
Ceratocystiopsis quercina sp. nov. Associated with Platypus cylindrus on Declining Quercus suber in Portugal

 ,
,  and
and
Abstract
:Simple Summary
Abstract
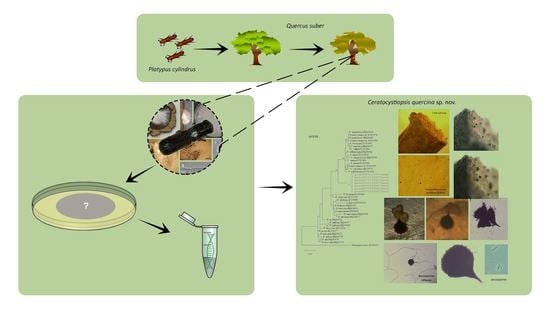
1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
2.2. Microscopic Observation and Descriptions
3. Results
3.1. Fungal Isolation and Identification
3.2. Phylogenetic Analyses
3.3. Morphology and Taxonomy
- (1)
- MycoBank: MB 841995;
- (2)
- Holotype: LISE 96335;
- (3)
- Etymology: Named after the host genus from which it was isolated, Quercus;
- (4)
- Host trees/distribution: On galleries of Quercus suber in Portugal = on mycangia of Platypus cylindrus.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seabra, A.F. Contribuição para a história da Entomologia em portugal. Publicações Dir. Geral Serv. Florestais Aquícolas 1939, 6, 1–20. [Google Scholar]
- Baeta-Neves, C.M. Lista dos insectos da biocenose do sobreiro (Quercus suber L.). Bol. Soc. Port. Ciên. Nat. 1950, 111, 33–46. [Google Scholar]
- Español, F. Los Platipodidos de Cataluña (Col. Phytophagoidea). Bol. Ser. Plagas 1964, 7, 115–117. [Google Scholar]
- Ferreira, M.C.; Ferreira, G.W. Platypus cylindrus F. (Coleóptera: Platipodidae) Plaga de Quercus suber. Bol. San. Veg. Plagas 1989, 4, 301–305. [Google Scholar]
- Sousa, E.M. Contribuition à l’étude de la biologie de populations de Platypus cylindrus (Coleoptera: Platypodidae) dans des peuplements de chênes liège au Portugal. In Thèse Pour L’Obtention du Diplôme de Doctorat; Université de Lyon: Lyon, France, 1996; p. 153. [Google Scholar]
- Sousa, E.M.; Débouzie, D. Caractéristiques bioécologiques de Platypus cylindrus au Portugal. IOBC/Wprs Bull. 2002, 25, 75–83. [Google Scholar]
- Batra, L.R. Ecology of ambrosia fungi and their dissemination by beetles. Trans. Kans. Acad. Sci. 1963, 66, 213–236. [Google Scholar] [CrossRef]
- Beaver, R.A. Insect-fungus relationships in the bark and ambrosia beetles. In Insect-Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 121–143. [Google Scholar]
- Francke-Grosmann, H. Ectosymbiosis in wood-inhabiting insects. In Symbiosis; Henry, S.M., Ed.; Academic Press: New York, NY, USA, 1967; Volume 11, pp. 142–206. [Google Scholar]
- Fraedrich, S.W.; Harrington, T.C.; Rabaglia, R.J.; Ulyshen, M.D.; Mayfield, A.E.; Hanula, J.L.; Eickwort, J.M.; Miller, D.R. A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other Lauraceae in the southeastern United States. Plant. Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Inácio, M.L.; Henriques, J.; Sousa, E. Mycobiota associated with Platypus cylindrus Fab. (Coleoptera: Platypodidae) on cork oak in Portugal. IOBC/Wprs Bull. 2010, 57, 87–95. [Google Scholar]
- Sousa, E.M.; Débouzie, D.D.; Pereira, H. The role of the insect Platypus cylindrus F. (Coleoptera, Platypodidae) in the decline of cork oak stands in Portugal. IOBC/Wprs Bull. 1995, 18, 24–37. [Google Scholar]
- Sousa, E.M.; Débouzie, D.D. Biological characteristics of Platypus cylindrus F. in Portugal. IOBC/Wprs Bull. 2002, 25, 75–83. [Google Scholar]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Strzałka, B.; Jankowiak, R.; Bilański, P.; Patel, N.; Hausner, G.; Linnakoski, R.; Solheim, H. Two new species of Ophiostomatales (Sordariomycetes) associated with the bark beetle Dryocoetes alni from Poland. MycoKeys 2020, 68, 23–48. [Google Scholar] [CrossRef] [PubMed]
