Inhibitory Effect and Mechanism of Scutellarein on Melanogenesis


1
Bionics Program, Tokyo University of Technology Graduate School, 1404-1 Katakuramachi, Hachioji City, Tokyo 192-0982, Japan
2
School of Bioscience and Biotechnology, Tokyo University of Technology, 1404-1 Katakuramachi, Hachioji City, Tokyo 192-0982, Japan
*
Author to whom correspondence should be addressed.
Cosmetics 2021, 8(1), 15; https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010015
Submission received: 9 January 2021
/
Revised: 31 January 2021
/
Accepted: 9 February 2021
/
Published: 15 February 2021
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fairer skin is preferred in many Asian countries and there is a high demand for skin whitening and lightening products. However, in recent years, problems related to the safety of using whitening agents have emerged. This study demonstrates that plant-derived scutellarein effectively inhibits melanogenesis in B16 melanoma cells. However, baicalein, which is similar to scutellarein in its chemical structure, does not show any inhibitory effect on melanogenesis. Cellular tyrosinase activity is decreased by scutellarein in a dose-dependent manner. No cytotoxicity is observed at the effective concentration range. Additionally, both the protein and mRNA levels of tyrosinase are significantly decreased by scutellarein. Further, the risk of leukoderma development also is determined by evaluating the production of free hydroxyl radicals (˙OH); scutellarein treatment does not induce ˙OH production. Scutellarein shows no risk of causing leukoderma. Our results suggest that scutellarein or plant extracts containing high concentrations of scutellarein have the potential to inhibit melanin production and serve as cosmetic skin-lightening agents.
1. Introduction
Accompanying an increase in people’s awareness of their own beauty and esthetic appeal, an increasing number of people are pursuing cleaner and fairer skin [1]. Recently, the demand for whitening products, mainly skin-lightening cosmetics, has increased [2]. According to the results of a cosmetics market survey conducted by Japan’s Fuji Keizai in 2018 [3], consumers have high expectations for medicinal cosmetics with anti-freckle and anti-pigmentation properties, and the market demand for skin-lightening cosmetics is expected to gradually increase in the future. However, the leukoderma incident of Kanebo in 2013 [4] brought more attention to the safety of cosmetic whitening ingredients. Approximately 20 active ingredients, including hydroquinone-β-d-glucoside (arbutin, ARB), 4-n-butylresorcinol (4BR), 3-O-ethyl ascorbic acid, tranexamic acid, and 4-methoxysalicylic acid potassium salt (4MSK), have been approved for use in quasi-drug cosmetics in Japan [5,6,7,8,9]. However, rhododendrol [(±) 4-(3-hydroxybutyl) phenol; 4HP], and magnolignan (2,2-dihydroxy-5,5-dipropyl-biphenyl; ML) react with tyrosinase substrates to generate high amounts of free hydroxyl radicals (˙OH), thereby exerting toxic effects on melanocytes [10]. Thus, over time, melanocyte destruction could occur, and leukoderma may become permanent [11]. Therefore, there is currently an increased interest in the development of new, safe, and effective whitening agents from traditional herbs [12]. Recently, researchers have focused on the whitening properties of plant-derived compounds and have found that compounds such as the ethanol extract of Angelica gigas [13] and plant-derived luteolin [14] have an inhibitory effect on melanogenesis.
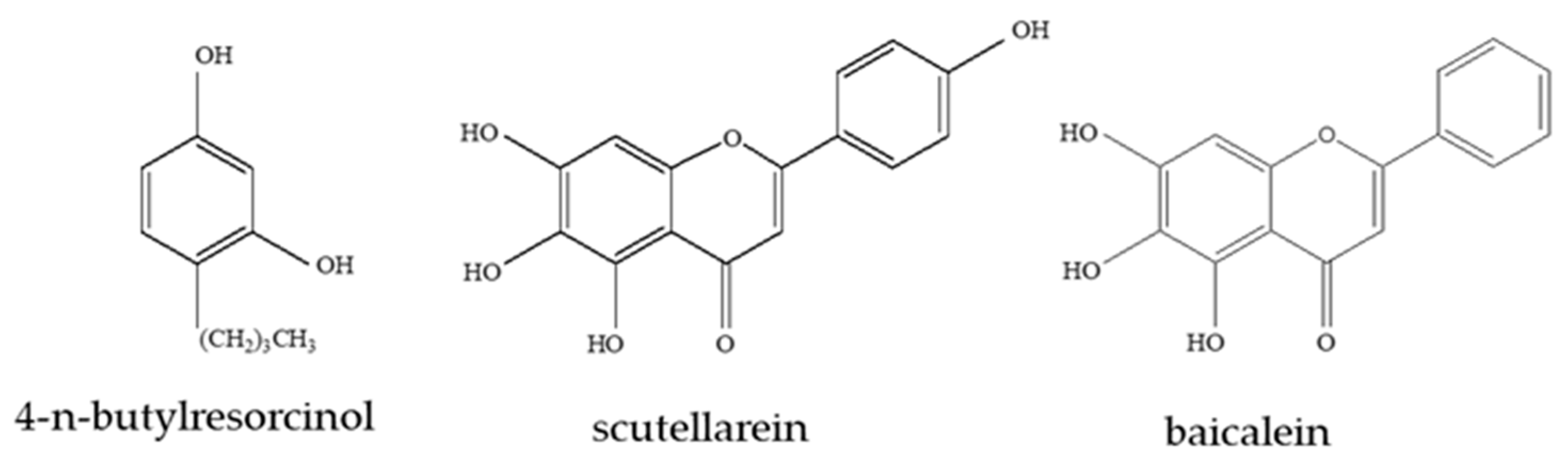
Scutellarein and baicalein (Figure 1) are bioactive flavones purified from the medicinal plant Scutellaria baicalensis Georgi (SBG) [15], which has been used for the treatment of various inflammatory diseases, hepatitis, tumors, and diarrhea, in East Asian countries [16]. Low levels of scutellarein are observed in the aerial part of SBG; scutellarein exhibits high antioxidant activity [17] and, consequently, is used as a medicine to treat inflammation and neurological diseases [18]. Baicalein mainly accumulates in the roots of SBG and exhibits free radical-scavenging activity [19]. Additionally, studies have shown that SBG plant extracts inhibit melanogenesis [20]. However, there is no related research on the whitening effects of scutellarein and baicalein.
Hyperpigmentation of solar lentigo arises primarily from increased melanogenesis of existing melanocytes in the basal layer of the epidermis, as well as from increased melanosome complexes in keratinocytes [21]. Although melanin is the main culprit in skin darkening, the precise reason is the accumulation of melanin in the keratinocytes and melanocytes in the perinuclear area—like “caps” on the nucleus—which helps in protecting the DNA from ultraviolet rays [22]. The biosynthesis process is controlled by a cascade of tyrosinase, tyrosinase related protein-1, and tyrosinase related protein-2 (TRP-1 and TRP-2) and is very complicated. Tyrosine is hydroxylated to 3-(3,4-dihydroxyphenyl)-alanine (DOPA) by tyrosinase, which is the rate-limiting step in melanogenesis [23]. DOPA is then oxidized to dopaquinone, which undergoes autoxidation to form dopachrome, then catalyzed by TRP-2, resulting in exhibition of dopachrome tautomerase activity, to dihydroxyindole carboxylic acid (DHICA). DHICA is oxidized by TRP-1 to indolequinone, which eventually forms eumelanin. Concurrently, pheomelanin synthesis is accomplished through cysteine. Additionally, researchers found that unlike tyrosinase and TRP-1, which are mostly distributed in mature melanosomes, TRP-2 is concentrated in the perinuclear area [24]. Microphthalmia-associated transcription factor (MITF) is a transcription factor known to be essential for melanocyte development. Additionally, it regulates the transcription of three major pigmentation enzymes: tyrosinase, TRP-1, and TRP-2 [25].
Here, scutellarein and baicalein are used as experimental objects to explore their effects on melanin production in B16 cells. During the present study, 4-n-butylresorcinol (Figure 1) is used as a positive control to examine the effect of scutellarein on the tyrosinase activity, cytotoxicity, and melanin content of B16 cells. The effects of scutellarein on the expression of tyrosinase also are explored. Further, the potential of scutellarein to cause leukoderma is predicted by the measurement of the levels of free hydroxyl radicals (˙OH) and, also, its safety for use as a skin-lightening agent is evaluated.
2. Materials and Methods
2.1. Materials
Dulbecco’s modified Eagle’s medium (DMEM) + GlutaMAXTM and 0.05% trypsin-ethylenediaminetetraacetic acid (EDTA) were purchased from Gibco (Thermo Fisher Scientific, Waltham, MA, USA). Phosphate-buffered saline (PBS) was purchased from TaKaRa Bio (Shiga, Japan). Fetal bovine serum, α-melanocyte stimulating hormone (α-MSH), and mushroom tyrosinase were purchased from Sigma-Aldrich (St. Louis, MO, USA). The cell counting kit-8 was purchased from Dojindo Laboratories (Kumamoto, Japan) and 20% HCl, NaCl, NaOH, 2-amino-2-hydroxymethyl-1,3-propanediol (Tris), glycine, 3-(3,4-dihydroxyphenyl)-l-alanine (L-DOPA), sodium dodecyl sulfate (SDS), sodium deoxycholate, 2-mercaptoethanol, dimethyl sulfoxide (DMSO), and rhododendrol were purchased from FUJIFILM Wako Pure Chemical (Osaka, Japan). Scutellarein and 4-n-butylresorcinol were purchased from the Tokyo Chemical Industry (Tokyo, Japan), and Triton X-100 and Tween 20 were procured from Bio-Rad Laboratories (Alfred Nobel Drive Hercules, CA, USA).
