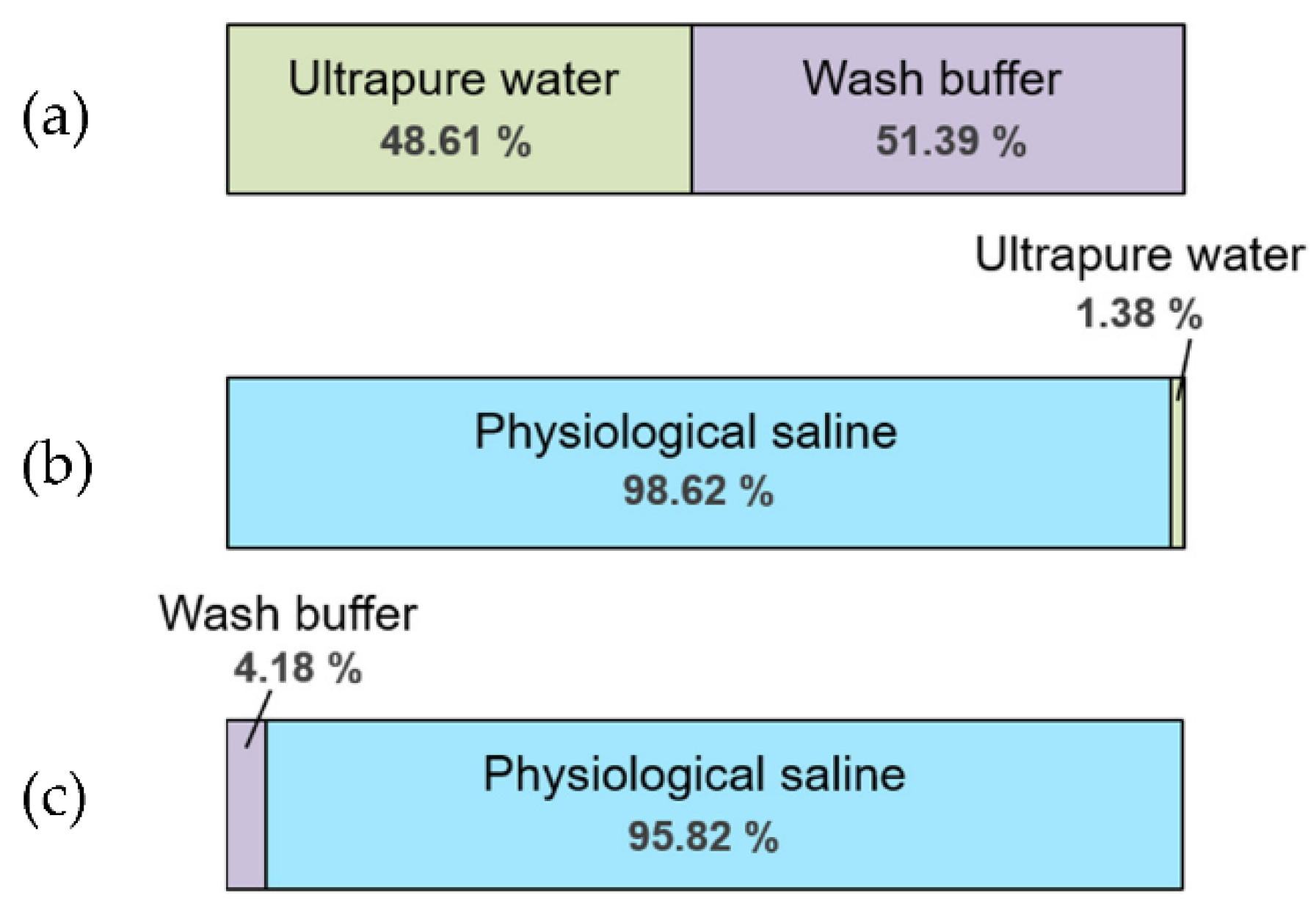
2.5. Selection of Liquid Appropriate for a Standard Solution of the PASS
2.6. PASS for Detection of Olfactory Response of C. elegans to Diacetyl
2.6.1. PASS
2.6.2. Conventional Assay
2.7. PASS for Detection of Taste Response of C. elegans to NaCl
2.7.1. PASS
2.7.2. Conventional Assay
2.8. Statistical Analysis
3. Results
3.1. Development of Pond Assay for the Sensory Systems of C. elegans
3.2. Selection of Liquid Appropriate for a Standard Solution Filling the Ponds of the PASS
3.3. Detection of Response of C. elegans to Diacetyl Based on the PASS
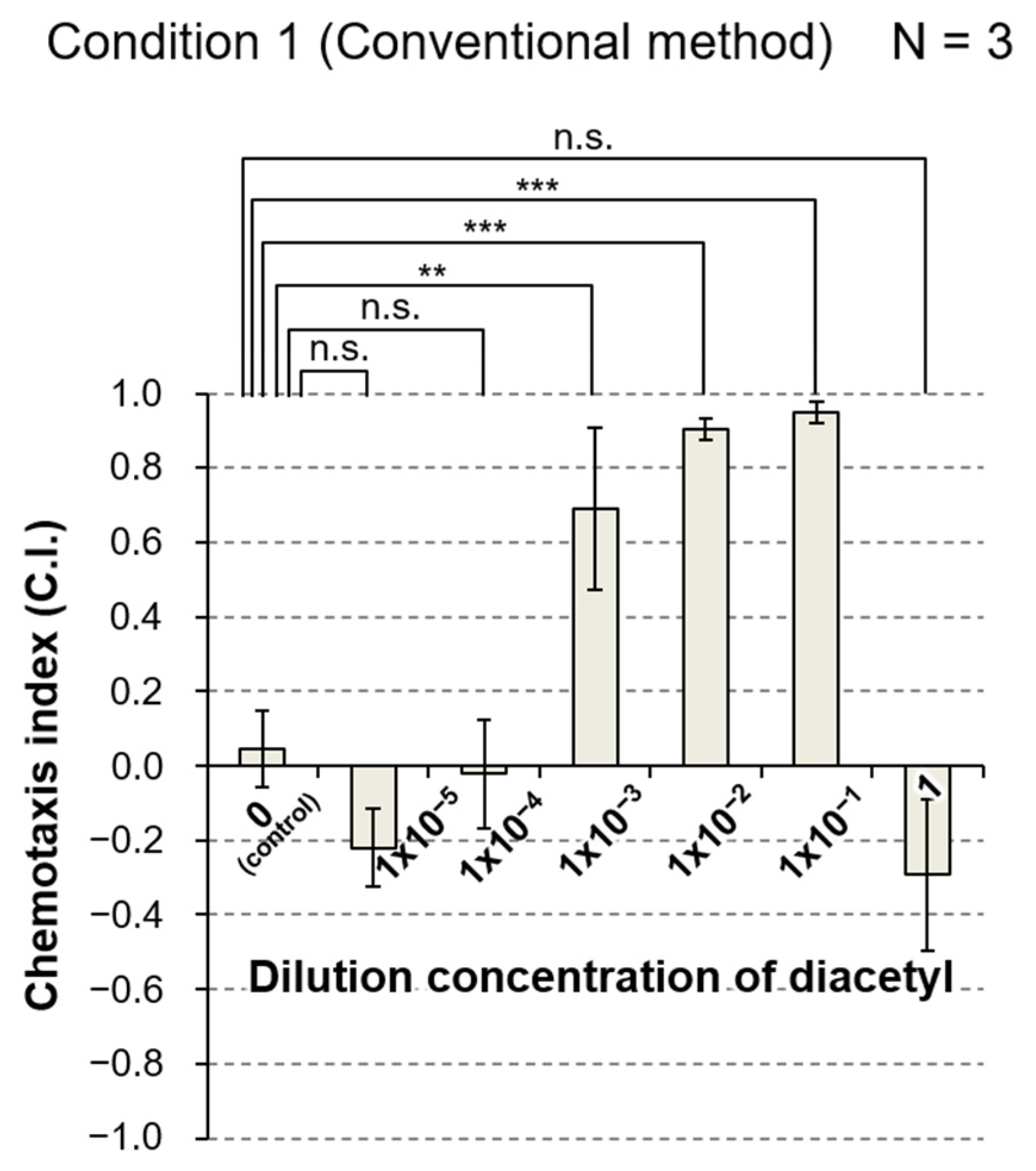
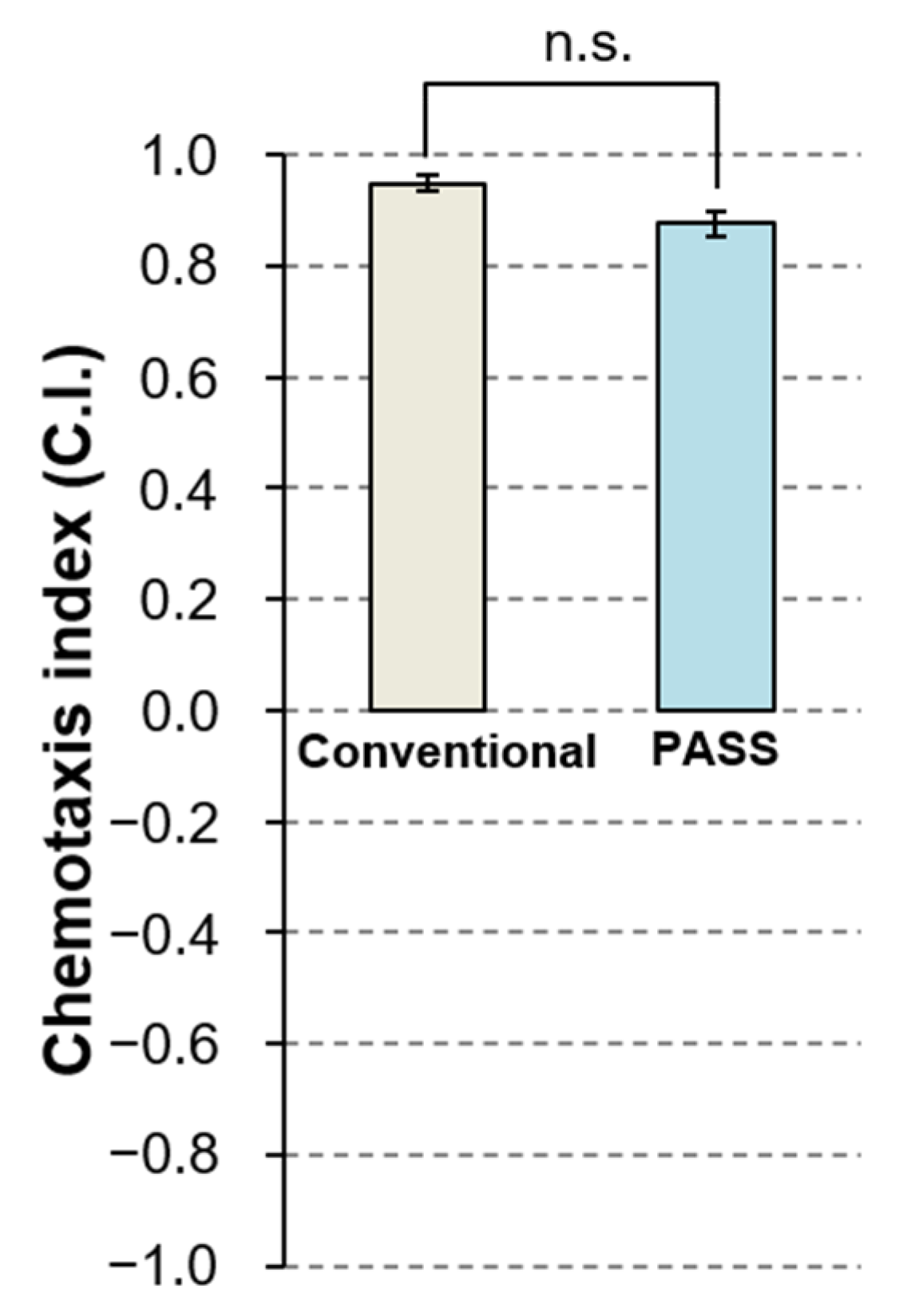
3.3.1. Comparison of the Conventional Method and the Novel PASS Method
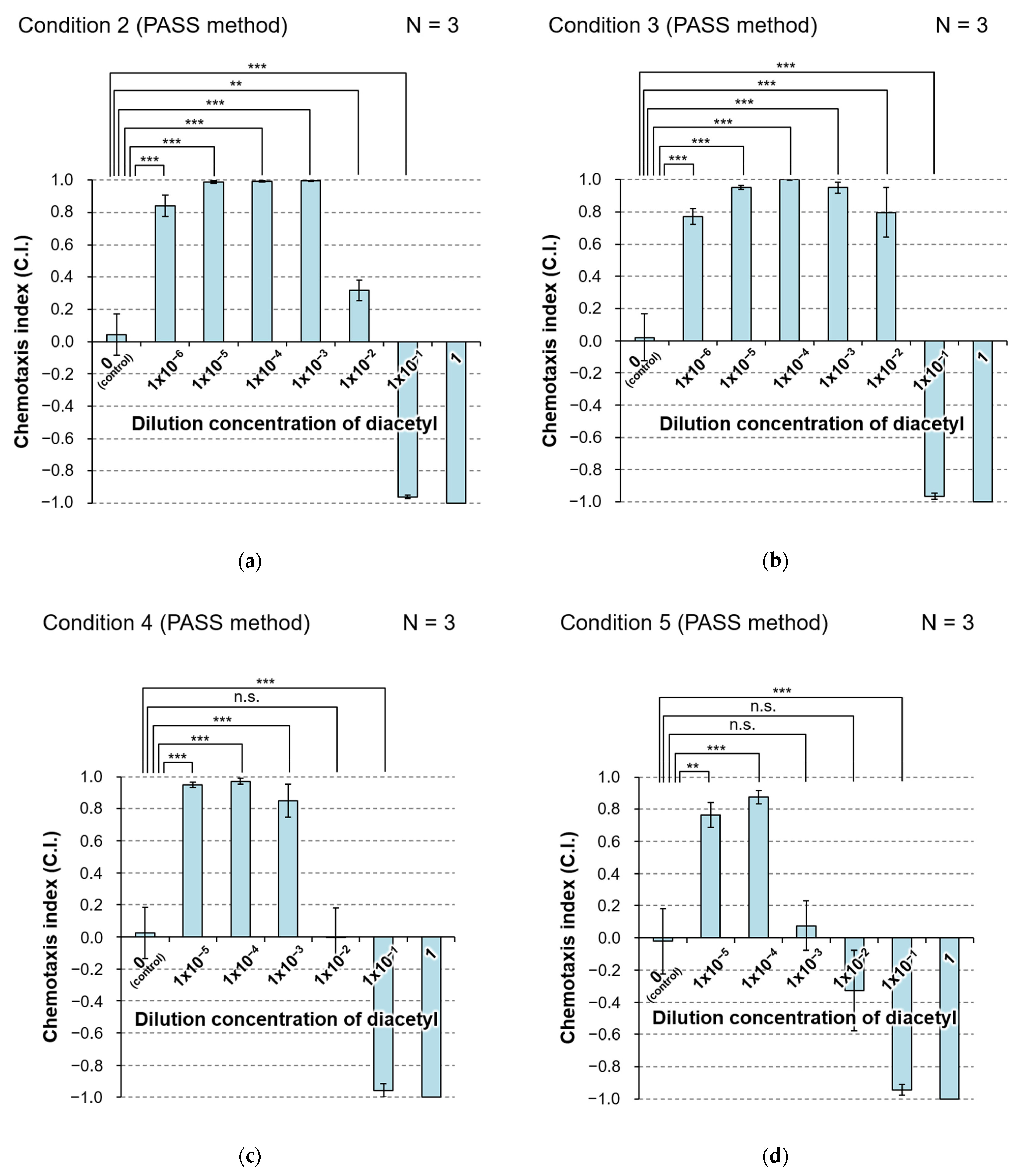
3.3.2. Effect of Volume of Agar Medium on Performance of PASS
3.3.3. Effect of Type of Solvent on Performance of the PASS
3.3.4. Effect of Size of the Plate on Performance of the PASS
3.3.5. Relationship between the Number of Assay Iterations and Reliability of the PASS Results
3.4. Detection of Chemotaxis of C. elegans to a Concentration Gradient of NaCl
4. Discussion
4.1. PASS Method without Anesthesia Improves Experimental Efficiency and Accuracy
4.2. Advance of Assay Method Leads to a Correct Understanding of the Sensory Ability of Living Organisms
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- White, J.G.; Southgate, E.; Thomson, J.N.; Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. B Biol. Sci. 1986, 314, 1–340. [Google Scholar] [CrossRef]