- Kok, L.T. Lipids of ambrosia fungi in the life of the mutualistic beetles. In Insect-Fungus Symbioses; Batra, L.R., Ed.; Halsted Press: Sussex, UK, 1979; pp. 33–52. [Google Scholar]
- Plattner, A.; Kim, J.; Reid, J.; Hausner, G.; Lim, Y.W.; Yamaoka, Y.; Breuil, C. Resolving taxonomic and phylogenetic incongruence within species Ceratocystiopsis minuta. Mycologia 2009, 101, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, M.J.; Barnes, I.; de Beer, Z.W.; Roux, J.; Wingfield, B.D.; Taerum, S.J. Novel associations between ophiostomatoid fungi, insects and tree hosts: Current status—future prospects. Biol. Invasions 2017, 19, 3215–3228. [Google Scholar] [CrossRef]
- Baker, J.M. Ambrosia beetle and their fungi, with particular reference to Platypus cylindrus Fab. Symp. Soc. Gen. Microbiol. 1963, 13, 323–354. [Google Scholar]
- Cassier, P.; Lévieux, J.; Morelet, M.; Rougon, D. The Mycangia of Platypus cylindrus Fab. and P. oxyurus Dufour (Coleoptera: Platypodidae). Structure and associated fungi. J. Insect Physiol. 1996, 42, 171–179. [Google Scholar] [CrossRef]
- Sousa, E.M.; Tomaz, I.L.; Moniz, F.A.; Basto, S. La Répartition Spatiale des Champignons Associés à Platypus cylindrus Fab. (Coleoptera: Platypodidae). Phytopath. Medit. 1997, 36, 153. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/42685302 (accessed on 12 December 2021).
- Morelet, M. Une espèce nouvelle de Raffaelea, isolée de Platypus cylindrus, coléoptère xylomycétophage des chênes. Ann. Soc. Sci. Nat. Archéol. Toulon 1998, 50, 185–193. [Google Scholar]
- Henriques, J.; Inácio, M.L.; Sousa, E. Ambrosia fungi in the insect-fungi symbiosis in relation to cork oak decline. Rev. Iberoam. Micol. 2006, 23, 185–188. [Google Scholar] [CrossRef]
- Belhoucine, L.; Bouhraoua, R.T.; Meijer, M.; Houbraken, J.; Harrak, M.J.; Samson, R.A.; Equihua-Martinez, A.; Pujade-Villar, J. Mycobiota associated with Platypus cylindrus (Coleoptera: Curculionidae, Platypodidae) in cork oak stands of North West Algeria, Africa. Afr. J. Microbiol. Res. 2011, 5, 4411–4423. [Google Scholar] [CrossRef]
- Inácio, M.L.; Henriques, J.; Lima, A.; Sousa, E.B. Ophiostomatoid fungi associated with cork oak mortality in Portugal. IOBC/WPRS Bull. 2012, 76, 89–92. [Google Scholar]
- Soulioti, N.; Tsopelas, P.; Woodward, S.; Holdenrieder, O. Platypus cylindrus, a vector of Ceratocystis platani in Platanus orientalis stands in Greece. For. Pathol. 2015, 45, 367–372. [Google Scholar] [CrossRef]
- Bellahirech, A.; Inácio, M.L.; Woodward, S.; Ben Jamâa, M.L.; Nóbrega, F. Ophiostoma tsotsi and Ophiostoma quercus associated with Platypus cylindrus F. (Coleoptera: Curculionidae) in cork oak stands in Tunisia. For. Pathol. 2018, 49, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Badler, H. Pathogenicity of Ceratocystis spp. in oaks under stress. In Proceedings of an International Congress Recent Advances in Studies on Oak Decline, Selva di Fasano, Brindisi, Italy, 13–18 September 1992; Luisi, N., Lerario, P., Vannini, A., Eds.; pp. 31–37. [Google Scholar]