2.2. Preparation of Test Compound
Scutellarein and baicalein solutions and the 4-butylresorcinol solution were prepared with dimethyl sulfoxide (DMSO) as the experimental and positive control groups, respectively.
2.3. Cell Culture
B16 mouse melanoma cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum at 37 °C in a 5% CO2 incubator. The cells were harvested using trypsin-EDTA.
2.4. Melanin Content Assay
B16 cells were cultured in a 100-mm dish (AGC Techno Glass, Shizuoka, Japan) at a concentration of 2.5 × 105 cells/mL for 24 h, then treated with the specific test compound and 20 ng/mL α-melanocyte stimulating hormone (α-MSH) at 37 °C in a 5% CO2 incubator for 72 h. Cells were exposed to trypsin and shaken gently to remove all cells from the dish. The obtained solution was clarified by centrifugation at 10,000 rpm for 10 min, the supernatant was removed, and the pellet was dissolved in an NaOH solution per the number of cells. Absorbance of the cell suspension was measured at 475 nm using the multi-detection microplate reader.
2.5. Cellular Viability Assay
B16 cells were cultured in a 24-well microplate (Corning Coster) at a concentration of 1.25 × 105 cells/mL for 24 h and treated with the specific test compound at 37 °C in a 5% CO2 incubator for 48 h. Cell counting kit-8 solution was added and the absorbance of the water-soluble formazan dye after 0 h and 2 h was measured at 450 nm using a multi-detection microplate reader. Cell viability was expressed as the difference between the two absorbance values.
2.6. Tyrosinase Activity Assay
B16 cells were cultured in a 96-well microplate (Corning Coster, Corning, NY, USA) at a concentration of 1.0 × 105 cells/mL for 24 h and treated with the specific test compound at 37 °C in a 5% CO2 incubator for 48 h. Triton X-100, 3-(3,4-dihydroxyphenyl)-l-alanine (DOPA), and phosphate-buffered saline (PBS) were added, then the absorbance of dopachrome after 0 h and 2 h was measured at 475 nm using a multi-detection microplate reader (PowerScan HT; DS Pharma Biomedical Co. Ltd., Osaka, Japan). Tyrosinase activity was expressed as the difference between the two absorbance values.
2.7. Mushroom Tyrosinase Activity Assay
Phosphate-buffered saline (PBS) (79 μL), the test compound (1 μL), and 10 mM 3-(3,4-dihydroxyphenyl)-l-alanine (10 μL) with 100 U/mL mushroom tyrosinase (10 μL) were added to each 96-well plate. The absorbance of dopachrome after 0 min and 10 min was measured at 475 nm using a multi-detection microplate reader. Mushroom tyrosinase activity was expressed as the difference between the two absorbance values.
2.8. Western Blotting Assay
B16 cells cultured in a 100-mm dish at a concentration of 2.5 × 105 cells/mL for 24 h were treated with the specific test compound at 37 °C in a 5% CO2 incubator for 72 h. Cells were washed with phosphate-buffered saline (PBS) three times and lysed with 2% sodium dodecyl sulfate (SDS) and 1% sodium deoxycholate (0.5 mL). Supernatants were collected as whole-cell lysates, and protein concentrations were determined using the BCA protein assay reagent (Thermo Fisher Scientific, Rockford, MA, USA). Each sample was adjusted to the same protein concentration and heated to 95 °C for 5 min. Proteins were separated by SDS polyacrylamide gel electrophoresis (SDS-PAGE) using a 4–20% gel (TEFCO, Hachioji, Japan), and then transferred to polyvinylidene fluoride (PVDF) membranes (Merck Millipore, Cork, Ireland). The membrane was blocked with 5% skim milk in tris-buffered saline (TBS) at room temperature for 30 min, then incubated with a mouse anti-β-actin antibody (diluted 1:1000; Proteintech Group, Rosemont, IL, USA) and a final anti-rabbit serum recognizing the anti-tyrosinase antibody (diluted 1:1000; Thermo Scientific) or anti-rabbit serum recognizing the microphthalmia-associated transcription factor (MITF) antibody (diluted 1:1000; Proteintech Group) with TBS at room temperature for 1 h. The membrane was washed three times with tris buffered saline with tween 20 (TBST) for 30 min and then incubated with a peroxidase-labeled anti-mouse antibody (GE Healthcare, Marlborough, MA, USA) for 30 min at room temperature. The membrane was washed three times with TBST for 3 h, and a western blotting detection reagent (GE Healthcare) was added; the membrane was then incubated for 5 min at room temperature. Finally, immunoreactive bands were visualized using a Lumino Graph (ATTO, Tokyo, Japan).
2.9. Reverse Transcription qPCR (PT-qPCR) Assay
B16 cells cultured in 35-mm dishes at a concentration of 1.5 × 105 cells/mL for 24 h were treated with the specific test compound at 37 °C in a 5% CO2 incubator. RNA was extracted from cells after 6, 12, and 24 h. Total RNA was prepared using the RNeasy Protect Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s protocol. Total RNA was reverse transcribed to cDNA using the One Step TB GreenTM PrimeScriptTM RT-PCR Kit II (TaKaRa Bio) and oligo (dT) primers (QIAGEN). Real-time PCR was performed using the QuantStudio® 5 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). A reverse transcription reaction was carried out under the following conditions: 42 °C for 5 min and 95 °C for 10 s; PCR amplification: 95 °C for 5 s and 60 °C for 34 s; dissociation protocol: 95 °C for 15 s, 60 °C for 1 min, and 95 °C for 15 s. Expression levels of the tyrosinase gene were normalized to those of the glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH). Relative changes in mRNA expression levels were calculated with the relative calibration method, and the levels were normalized to that of GAPDH. The experiment was performed in triplicate.
2.10. Measurements of Hydroxyl Radical (˙OH)
Phosphate-buffered saline (PBS) (78 μL), the specific test compound (2 μL), and hydroxyphenyl fluorescein (HPF; 5 μmol/L, Goryo Chemical, Sapporo, Japan), with or without 100 U/mL mushroom tyrosinase (10 μL), were added to each 96-well plate and allowed to react at 37 °C. The fluorescence intensity of the compound generated by free hydroxyl radical (˙OH) (excitation, 485 nm; fluorescence, 528 nm) was measured after 30 min using the multi-detection microplate reader.
2.11. Data Analysis
Data are expressed as mean ± standard deviation (SD). An unpaired t-test (two-tailed) was performed to compare the data of each test compound to the control data using an Excel for Microsoft Office Professional Plus 2016; p < 0.05 was considered statistically significant.
3. Results
3.1. Effects of Scutellarein and Baicalein on the Melanin Content of B16 Cells
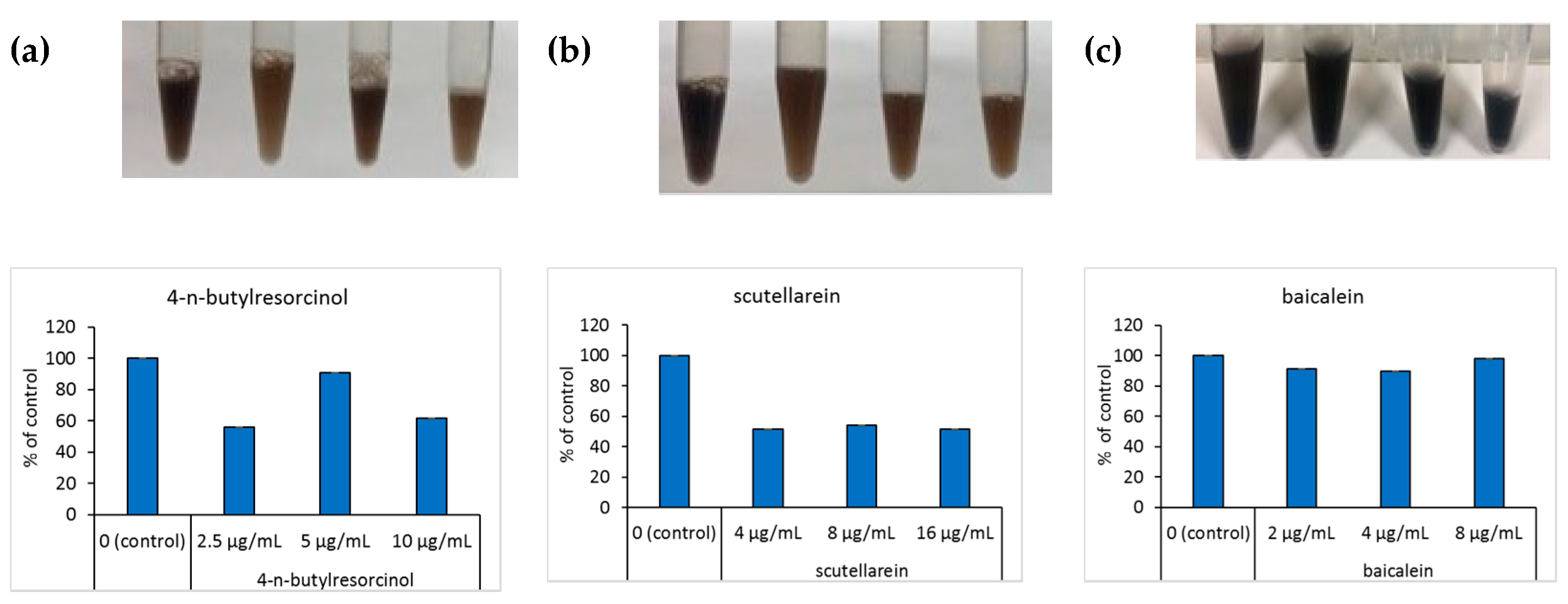
When B16 cells were incubated with scutellarein and α-melanocyte stimulating hormone (α-MSH), the cell pellet dissolved in the NaOH solution was visibly black. The color of the dissolved cell pellet was compared using NaOH at different concentrations, with the absorbance measured at 475 nm. Melanin content was significantly decreased in cells treated with 4-n-butylresorcinol and scutellarein compared with the untreated control; a clear dose-dependent decrease in cellular melanogenesis was observed (Figure 2a,b). To contrast, the color of the cell pellet dissolved in NaOH, corresponding to baicalein treatment at different concentrations, did not show any significant change, and there was no difference in absorbance (Figure 2c).