- Ward, S. Chemotaxis by the Nematode Caenorhabditis elegans: Identification of Attractants and Analysis of the Response by Use of Mutants. Proc. Natl. Acad. Sci. USA 1973, 70, 817–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusenbery, D.B. Analysis of chemotaxis in the nematode Caenorhabditis elegans by countercurrent separation. J. Exp. Zool. 1974, 188, 41–47. [Google Scholar] [CrossRef]
- Hedgecock, E.M.; Russell, R.L. Normal and mutant thermotaxis in the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1975, 72, 4061–4065. [Google Scholar] [CrossRef] [Green Version]
- Bargmann, C.I.; Horvitz, H.R. Chemosensory neurons with overlapping functions direct chemotaxis to multiple chemicals in C. elegans. Neuron 1991, 7, 729–742. [Google Scholar] [CrossRef]
- Bargmann, C.I.; Hartwieg, E.; Horvitz, H.R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 1993, 74, 515–527. [Google Scholar] [CrossRef]
- Sengupta, P.; Chou, J.; Bargmann, C.I. Odr-10 Encodes a Seven Transmembrane Domain Olfactory Receptor Required for Responses to the Odorant Diacetyl. Cell 1996, 84, 899–909. [Google Scholar] [CrossRef] [Green Version]
- Colbert, H.A.; Bargmann, C.I. Environmental signals modulate olfactory acuity, discrimination, and memory in Caenorhabditis elegans. Learn. Mem. 1997, 4, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Troemel, E.R.; Kimmel, B.E.; Bargmann, C.I. Reprogramming Chemotaxis Responses: Sensory Neurons Define Olfactory Preferences in C. elegans. Cell 1997, 91, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Mori, I.; Ohshima, Y. Molecular