- Degreef, J. Isolation of Ophiostoma querci (Georgev.) Nannfeldt from declining oaks in Belgium: Selection techniques and pathogenicity test. In Proceedings of an International Congress Recent Advances in Studies on Oak Decline, Selva di Fasano, Brindisi, Italy, 13–18 September 1992; Luisi, N., Lerario, P., Vannini, A., Eds.; pp. 471–473. [Google Scholar]
- Delatour, C.; Menard, A.; Vautrot, A.; Simonin, G. Pathogenicity assessment of Ophiostomatales: Ophiostoma querci on oak compared to O. novo-ulmi on elm. In Proceedings of an International Congress Recent Advances in Studies on Oak Decline, Selva di Fasano, Brindisi, Italy, 13–18 September 1992; Luisi, N., Lerario, P., Vannini, A., Eds.; 1992; pp. 59–65. [Google Scholar]
- Harrington, T.C.; McNew, D.; Steimel, J.; Hofstra, D.; Farrell, R. Phylogeny and taxonomy of the Ophiostoma piceae complex and the Dutch elm disease fungi. Mycologia 2001, 93, 111–136. [Google Scholar] [CrossRef] [Green Version]
- de Beer, Z.W.; Wingfield, B.D.; Wingfield, M. The Ophiostoma piceae complex in the Southern Hemisphere: A phylogenetic study. Mycol. Res. 2003, 107, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, R.D.; de Beer, Z.W.; Jacobs, K.; Wingfield, B.D.; Wingfield, M.J. Multigene phylogenies define Ceratocystiopsis and Grosmannia distinct from Ophiostoma. Stud. Mycol. 2006, 55, 75–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Kim, S.H.; Lee, S.; Breuil, C. Distinguishing Ophiostoma ips and Ophiostoma montium, two bark beetle-associated sapstain fungi. FEMS Microbiol. Lett. 2003, 222, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Gorton, C.; Kim, S.H.; Henricot, B.; Webber, J.; Breuil, C. Phylogenetic analysis of the bluestain fungus Ophiostoma minus based on partial ITS rDNA and β-tubulin gene sequences. Mycol. Res. 2004, 108, 759–765. [Google Scholar] [CrossRef]
- Lim, Y.W.; Alamouti, S.M.; Kim, J.J.; Lee, S.; Breuil, C. Multigene phylogenies of Ophiostoma clavigerum and closely related species from bark beetle-attacked Pinus in North America. FEMS Microbiol. Lett. 2004, 237, 89–96. [Google Scholar] [CrossRef]
- Zhou, X.; de Beer, Z.W.; Wingfield, M.J. DNA sequence comparisons of Ophiostoma spp., including Ophiostoma aurorae sp. nov., associated with pine bark beetles in South Africa. Stud. Mycol. 2006, 55, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Wingfield, B.D.; Viljoen, C.D.; Wingfield, M.J. Phylogenetic relationships of ophiostomatoid fungi associated with Protea infructescences in South Africa. Mycol. Res. 1999, 103, 1616–1620. [Google Scholar] [CrossRef] [Green Version]
- Nel, W.J.; Wingfield, M.J.; de Beer, Z.W.; Duong, T.A. Ophiostomatalean fungi associated with wood boring beetles in South Africa including two new species. Antonie Leeuwenhoek 2021, 114, 667–686. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Duong, T.; Taerum, S.; Wingfield, M.; Zhou, X.; Yin, M.; de Beer, Z. Ophiostomatoid fungi associated with the spruce bark beetle Ips typographus, including 11 new species from China. Persoonia 2019, 42, 50–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowiak, R.; Solheim, H.; Bilański, P.; Marincowitz, S.; Wingfield, M.J. Seven new species of Graphilbum from conifers in Norway, Poland, and Russia. Mycologia 2020, 112, 1240–1262. [Google Scholar] [CrossRef] [PubMed]