3.2. Effects of Scutellarein on the Tyrosinase Activity in the Well
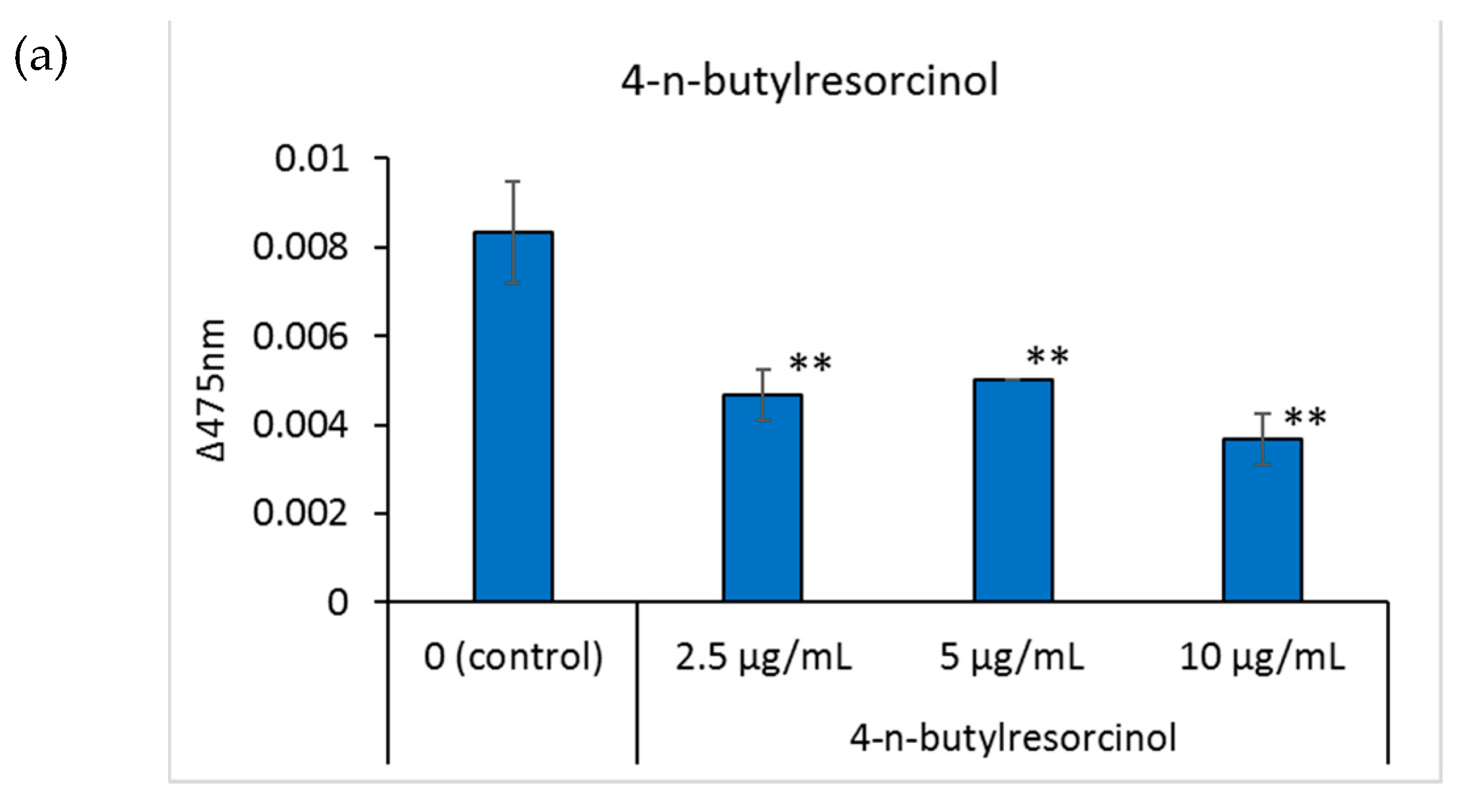
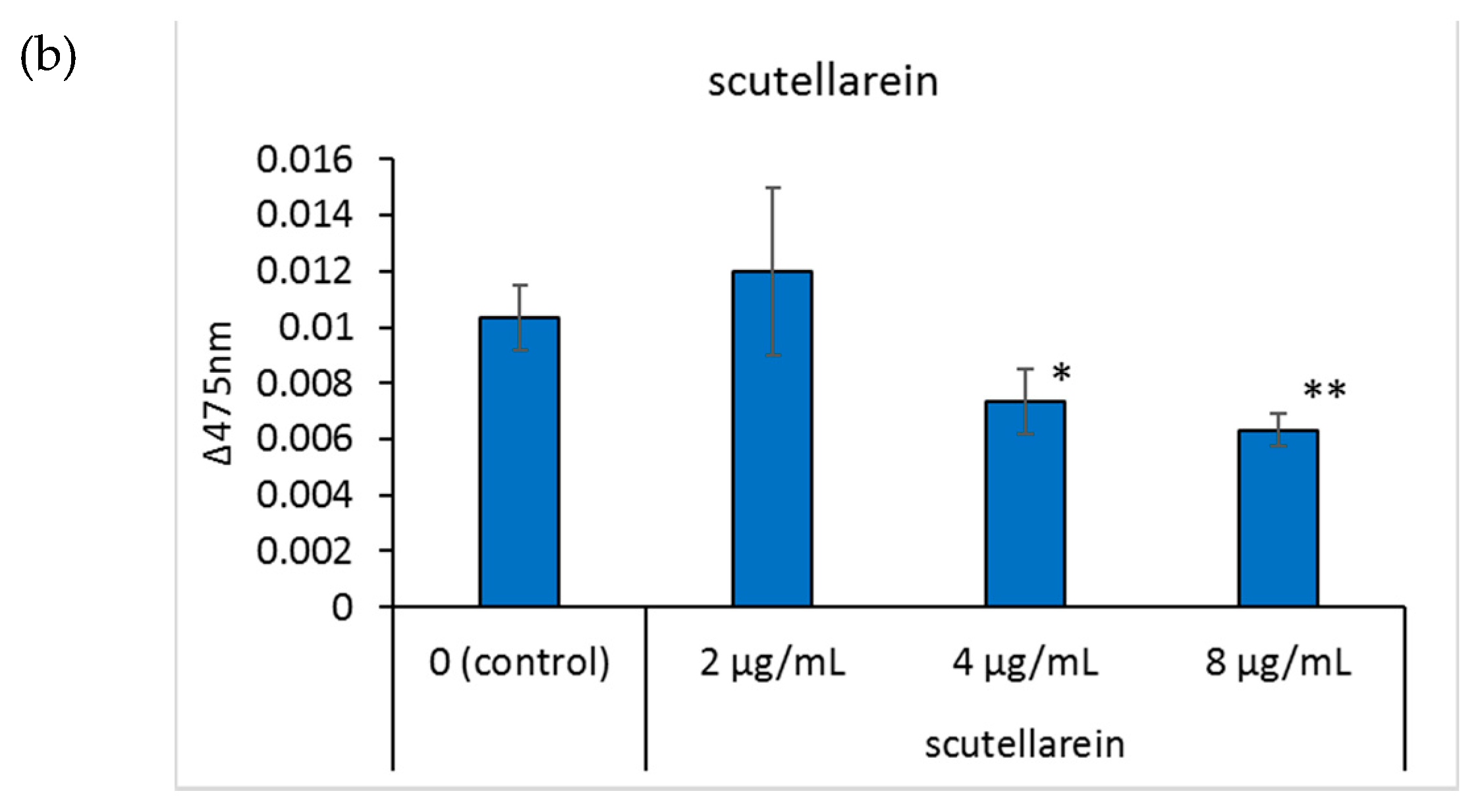
Tyrosinase is an important enzyme for melanin production, and its regulation is one of the factors affecting skin pigmentation [25]. Tyrosinase activity in the well was determined by adding 3-(3,4-dihydroxyphenyl)-l-alanine to the cells and measuring the absorbance of the produced dopachrome at 475 nm. Regarding the positive control, 4-n-butylresorcinol dose-dependently inhibited tyrosinase activity at 2.5, 5, and 10 μg/mL (Figure 3a). Compared to the control group, when the concentration of scutellarein in the medium was 4 and 8 μg/mL, scutellarein dose-dependently inhibited tyrosinase activity (Figure 3b).