neurogenetics of chemotaxis and thermotaxis in the nematode Caenorhabditis elegans. BioEssays 1997, 19, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Mori, I. Genetics of Chemotaxis and Thermotaxis in the Nematode Caenorhabditis elegans. Annu. Rev. Genet. 1999, 33, 399–422. [Google Scholar] [CrossRef] [PubMed]
- Hirotsu, T.; Saeki, S.; Yamamoto, M.; Iino, Y. The Ras-MAPK pathway is important for olfaction in Caenorhabditis elegans. Nature 2000, 404, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, H.S.; Ferkey, D.M.; Apicella, A.J.; Lans, H.; Sharmeen, T.; Chen, W.; Lefkowitz, R.J.; Jansen, G.; Schafer, W.R.; Hart, A.C. G Protein-Coupled Receptor Kinase Function Is Essential for Chemosensation in C. elegans. Neuron 2004, 42, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.C.; Thiele, T.R.; Faumont, S.; Moravec, M.L.; Lockery, S.R. Step-Response Analysis of Chemotaxis in Caenorhabditis elegans. J. Neurosci. 2005, 25, 3369–3378. [Google Scholar] [CrossRef] [Green Version]
- Palmitessa, A.; Hess, H.A.; Bany, I.A.; Kim, Y.-M.; Koelle, M.R.; Benovic, J.L. Caenorhabditus elegans Arrestin Regulates Neural G Protein Signaling and Olfactory Adaptation and Recovery. J. Biol. Chem. 2005, 280, 24649–24662. [Google Scholar] [CrossRef] [Green Version]
- Olsen, D.P.; Phu, D.; Libby, L.J.M.; Cormier, J.A.; Montez, K.M.; Ryder, E.F.; Politz, S.M. Chemosensory control of surface antigen switching in the nematode Caenorhabditis elegans. Genes Brain Behav. 2007, 6, 240–252. [Google Scholar] [CrossRef]