- Roets, F.; De Beer, Z.W.; Crous, P.W.; Dreyer, L.L.; Wingfield, M.J. Ophiostoma gemellus and Sporothrix variecibatus from mites infesting Protea infructescences in South Africa. Mycologia 2008, 100, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.; Geiser, D.M.; Hawksworth, D.L.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Massoumi Alamouti, S.; Tsui, C.M.; Breuil, C. Multigene phylogeny of filamentous ambrosia fungi associated with ambrosia and bark beetles. Mycol. Res. 2009, 113, 822–835. [Google Scholar] [CrossRef]
- Zanzot, J.W.; de Beer, Z.W.; Eckhardt, L.G.; Wingfield, M.J. A new Ophiostoma species from loblolly pine roots in the southeastern United States. Mycol. Progress 2010, 9, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P.A. Chromotaxia seu nomenclator colorum polyglottus adclitis speciminibus coloratis ad botanicorum et zoologorum. Patavii 1891, 22. [Google Scholar]
- Riddell, R.W. Permanent stained mycological preparations obtained by slide culture. Mycologia 1950, 42, 265–270. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.J.; Fung, S.; Breuil, C. A PCR-RFLP marker distinguishing Ophiostoma clavigerum from morphologically similar Leptographium species associated with bark beetles. Can. J. Bot. 2003, 81, 1104–1112. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kimura, K.; Ito, S. Genetic characterization of Raffaelea quercivora isolates collected from areas of oak wilt in Japan. Mycoscience 2010, 51, 310–316. [Google Scholar] [CrossRef]
- Harrington, T.C.; Aghayeva, D.N.; Fraedrich, S.W. New combinations in Raffaelea, Ambrosiella, and Hyalorhinocladiella, and four new species from the redbay ambrosia beetle. Xyleborus glabratus. Mycotaxon 2010, 111, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: Genes for Phylogenetics; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and its near relatives. In the Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematic; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1998; pp. 225–233. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Kusaba, M.; Tsuge, T. Phylogeny of Alternaria fungi known to produce host-specific toxins on the basis of variation in internal transcribed spacers of ribosomal DNA. Curr. Genet. 1995, 28, 491–498. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1330. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, H.; Begerow, D.; Oberwinkler, F. Identification of the ambrosia fungus of Xyleborus monographus and X. dryographus (Coleoptera: Curculionidae, Scolytinae). Mycol. Prog. 2004, 3, 95–102. [Google Scholar] [CrossRef]
- Gebhardt, H.; Oberwinkler, F. Conidial development in selected ambrosial species of the genus Raffaelea. Antonie Leeuwenhoek 2005, 88, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing 524 platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Inácio, M.L.; Marcelino, J.; Lima, A.; Sousa, E.; Nóbrega, F. Raffaelea quercina sp. nov. associated with cork oak (Quercus suber L.) decline in Portugal. Forests 2021, 12, 513. [Google Scholar] [CrossRef]