3.3. Effects of Scutellarein on the Viability of B16 Cells
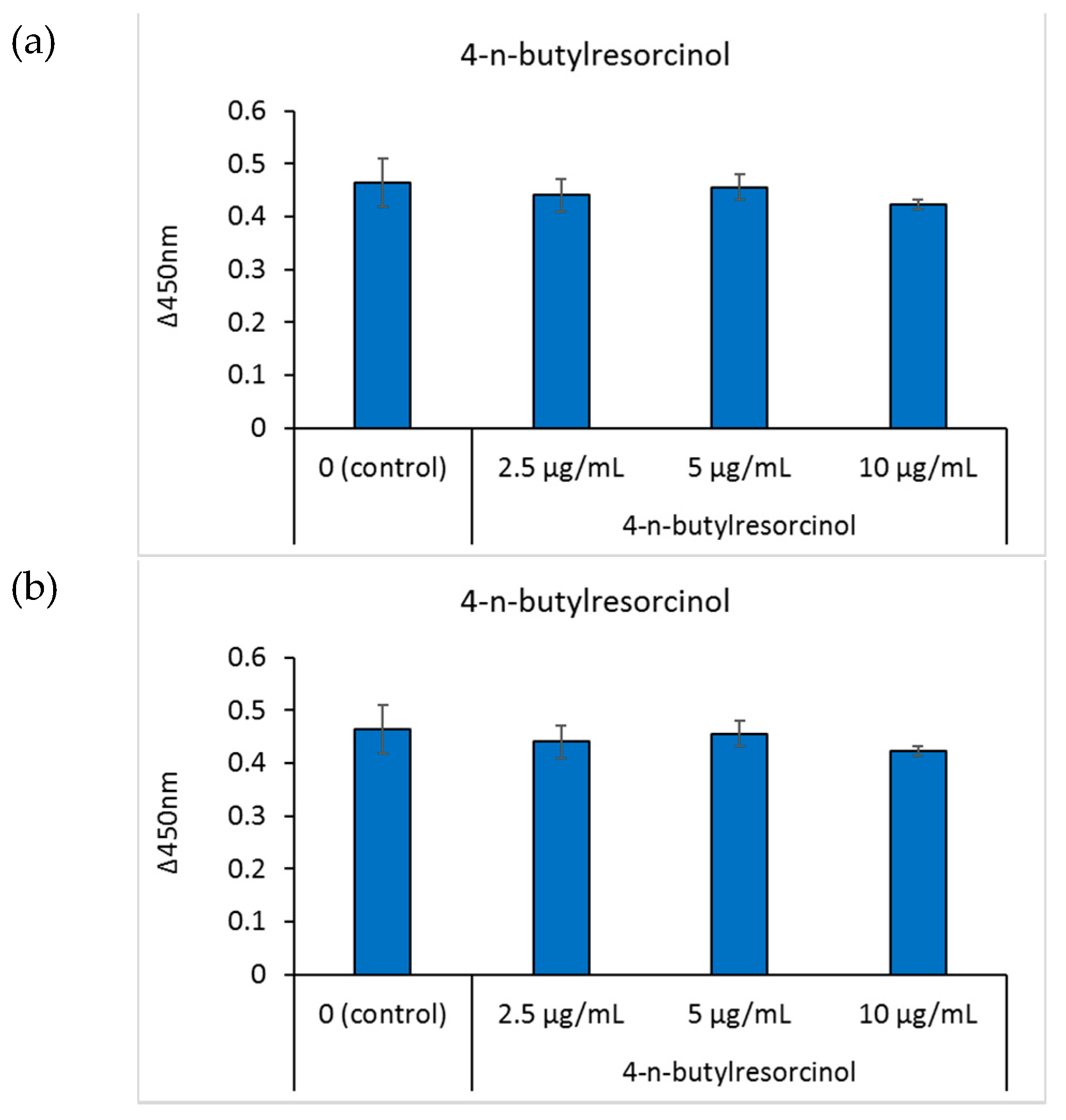
Cell viability was determined by reacting the cells with the cell counting kit-8 solution and measuring the absorbance of the water-soluble formazan dye at 450 nm. After cells were cultured in media containing different concentrations of 4-n-butylresorcinol for 48 h, there was no tendency toward a decreased cell viability, indicating that 4-n-butylresorcinol and scutellarein exhibited no cytotoxicity within the experimental concentration range (Figure 4a,b).
3.4. Effect of Scutellarein and Baicalein on Mushroom Tyrosinase Activity
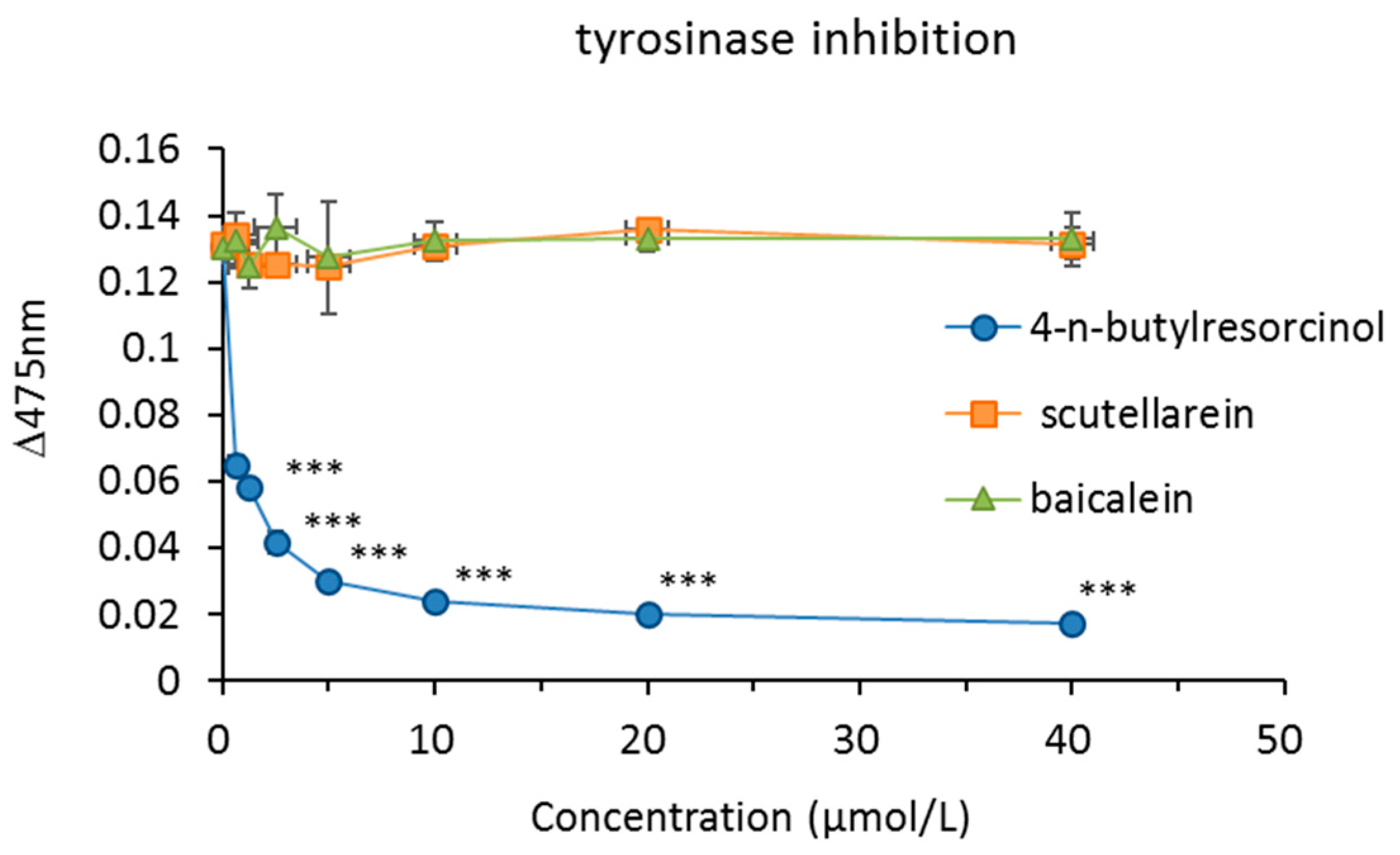
The inhibitory activity of scutellarein and baicalein on mushroom tyrosinase was investigated. 4-n-Butylresorcinol was used as a positive control. 4-n-Butylresorcinol showed a dose-dependent inhibitory effect, but scutellarein and baicalein did not inhibit the activity of mushroom tyrosinase (Figure 5).