- Sakashita, T.; Hamada, N.; Suzuki, M.; Ikeda, D.D.; Yanase, S.; Ishii, N.; Kobayashi, Y. Effects of γ-ray irradiation on olfactory adaptation to benzaldehyde in Caenorhabditis elegans. Biol. Sci. Space 2007, 21, 117–120. [Google Scholar] [CrossRef]
- Matsuura, T.; Endo, S.; Iwamoto, R.; Takahashi, H.; Ichinose, M. Developmental changes in chemotactic response and choice of two attractants, sodium acetate and diacetyl, in the nematode Caenorhabditis elegans. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 920–927. [Google Scholar] [CrossRef]
- Luo, L.; Gabel, C.V.; Ha, H.-I.; Zhang, Y.; Samuel, A.D.T. Olfactory Behavior of Swimming C. elegans Analyzed by Measuring Motile Responses to Temporal Variations of Odorants. J. Neurophysiol. 2008, 99, 2617–2625. [Google Scholar] [CrossRef] [Green Version]
- Tsunozaki, M.; Chalasani, S.H.; Bargmann, C.I. A Behavioral Switch: cGMP and PKC Signaling in Olfactory Neurons Reverses Odor Preference in C. elegans. Neuron 2008, 59, 959–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Jee, C.; McIntire, S.L. Ethanol preference in C. elegans. Genes Brain Behav. 2009, 8, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Ferkey, D.M.; Sengupta, P.; L’Etoile, N.D. Chemosensory signal transduction in Caenorhabditis elegans. Genetics 2021, 217, iyab004. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hirotsu, T.; Tagawa, T.; Oda, S.; Wakabayashi, T.; Iino, Y.; Ishihara, T. Odour concentration-dependent olfactory preference change in C. elegans. Nat. Commun. 2012, 3, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, G.; Uozumi, T.; Kiriyama, K.; Kamizaki, T.; Hirotsu, T. Screening of Odor-Receptor Pairs in Caenorhabditis elegans Reveals Different Receptors for High and Low Odor Concentrations. Sci. Signal. 2014, 7, ra39. [Google Scholar] [CrossRef]