- Musvuugwa, T.; de Beer, Z.W.; Duong, T.A.; Dreyer, L.L.; Oberlander, K.C.; Roets, F. New species of Ophiostomatales from Scolytinae and Platypodinae beetles in the Cape Floristic Region, including the discovery of the sexual state of Raffaelea. Antonie Leeuwenhoek J. Microbiol. 2015, 108, 933–950. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.G.; Harrington, T.C.; Ranger, C. First report of a sexual state in an ambrosia fungus: Ambrosiella cleistominuta sp. nov. associated with the ambrosia beetle Anisandrus maiche. Botany 2017, 95, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Nkuekam, G.K.; DeBeer, W.Z.; Wingfield, M.J.; Roux, J. A diverse assemblage of Ophiostoma species, including two new taxa on eucalypt trees in South Africa. Mycol. Progress 2011, 11, 515–533. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T. Fungal associates of European bark beetles with special emphasis on the ophiostomatoid fungi. In Bark and Wood Boring Insects in Living Trees in EUROPE, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H.F., Eds.; Springer: Amsterdam, The Netherlands, 2007; pp. 181–235. [Google Scholar]
- Harrington, T.C.; Fraedrich, S.W.; Aghayeva, D.N. Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Kim, S.; Harrington, T.C.; Lee, J.C.; Seybold, S.J. Leptographium tereforme sp. nov. and other Ophiostomatales isolated from the root-feeding bark beetle Hylurgus ligniperda in California. Mycologia 2011, 103, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Desprez-Loustau, M.; Marçais, B.; Nageleisen, L.-M.; Piou, D.; Vannini, A. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef] [Green Version]
- de Beer, W.; Procter, M.; Wingfield, M.J.; Marincowitz, S.; Duong, T.A. Generic boundaries in the Ophiostomatales reconsidered and revised. Stud. Mycol. 2022, 101, 57–120. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Source a | Country | Associated Insect or Host | GenBank Accession nrs. b | |||
|---|---|---|---|---|---|---|---|
| nSSU rDNA | nLSU rDNA | 5.8S-ITS2 | TUB | ||||
| C. quercina sp. nov. | PC05.005, C2510 | Portugal | Quercus suber | JF909502 | JF909532 | JF909518 | MZ450136 |
| C. quercina sp. nov. | PC05.006, C2511 | Portugal | Platypus cylindrus | JF909503 | JF909533 | JF909519 | MZ450137 |
| C. quercina sp. nov. | PC06.022, C2519 | Portugal | P. cylindrus | JF909504 | JF909534 | JF909520 | MZ450138 |
| C. quercina sp. nov. | PC06.032, C2508 | Portugal | P. cylindrus | JF909505 | JF909535 | JF909521 | MZ450139 |
| C. quercina sp. nov. | PC06.034, C2507 | Portugal | P. cylindrus | JF909506 | JF909544 | JF909522 | MZ450140 |
| C. quercina sp. nov. | PC07.004, C2517 | Portugal | Q. suber | JF909507 | JF909545 | JF909523 | MZ450141 |
| C. quercina sp. nov. | PC07.007, C2509 | Portugal | P. cylindrus | JF909508 | JF909536 | JF909524 | MZ450142 |
| O. abietinum | CBS125.89 | Mexico | Abies vejari | AF155685 | AY924382 | ||
| O. abietinum | CMW1468 | Canada | Dendroctonus ponderosa | AY280468 | |||
| O. abietinum | CMW109 | USA | Pinus echinata | AY280469 | |||
| O. africanum | CMW1104 | South Africa | Protea caffra | DQ316147 | DQ316162 | ||
| O. ambrosium | CBS 210.64 | AF137285 | |||||
| O. araucariae | CMW671 | Chile | Araucaria araucaria | DQ294373 | |||
| O. araucariae | CBS 114.68 | Chile | Araucaria araucaria | KU184289 | |||
| O. arborea | TUB F4270 | Germany | Ips typographus | AY497511 | |||
| O. aurorae | CMW19364 | South Africa | Pinus elliottii Hylurgus angustatus | DQ396798 | DQ396802 | ||
| O. abieticola | YCC478 | Japan | Ips subelongatus | GU134156 | |||
| O. bicolor | TUB F4269 | Germany | Ips typographus | AY497512 | |||