3.5. Effects of Scutellarein on Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Protein Expressions
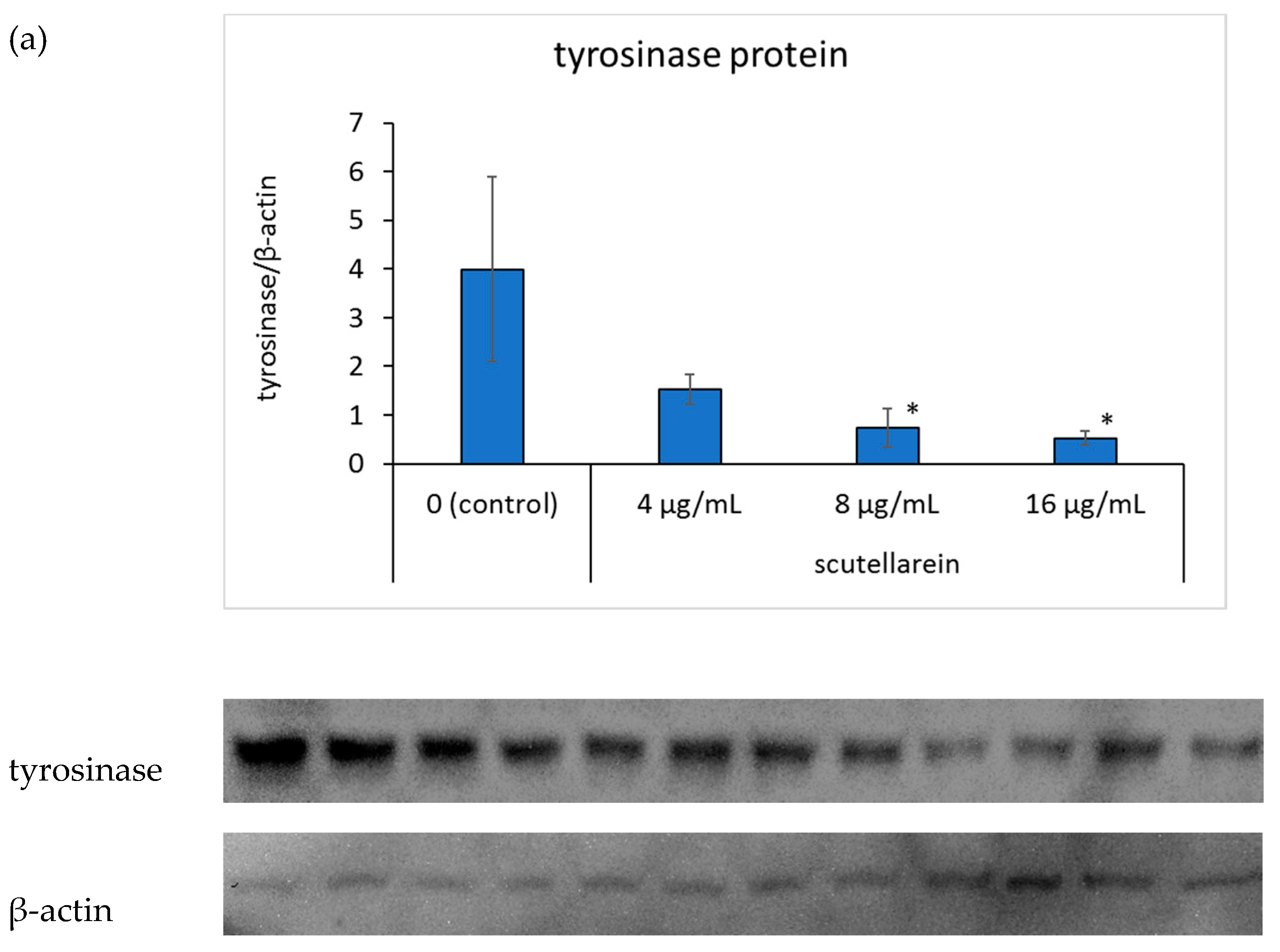
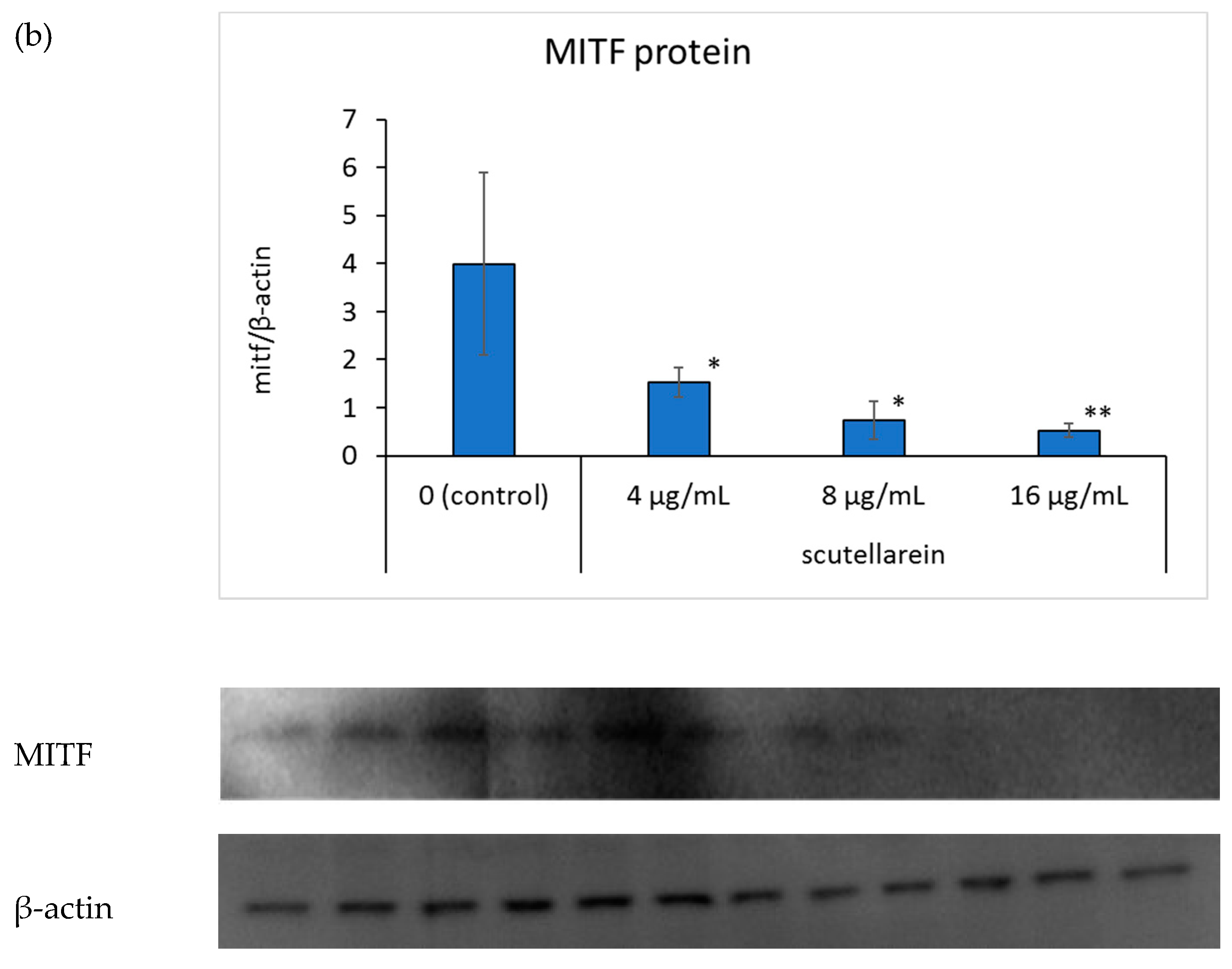
To further elucidate the mechanism of scutellarein in melanogenesis, we focused on tyrosinase and microphthalmia-associated transcription factor (MITF) protein expressions. Western blot analysis, using a specific antibody against tyrosinase, suggested that tyrosinase protein expression was inhibited in a dose-dependent manner when scutellarein concentration in the medium was 8 and 16 μg/mL (p < 0.05, p <0.05, respectively; Figure 6a).Additionally, the expression of MITF also showed in a dose-dependent manner when scutellarein concentration in the medium was 4, 8, and 16 μg/mL (p < 0.05, p < 0.05, p < 0.01, respectively; Figure 6b).
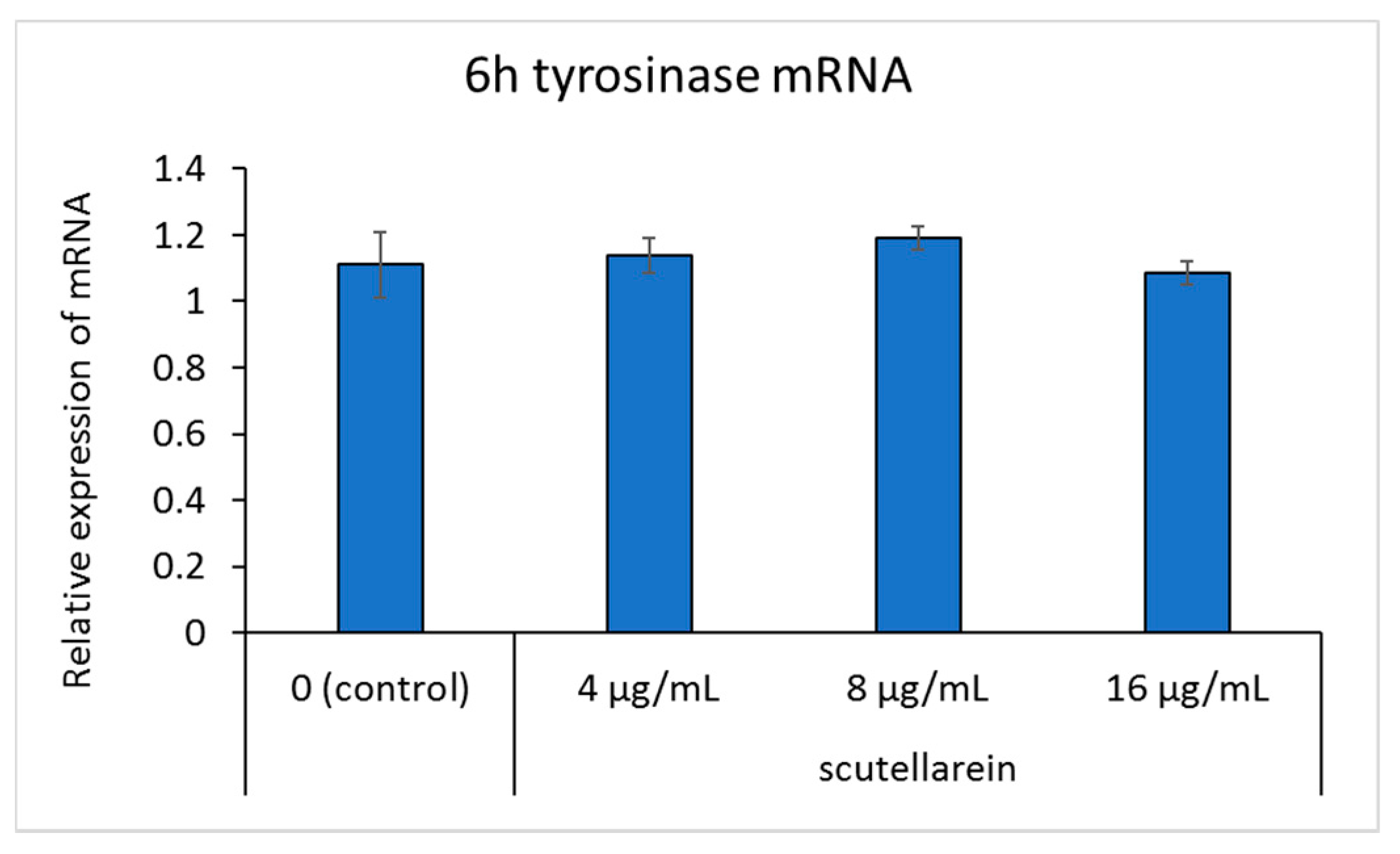
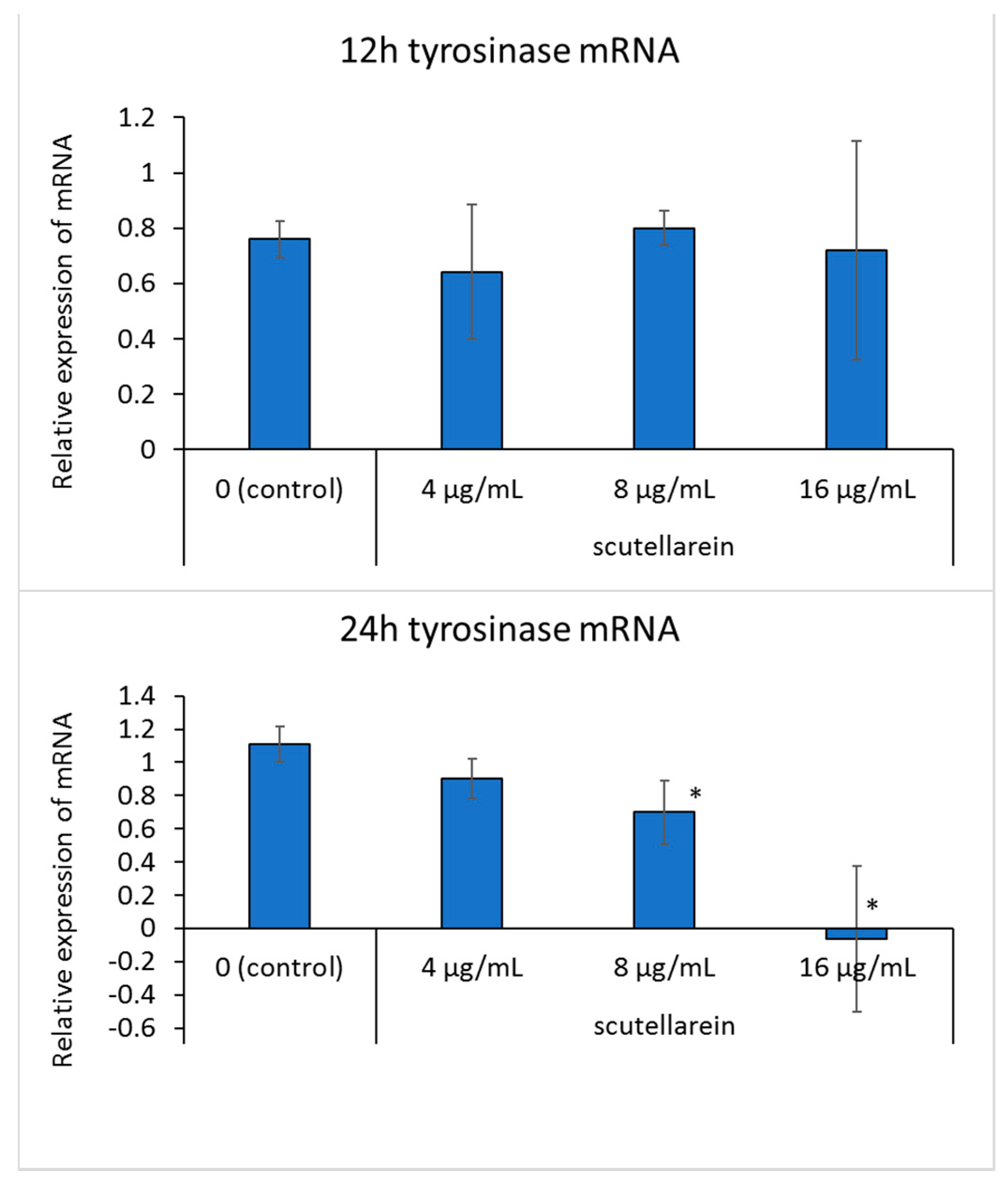
3.6. Effect of Scutellarein on Tyrosinase mRNA Expression
After adding scutellarein for 6, 12, and 24 h, tyrosinase mRNA expression in B16 cells was detected. However, quantitative analysis showed that after 6 and 12 h, tyrosinase mRNA expression in each concentration condition did not change significantly. After 24 h, tyrosinase mRNA levels were significantly downregulated at a concentration of 8 and 16 μg/mL (p < 0.05) in B16 cells, compared to the untreated control (Figure 7).
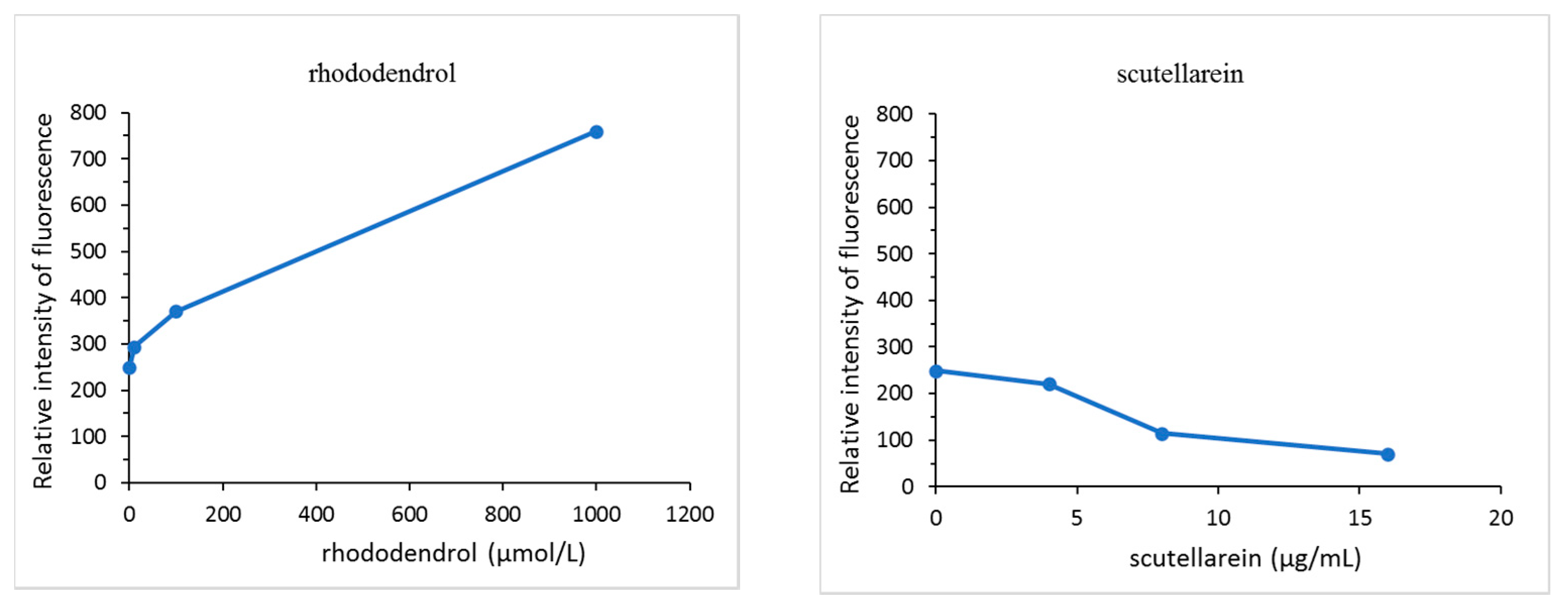