- Tangrodchanapong, T.; Sobhon, P.; Meemon, K. Frondoside A Attenuates Amyloid-β Proteotoxicity in Transgenic Caenorhabditis elegans by Suppressing Its Formation. Front. Pharmacol. 2020, 11, 553579. [Google Scholar] [CrossRef]
- Saeki, S.; Yamamoto, M.; Iino, Y. Plasticity of chemotaxis revealed by paired presentation of a chemoattractant and starvation in the nematode Caenorhabditis elegans. J. Exp. Biol. 2001, 204, 1757–1764. [Google Scholar] [CrossRef]
- Tomioka, M.; Adachi, T.; Suzuki, H.; Kunitomo, H.; Schafer, W.; Iino, Y. The Insulin/PI 3-Kinase Pathway Regulates Salt Chemotaxis Learning in Caenorhabditis elegans. Neuron 2006, 51, 613–625. [Google Scholar] [CrossRef] [Green Version]
- Torayama, I.; Ishihara, T.; Katsura, I. Caenorhabditis elegans Integrates the Signals of Butanone and Food to Enhance Chemotaxis to Butanone. J. Neurosci. 2007, 27, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Sakashita, T.; Hamada, N.; Ikeda, D.D.; Yanase, S.; Suzuki, M.; Ishii, N.; Kobayashi, Y. Modulatory effect of ionizing radiation on food-NaCl associative learning: The role of γ subunit of G protein in Caenorhabditis elegans. FASEB J. 2008, 22, 713–720. [Google Scholar] [CrossRef]