| O. fasciatum | UM56 | Brit. Columbia | Pseudotsuga menziesii | EU913680 | EU913759 | ||
| O. floccosum | AU55-6 | AF234836 | |||||
| O. floccosum | CNB 117A | Spain | Pinus pinaster | ||||
| O. floccosum | CMW 171 | USA | Pinus ponderosa | DQ296087 | |||
| O. floccosum | 387N | Canada, CC | unknown | AF139810 | |||
| O. fusiforme | CMW9968 | Azerbaijan | Populus nigra | DQ294354 | AY280461 | ||
| O. fusiforme | CMW7131 | Austria | Quercus petraea | AY280497 | AY280464 | ||
| O. ips | CBS 137.36 | USA | Ips sp. | EU913724 | |||
| O. ips | CMW7075 | USA | Ips integer | DQ294381 | DQ296101 | ||
| O. ips | KUC2120 | Pinus sp. | AY172021 | ||||
| O. karelicum | CMW23099 | Russia | EU443773 | ||||
| O. lunatum | CBS 112928, CMW10564 | Austria | Larix decidua | AY280486 | AY280467 | ||
| O. longisporum | WIN(M)48 | HQ634831 | |||||
| O. minus | WIN(M)861 | HQ634820 | |||||
| O. nigrocarpum | CMW651 | USA | Pseudotsuga menziesii | DQ294356 | AY280480 | ||
| O. nigrocarpum | Ci-203 | Chile | Pinus radiata | EF506941 | |||
| O. nigrocarpum | CMW1468 | Canada | Dendroctonus ponderosae | AF484457 | |||
| O. novo-ulmi | CMW10573 | Austria | Picea abies | DQ294375 | DQ296095 | ||
| O. phasma | CMW20676 | South Africa | Protea laurifolia | ||||
| O. piceae | AU100-1 | AF234837 | |||||
| O. piceae | CMW8093 | Canada | Tetropium sp. | DQ296091 | |||
| O. piceae | JCM6016 | AB007663 | |||||
| O. pilliferum | CMW7879 | South Africa | Pinus sylvestris | DQ294377 | |||
| O. pilliferum | CBS 129.32 | Germany | AJ243295 | AF221628 | |||
| O. pilliferum | CBS 158.74 | Chile | DQ470955 | ||||
| O. quercus | CMW3110 | USA | Junlans cinerea | DQ294376 | DQ296096 | ||
| O. quercus | CMW 2467 | France | Quercus sp. | ||||
| O. quercus | TUB F4272 | Germany | Xyleborus monographus | AY497515 | |||
| O. retusum | ATCC22324 | USA | HQ634841 | ||||
| O. ulmi | CBS 298.87 | Netherlands | M83261 | ||||
| O. ulmi | CMW1462 | USA | Ulmus procera | DQ294374 | DQ296094 | ||
| A. macrospora | CBS 367.53, C2231 | Sweden | Ips acuminatus | EU170284 | |||
| A. tingens | CBS 366.53 | Sweden | Xyleborus glabratus gallery | EU170277 | |||
| O. gossypinum var. robustum | MUCL18357 | Spain | - | AY924388 | |||
| O. stenoceras | UCB 57.013 | - | M85054 | ||||
| O. stenoceras | CMW2530 | Colombia | Eucalyptus grandis | AF484460 | |||
| O. stenoceras | CBS 139.51 | DQ836904 | |||||
| O. stenoceras | CBS 237.32 CMW3202 | Norway | Pinus sp. | DQ296074 | |||
| O. stenoceras | CBS 237.32 | Norway | Pinus sp. | AY280471 | |||
| O. stenoceras | CMW2524 | South Africa | Acacia mearnsii | AF484459 | |||
| O. stenoceras | Germany | Natrix natrix | |||||
| O. stenoceras | CMW4007 | Colombia | Eucalyptus sp. soil | AF484464 | |||
| O. stenoceras | Germany | Python regius | |||||
| O.triangulosporum | DSMZ4934 | Brasil | Araucaria araucaria | AY934525 | |||
| O.torulosum | TUB F3258 | Germany | Trypodendron lineatum | AY497517 | |||
| S. lignivora | CMW18600 | South Africa | Eucalyptus sp. | EF139119 | EF139104 | ||
| Ceratocystiopsis sp. | AM434-K2G-1 | USA | Pinus sp. | KT264634 | |||
| Ceratocystiopsis sp. (Cop. minuta-like) | Cop. sp. 1i | Brit. Columbia | D. ponderosae gallery | EU913667 | EU913746 | ||
| Ceratocystiopsis sp. | Cop. sp3i | Brit. Columbia | Picea glauca, Ips perturbatus | EU913676 | EU913755 | ||