3.7. Hydroxyl Radical (˙OH) Generation from Scutellarein in the Presence of Tyrosinase
A comparison of the fluorescence intensity of the tested compounds in the presence of tyrosinase after 30 min is shown in Figure 8. The amount of free hydroxyl radicals (˙OH) generated did not change in the absence of tyrosinase. Compared to the control group, rhododendrol was found to generate ˙OH at 10 μmol/L (1.66 μg/mL), 100 μmol/L (16.6 μg/mL), and 1000 μmol/L (166 μg/mL). To contrast, the generation of ˙OH at each concentration of scutellarein was significantly lower than the control level at 4 μg/mL (14 μmol/L), 8 μg/mL (28 μmol/L), and 16 μg/mL (56 μmol/L) and, as the concentration increased, the levels of ˙OH decreased.
4. Discussion
Since the molecular structure of melanin facilitates ultraviolet light absorption, it has the ability to resist ultraviolet light [26]. Melanocytes are generally distributed in the hair, eyes, and skin. Since the 21st century, researchers have discovered many whitening agents used in drugs for treating pigment diseases or in skin-whitening cosmetics. Most whitening agents may be classified based on their different mechanisms of action, including the interference with melanin synthesis and transport and the acceleration of melanin decomposition [27]. Additionally, using a novel 675 nm laser may be considered promising when treating benign pigmented lesions, due to a low risk of side effects and simple post-treatment management [28].