- Suzuki, M.; Sakashita, T.; Hattori, Y.; Yokota, Y.; Kobayashi, Y.; Funayama, T. Development of ultra-thin chips for immobilization of Caenorhabditis elegans in microfluidic channels during irradiation and selection of buffer solution to prevent dehydration. J. Neurosci. Methods 2018, 306, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.G.; Oyama, K.; Taguchi, M. A simple method for production of hydrophilic, rigid, and sterilized multi-layer 3D integrated polydimethylsiloxane microfluidic chips. Lab Chip 2020, 20, 2354–2363. [Google Scholar] [CrossRef] [PubMed]
- Hirotsu, T.; Sonoda, H.; Uozumi, T.; Shinden, Y.; Mimori, K.; Maehara, Y.; Ueda, N.; Hamakawa, M. A Highly Accurate Inclusive Cancer Screening Test Using Caenorhabditis elegans Scent Detection. PLoS ONE 2015, 10, e0118699. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.; Feria, N.S.; Yoshioka, A.; Tu, E.; Civitci, F.; Estes, S.; Wagner, J.T. A Caenorhabditis elegans behavioral assay distinguishes early stage prostate cancer patient urine from controls. Biol. Open 2021, 10, bio057398. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Konno, M.; Ozaki, M.; Kawamoto, K.; Chijimatsu, R.; Kondo, N.; Hirotsu, T.; Ishii, H. Scent test using Caenorhabditis elegans to screen for early-stage pancreatic cancer. Oncotarget 2021, 12, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Lanza, E.; Di Rocco, M.; Schwartz, S.; Caprini, D.; Milanetti, E.; Ferrarese, G.; Lonardo, M.T.; Pannone, L.; Ruocco, G.; Martinelli, S.; et al. C. elegans-based chemosensation strategy for the early detection of cancer metabolites in urine samples. Sci. Rep. 2021, 11, 17133. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Method | Plate Size | Volume of Agar | Number of Ponds | Solvent | Control Solution | Volume of Liquid | Figure | |