| Ceratocystiopsis sp. (Cop. manitobense-like) | Cop. sp3ii | Brit. Columbia | Ips perturbatus | EU913717 | EU913756 | ||
| Ceratocystiopsis sp. (Cop. minuta-like) | YCC329 | Japan | L. kaempferi, Ips sp. | EU913671 | EU913750 | ||
| C. brevicomes | CBS 333.97 | USA | Dendroctonus brevicomis | HQ202311 | EU913683 | EU913761 | |
| C. collifera | WIN(M)908 | HQ634832 | |||||
| C. collifera | CBS 126.89 | Mexico | Dendroctonus valens | EU913681 | MH862160 | EU913760 | |
| C. concentrica | WIN(M)53 | Canada | HQ634849 | ||||
| C. concentrica | WIN(M) 71-07 | Canada | AF135571 | ||||
| C. manitobensis | UM237 | Canada | Ips perturbatus | EU984266 | DQ268607 | DQ268610 | |
| C. manitobensis | UM237 | Canada | Ips perturbatus | EU913714 | |||
| C. manitobensis | WIN(M)237 | Canada | HQ634850 | ||||
| C. manitobensis | CW13792 | Canada | Pinus resinosa | DQ294358 | DQ296078 | ||
| C. minuta | RJ5095 (UM1533) | Poland | Picea sp. Ips typographus | EU913698 | |||
| C. minuta | RJ191(UM 1535) | Poland | Picea sp. Ips typographus | EU913700 | |||
| C. minuta | RJ705 (UM 1532) | Poland | Picea abies Ips typographus | EU913656 | EU913736 | ||
| C. minuta | CBS463.77 | Mexico, USA | Picea engelmanii | EU913645 | EU913725 | ||
| C. minuta | WIN(M)1532 | Canada | HQ634827 | ||||
| C. minuta | YCC139 | Japan | Picea jezoensis Ips sp. | EU913732 | |||
| C. minuta-bicolor | South Africa | Pinus sp. | DQ294359 | ||||
| C. minuta-bicolor | UAMH9551 | Canada | Pinus contorta | ||||
| C. minuta-bicolor | WIN(M)480 | Canada | HQ634848 | ||||
| C. minuta-bicolor | CBS635.66 (UM844) | USA | Pinus contorta | EU913745 | |||
| C. minuta-bicolor | UM 480 | Canada | Pinus contorta | EU913705 | |||
| C. minima | WIN(M)85 | Canada | HQ634856 | ||||
| C. minima | CBS 182.86 | USA | Pinus banksiana | EU913663 | EU913706 | EU913743 | |
| C. pallidobrunnea | UM51 | Canada | Populus tremuloides | EU913682 | |||
| C. pallidobrunnea | WIN(M)51 | Canada | HQ634842 | ||||
| C. parva | JR71-21 | Canada | HQ595735 | ||||
| C. ranaculosa | CMW13940 | USA | Pinus echinata | DQ294357 | DQ296077 | ||
| C. ranaculosa | WIN(M)919 | HQ634840 | |||||
| C. ranaculosa | CBS 216.88 | USA | |||||
| C. rollhanseniana | CW13791 | Norway | Pinus sylvestris | DQ294362 | DQ296082 | ||
| C. rollhanseniana | WIN(M)110 | Canada | HQ634834 | EU913719 | |||
| C. rollhanseniana | UM110 | Norway | Pinus sylvestris | EU913758 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inácio, M.L.; Marcelino, J.; Lima, A.; Sousa, E.; Nóbrega, F. Ceratocystiopsis quercina sp. nov. Associated with Platypus cylindrus on Declining Quercus suber in Portugal. Biology 2022, 11, 750. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050750
Inácio ML, Marcelino J, Lima A, Sousa E, Nóbrega F. Ceratocystiopsis quercina sp. nov. Associated with Platypus cylindrus on Declining Quercus suber in Portugal. Biology. 2022; 11(5):750. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050750
Chicago/Turabian StyleInácio, Maria L., José Marcelino, Arlindo Lima, Edmundo Sousa, and Filomena Nóbrega. 2022. "Ceratocystiopsis quercina sp. nov. Associated with Platypus cylindrus on Declining Quercus suber in Portugal" Biology 11, no. 5: 750. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050750