Scutellarein and baicalein are flavones widely found in perennial herbs [29]. Presently, many scientists have confirmed that scutellarein has high anticancer activity and can be used as a drug for clinical treatment [30]. This study used three experiments assessing melanin content to explore the effects of scutellarein and baicalein on melanin production. The melanin content of B16 cells treated with scutellarein was significantly reduced after three days of culture; however, there was no change in the melanin content following baicalein treatment. We observed that scutellarein had an inhibitory effect on tyrosinase activity and showed no cytotoxicity at concentrations of 2 μg/mL, 4 μg/mL, 8 μg/mL, and 16 μg/mL. No enzyme inhibitory effect on tyrosinase was observed in scutellarein and baicalein. Therefore, in subsequent experiments, baicalein was not used as the tested compound. The chemical structures of scutellarein and baicalein were compared, and it was found that scutellarein has a 4-hydroxyphenyl in the 2-position, which has a stronger reducibility than baicalein, which has a phenyl group at the 2-position. This may explain why baicalein did not show any melanin inhibitory effect. The 4-substituted phenolic (para-phenol) compounds were previously shown to be directly toxic to melanocytes in the presence of tyrosinase [31]. Measuring the amount of free hydroxyl radicals (˙OH) generated in the presence of tyrosinase is an in vitro method that predicts the risk of the development of chemical leukoderma due to a particular compound [10]. Compared to the positive control, that is rhododendrol, scutellarein showed no ˙OH generation activity. Moreover, as the concentration of scutellarein increased, the ˙OH generation activity showed a downward trend. Scutellarein has been reported to have good antioxidant and anti-inflammatory effects, which may be one of the reasons for the inhibition of ˙OH generation. 4-n-Butylresorcinol did not show ˙OH generation activity in the presence of tyrosinase as previously reported [10].
Additionally, the effect of scutellarein on tyrosinase and microphthalmia-associated transcription factor (MITF) expressions also were tested. When scutellarein concentrations were 8 μg/mL and 16 μg/mL, tyrosinase protein levels decreased, and MITF protein levels decreased at the concentrations of 4 μg/mL, 8 μg/mL, and 16 μg/mL. Concurrently, tyrosinase mRNA levels decreased in a dose-dependent manner in the culture medium at concentrations of 8 μg/mL and 16 μg/mL. These results show that scutellarein affects melanin synthesis by inhibiting the expression of MITF and its down regulation tyrosinase protein. Conversely, 4-n-butylresorcinol has been reported to suppress melanin production by a mechanism that directly inhibits tyrosinase [8]. These results show that scutellarein affects melanin synthesis by inhibiting tyrosinase expression. However, in terms of gene expression, other factors that affect melanin production require further research. Furthermore, the effect of scutellarein on the synthesis and decomposition of melanosomes via different pathways is still being investigated by our group.
To conclude, our results revealed that scutellarein has an inhibitory effect on melanin production at the concentration of any cytotoxicity; inhibition of ˙OH generation also was observed. While investigating the mechanism of scutellarein’s inhibition of melanin production, we found that tyrosinase expression was inhibited, which ultimately affected the synthesis of melanin in cells. To explore whether scutellarein could serve as a promising new skin-lightening agent, further experiments and research, such as the cutaneous absorption experiment, concentration setting experiment, and clinical trials, are needed.
Author Contributions
L.D., L.G. and K.M. performed the experiments. K.M. designed the study and performed the data analysis. L.D., L.G. and K.M. interpreted the data and drafted the manuscript. K.M. supervised the study and critically revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nimitphong, H.; Holick, M.F. Vitamin D status and sun exposure in southeast Asia. Dermato-Endocrinology 2013, 5, 34–37. [Google Scholar] [CrossRef] [Green Version]
- Li, E.P.H.; Min, H.J.; Belk, R.W. Skin Lightening and Beauty in Four Asian Cultures. In NA-Adv. in Consumer Research; Lee, A.Y., Soman, D., Eds.; Association for Consumer Research: Duluth, MN, USA, 2008; Volume 35, pp. 444–449. [Google Scholar]
- Fuji-Keizai. Kinouseikeshouhin Marketing Youran 2019–2020 (Functional Cosmetics Marketing Handbook 2019–2020); Fuji-Keizai: Tokyo, Japan, 2019; pp. 64–80. (In Japanese) [Google Scholar]
- Homepage of Kanebo Cosmetics, Inc. The Number of Checks of the White spots’ Condition, and Recovery, a Reconciliation Situation/the Number of Object Recall. Available online: https://www.kanebo-cosmetics.jp/information/correspondence/data_2016.html (accessed on 12 February 2021). (In Japanese).
- Maeda, K.; Fukuda, M. Arbutin: Mechanism of its depigmenting action in human melanocyte culture. J. Pharmacol. Exp. Ther. 1996, 276, 765–769. [Google Scholar]
- Maeda, K.; Naganuma, M. Topical trans-4-aminomethylcyclohexanecarboxylic acid prevents ultraviolet radiation-induced pigmentation. J. Photochem. Photobiol. B 1998, 47, 136–141. [Google Scholar] [CrossRef]
- Maeda, K.; Inoue, Y.; Nishikawa, H.; Miki, S.; Urushibata, O.; Miki, T.; Hatao, M. Involvement of melanin monomers in the skin persistent UVA-pigmentation and effectiveness of vitamin C ethyl on UVA-pigmentation. J. Jpn. Cosmet. Sci. Soc. 2003, 27, 257–268. (In Japanese) [Google Scholar]
- Kim, D.S.; Kim, S.Y.; Park, S.H.; Choi, Y.G.; Kwon, S.B.; Kim, M.K.; Na, J.I.; Youn, S.W.; Park, K.C. Inhibitory effects of 4-n-butylresorcinol on tyrosinase activity and melanin synthesis. Biol. Pharm. Bull. 2005, 28, 2216–2219. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K. Advances in development of skin whitening agents. Fragr. J. 2008, 36, 65–67. (In Japanese) [Google Scholar]
- Gu, L.; Zeng, H.; Takahashi, T.; Maeda, K. In vitro methods for predicting chemical leukoderma caused by quasi-drug cosmetics. Cosmetics 2017, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Ohtsuji, H.; Miyahara, S. Two cases of leucoderma, presumably due to nonyl or octylphenol in synthetic detergents. Ind. Health (Kawasaki) 1970, 8, 192–196. [Google Scholar] [CrossRef]
- Xu, P.; Su, S.; Tan, C.; Lai, R.-S.; Min, Z.-S. Effects of aqueous extracts of Ecliptae herba, Polygoni multiflori radix praeparata and Rehmanniae radix praeparata on melanogenesis and the migration of human melanocytes. J. Ethnopharmacol. 2017, 195, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Lv, N.; Koo, J.-H.; Yoon, H.-Y.; Yu, J.; Kim, K.-A.; Choi, I.-W.; Kwon, K.-B.; Kwon, K.-S.; Kim, H.-U.; Park, J.-W.; et al. Effect of Angelica gigas extract on melanogenesis in B16 melanoma cells. Int. J. Mol. Med. 2007, 20, 763–767. [Google Scholar] [PubMed] [Green Version]
- Choi, M.Y.; Song, H.S.; Hur, H.S.; Sim, S.S. Whitening activity of luteolin related to the inhibition of cAMP pathway in α-MSH-stimulated B16 melanoma cells. Arch. Pharm. Res. 2008, 31, 1166–1171. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Downey, F.; Ng, C.K.Y. Comparative analysis of bioactive phytochemicalsfrom Scutellaria baicalensis, Scutellaria lateriflora, Scutellariaracemosa, Scutellaria tomentosa and Scutellaria wrightii by LC-DAD-MS. Metabolomics 2011, 7, 446–453. [Google Scholar] [CrossRef]
- Han, J.; Ye, M.; Xu, M.; Sun, J.; Wang, B.; Guo, D. Characterization of flavonoids in the traditional Chinese herbal medicine-Huangqin by liquid chromatography coupled with electrospray ionization mass spectrometry. J. Chromatogr. B 2007, 848, 355–362. [Google Scholar] [CrossRef]
- Sadasivam, K.; Kumaresan, R. A comparative DFT study on the antioxidant activity of apigenin and scutellarein flavonoid compounds. Mol. Phys. 2011, 109, 839–852. [Google Scholar] [CrossRef]
- Sung, N.Y.; Kim, M.-Y.; Cho, J.Y. Scutellarein reduces inflammatory responses by inhibiting Src kinase activity. Korean J. Physiol. Pharmacol. 2015, 19, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Hamada, H.; Hiramatsu, M.; Edamatsu, R.; Mori, A. Free radical scavenging action of baicalein. Arch. Biochem. Biophys. 1993, 306, 261–266. [Google Scholar] [CrossRef]
- Hong, S.J.; Kim, K.J. Scutellaria baicalensis Georgi (SBG) inhibits melanin synthesis in mouse B16 melanoma cells. J. Korean Orient. Med. Ophthalmol. Otolaryngol. Dermatol. 2009, 22, 104–117. [Google Scholar]
- Maeda, K. Large melanosome complexis increased in keratinocytes of solar lentigo. Cosmetics 2017, 4, 49. [Google Scholar]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- Makpol, S.; Jam, F.A.; Rahim, N.A.; Khor, S.C.; Ismail, Z.; Yusof, Y.A.M.; Wan Ngah, W.Z. Comparable down-regulation of TYR, TYRP1 and TYRP2 genes and inhibition of melanogenesis by tyrostat, tocotrienol-rich fraction and tocopherol in human skin melanocytes improves skin pigmentation. Clin. Ter. 2014, 165, 39–45. [Google Scholar]
- D’Mello, S.; Finlay, G.J.; Baguley, B.C.; Askarian, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar]
- Stanojević, M.R.; Stanojević, Z.; Jovanović, D.L. Ultraviolet radiation and melanogenesis. Arch. Oncol. 2004, 12, 203–205. [Google Scholar] [CrossRef]
- Smit, N.; Vicanova, J.; Pavel, S. The hunt for natural skin whitening agents. Int. J. Mol. Sci. 2009, 10, 5326–5349. [Google Scholar] [CrossRef] [PubMed]
- Nisticò, S.P.; Tolone, M.; Zingoni, T.; Tamburi, F.; Scali, E.; Bennardo, L.; Cannarozzo, G. A new 675 nm laser device in the treatment of melasma:results of a prospective observational study. Photobiomod. Photomed. Laser Surg. 2020, 38, 560–564. [Google Scholar] [CrossRef]
- Li, J.; Tian, C.; Xia, Y.; Mutanda, I.; Wang, K.; Wang, Y. Production of plant-specific flavones baicalein and scutellarein in an engineered E. coli from available phenylalanine and tyrosine. Metab. Eng. 2019, 52, 124–133. [Google Scholar] [PubMed]
- Shi, X.; Chen, G.; Liu, X.; Qiu, Y.; Yang, S.; Zhang, Y.; Fang, X.; Zhang, C.; Liu, X. Scutellarein inhibits cancer cell metastasis in vitro and attenuates the development of fibrosarcoma in vivo. Int. J. Mol. Med. 2015, 35, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, N.P.; Peters, K.; Menko, W.; Westerhof, W.; Pavel, S.; Riley, P.A. Cytotoxicity of a selected series of substituted phenols towards cultured melanoma cells. Melanoma Res. 1992, 2, 295–304. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The chemical structure of 4-n-butylresorcinol, scutellarein [5,6,7-trihydroxy-2-(4-hydroxyphenyl)-chromen-4-one] and baicalein (5,6,7-trihydroxy-2-phenyl-chromen-4-one) derived from Scutellaria baicalensis Georgi (SBG).