|---|---|---|---|---|---|---|---|---|---|
| Test | Control | ||||||||
| 1 | Conventional | ⌀10 cm | 10 mL | - (2 spots) | - (2 spots) | Wash buffer | Wash buffer | 1 µL | 5, S3a |
| 2 | PASS | ⌀10 cm | 10 mL | 2 | 2 | Wash buffer | Wash buffer | 35 µL | 6a, S3b |
| 3 | PASS | ⌀10 cm | 20 mL | 2 | 2 | Wash buffer | Wash buffer | 80 µL | 6b, S2a, S3c |
| 4 | PASS | ⌀10 cm | 10 mL | 2 | 2 | Ultrapure water | Ultrapure water | 35 µL | 6c, S2b, S3d |
| 5 | PASS | ⌀6 cm | 3 mL | 1 | 1 | Ultrapure water | Ultrapure water | 25 µL | 6d, S3e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, M.; Hattori, Y.; Saito, T.; Harada, Y. Pond Assay for the Sensory Systems of Caenorhabditis elegans: A Novel Anesthesia-Free Method Enabling Detection of Responses to Extremely Low Chemical Concentrations. Biology 2022, 11, 335. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020335
Suzuki M, Hattori Y, Saito T, Harada Y. Pond Assay for the Sensory Systems of Caenorhabditis elegans: A Novel Anesthesia-Free Method Enabling Detection of Responses to Extremely Low Chemical Concentrations. Biology. 2022; 11(2):335. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020335
Chicago/Turabian StyleSuzuki, Michiyo, Yuya Hattori, Toshiyuki Saito, and Yoshinobu Harada. 2022. "Pond Assay for the Sensory Systems of Caenorhabditis elegans: A Novel Anesthesia-Free Method Enabling Detection of Responses to Extremely Low Chemical Concentrations" Biology 11, no. 2: 335. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020335