Figure 1.
The chemical structure of 4-n-butylresorcinol, scutellarein [5,6,7-trihydroxy-2-(4-hydroxyphenyl)-chromen-4-one] and baicalein (5,6,7-trihydroxy-2-phenyl-chromen-4-one) derived from Scutellaria baicalensis Georgi (SBG).

Figure 2.
Effects of the tested compounds on the melanin content of B16 cells. The upper part of the figure shows the appearance of the cell suspension after the addition of the NaOH solution. The lower part of the figure represents the absorbance of the cell pellet dissolved in NaOH; absorbance was measured at 475 nm. Data are expressed as a percentage of the control. (a): 4-n-butylresorcinol; (b): scutellarein; (c): baicalein.
Figure 2.
Effects of the tested compounds on the melanin content of B16 cells. The upper part of the figure shows the appearance of the cell suspension after the addition of the NaOH solution. The lower part of the figure represents the absorbance of the cell pellet dissolved in NaOH; absorbance was measured at 475 nm. Data are expressed as a percentage of the control. (a): 4-n-butylresorcinol; (b): scutellarein; (c): baicalein.

Figure 3.
The effects of the tested compounds on the tyrosinase activity of B16 cells. Tyrosinase activity was determined by calculating the absorbance change rate in 2 h at 475 nm. (a): 4-n-butylresorcinol; (b): scutellarein. Data are presented as the mean ± SD of experiments performed in triplicate. ∗ p < 0.05 compared to the control, ∗∗ p < 0.01 compared to the control.
Figure 3.
The effects of the tested compounds on the tyrosinase activity of B16 cells. Tyrosinase activity was determined by calculating the absorbance change rate in 2 h at 475 nm. (a): 4-n-butylresorcinol; (b): scutellarein. Data are presented as the mean ± SD of experiments performed in triplicate. ∗ p < 0.05 compared to the control, ∗∗ p < 0.01 compared to the control.


Figure 4.
The effects of the tested compounds on the viability of B16 cells. Cell viability was determined by calculating the absorbance change rate in 2 h at 450 nm. (a): 4-n-butylresorcinol; (b): scutellarein. Data are presented as the mean ± SD of experiments performed in triplicate.
Figure 4.
The effects of the tested compounds on the viability of B16 cells. Cell viability was determined by calculating the absorbance change rate in 2 h at 450 nm. (a): 4-n-butylresorcinol; (b): scutellarein. Data are presented as the mean ± SD of experiments performed in triplicate.

Figure 5.
The effects of the tested compounds on mushroom tyrosinase activity. Mushroom tyrosinase activity was determined by calculating the absorbance change rate in 10 min at 475 nm. 4-n-butylresorcinol: 40 μmol/L (6.64 μg/mL), 20 μmol/L (3.32 μg/mL), 10 μmol/L (1.66 μg/mL), 5 μmol/L (0.83 μg/mL), 2.5 μmol/L (0.42 μg/mL), 1.25 μmol/L (0.21 μg/mL), 0.625 μmol/L (0.10 μg/mL). scutellarein: 40 μmol/L (11.44 μg/mL), 20 μmol/L (5.72 μg/mL), 10 μmol/L (2.86 μg/mL), 5 μmol/L (1.43 μg/mL), 2.5 μmol/L (0.72 μg/mL), 1.25 μmol/L (0.36 μg/mL), 0.625 μmol/L (0.18 μg/mL). baicalein: 40 μmol/L (10.8 μg/mL), 20 μmol/L (5.4 μg/mL), 10 μmol/L (2.7 μg/mL), 5 μmol/L (1.35 μg/mL), 2.5 μmol/L (0.68 μg/mL), 1.25 μmol/L (0.34 μg/mL), 0.625 μmol/L (0.17 μg/mL). * p < 0.05, ** p < 0.01, *** p < 0.001 compared to the control. Data are presented as the mean ± SD of experiments performed in triplicate.
Figure 5.
The effects of the tested compounds on mushroom tyrosinase activity. Mushroom tyrosinase activity was determined by calculating the absorbance change rate in 10 min at 475 nm. 4-n-butylresorcinol: 40 μmol/L (6.64 μg/mL), 20 μmol/L (3.32 μg/mL), 10 μmol/L (1.66 μg/mL), 5 μmol/L (0.83 μg/mL), 2.5 μmol/L (0.42 μg/mL), 1.25 μmol/L (0.21 μg/mL), 0.625 μmol/L (0.10 μg/mL). scutellarein: 40 μmol/L (11.44 μg/mL), 20 μmol/L (5.72 μg/mL), 10 μmol/L (2.86 μg/mL), 5 μmol/L (1.43 μg/mL), 2.5 μmol/L (0.72 μg/mL), 1.25 μmol/L (0.36 μg/mL), 0.625 μmol/L (0.18 μg/mL). baicalein: 40 μmol/L (10.8 μg/mL), 20 μmol/L (5.4 μg/mL), 10 μmol/L (2.7 μg/mL), 5 μmol/L (1.35 μg/mL), 2.5 μmol/L (0.68 μg/mL), 1.25 μmol/L (0.34 μg/mL), 0.625 μmol/L (0.17 μg/mL). * p < 0.05, ** p < 0.01, *** p < 0.001 compared to the control. Data are presented as the mean ± SD of experiments performed in triplicate.

Figure 6.
The effects of scutellarein on tyrosinase protein (a) and microphthalmia-associated transcription factor (MITF) protein (b) expressions. B16 cells were treated with different concentrations of scutellarein for 48 h. Western blot analysis was then conducted using an anti-tyrosinase antibody or anti-MITF antibody; β-actin was used as a loading control. Fluorescence intensities of tyrosinase and β-actin were quantified using ImageJ. Band intensities were normalized to β-actin. Data are presented as the mean ± SD of experiments performed in triplicate. * p < 0.05 compared to the control. ** p < 0.01 compared to the control.
Figure 6.
The effects of scutellarein on tyrosinase protein (a) and microphthalmia-associated transcription factor (MITF) protein (b) expressions. B16 cells were treated with different concentrations of scutellarein for 48 h. Western blot analysis was then conducted using an anti-tyrosinase antibody or anti-MITF antibody; β-actin was used as a loading control. Fluorescence intensities of tyrosinase and β-actin were quantified using ImageJ. Band intensities were normalized to β-actin. Data are presented as the mean ± SD of experiments performed in triplicate. * p < 0.05 compared to the control. ** p < 0.01 compared to the control.


Figure 7.
The effects of scutellarein on tyrosinase mRNA expression. B16 cells were treated with different concentrations of scutellarein for 6, 12, and 24 h. RT-qPCR analysis was then performed using the glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) as a loading control. Data are presented as the mean ± SD of experiments performed in triplicate. * p < 0.05 compared to the control.
Figure 7.
The effects of scutellarein on tyrosinase mRNA expression. B16 cells were treated with different concentrations of scutellarein for 6, 12, and 24 h. RT-qPCR analysis was then performed using the glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) as a loading control. Data are presented as the mean ± SD of experiments performed in triplicate. * p < 0.05 compared to the control.


Figure 8.
A comparison of free hydroxyl radical (˙OH) generation by the tested compound in the presence of tyrosinase.
Figure 8.
A comparison of free hydroxyl radical (˙OH) generation by the tested compound in the presence of tyrosinase.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dai, L.; Gu, L.; Maeda, K. Inhibitory Effect and Mechanism of Scutellarein on Melanogenesis. Cosmetics 2021, 8, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010015
AMA Style
Dai L, Gu L, Maeda K. Inhibitory Effect and Mechanism of Scutellarein on Melanogenesis. Cosmetics. 2021; 8(1):15. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010015
Chicago/Turabian StyleDai, Liyun, Lihao Gu, and Kazuhisa Maeda. 2021. "Inhibitory Effect and Mechanism of Scutellarein on Melanogenesis" Cosmetics 8, no. 1: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics8010015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.