

Cell-Based Model Systems for Validation of Various Efficacy-Based Claims for Cosmetic Ingredients
 ,
,
Abstract
:1. Introduction
2. Cell-Based Strategies for Cosmetic Ingredients
2.1. Critical Parameters for In Vitro and Cell-Based Assays
- The color and turbidity of test substances can interfere with assay signals (e.g., absorbance and fluorescence).
- In many cases, the test compounds can exhibit metal ion chelation and redox cycling properties.
- The test substance can form protein aggregates or exhibit non-specific protein reactivity, which can be misinterpreted as mechanism-specific inhibition [10].
- Anti-acne activity
- Hair-growth-promoting activity
- Anti-aging and skin-rejuvenating activity
- Anti-psoriatic activity
- Wound-healing activity
- Anti-skin-hyperpigmentation activity
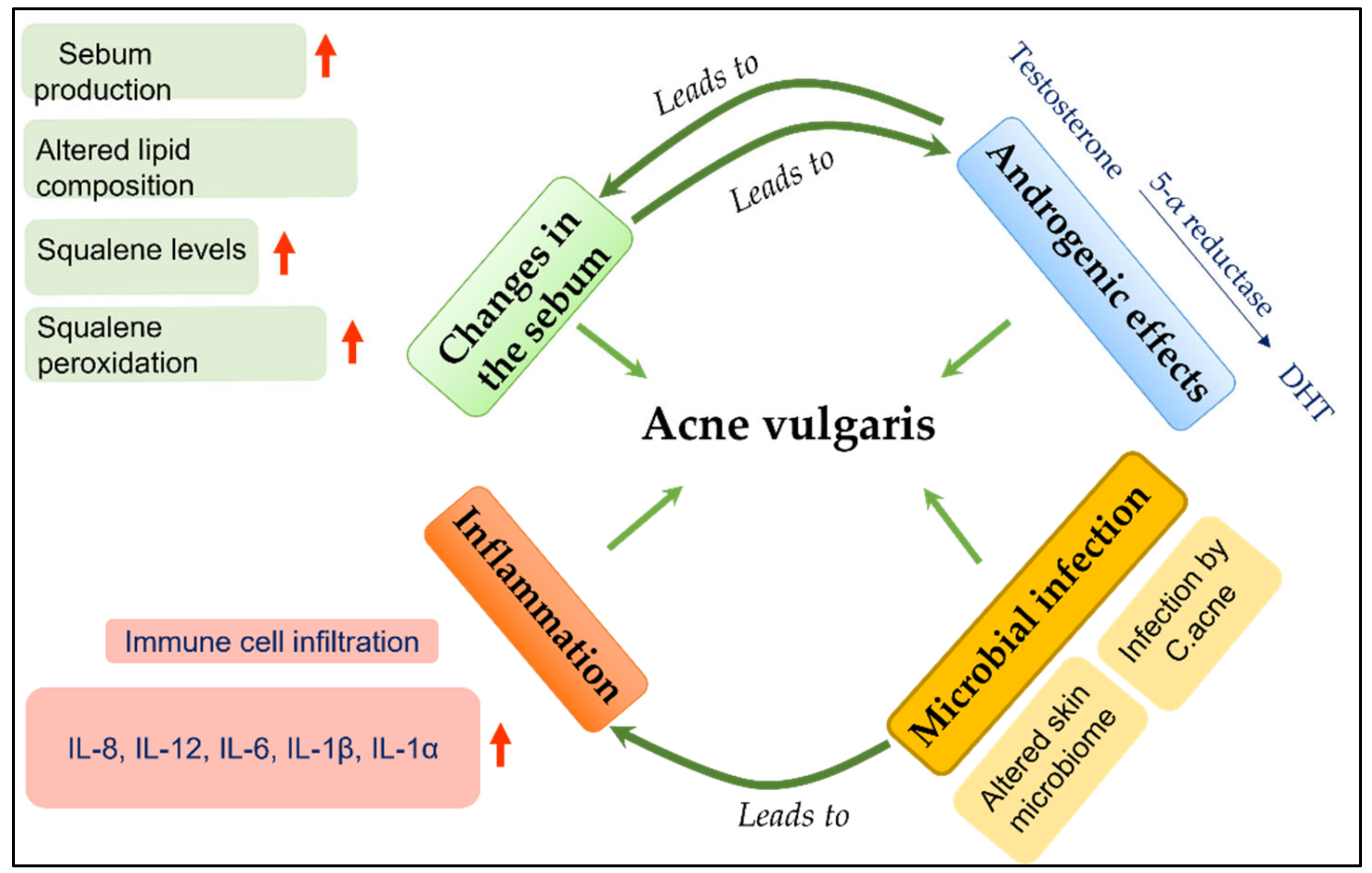
2.2. Evaluation of Effect of Cosmeticingredients on Acne Vulgaris
2.2.1. Evaluating the Anti-Inflammatory Activity of Cosmetic Ingredients against C. acnes-Induced Inflammation
2.2.2. Evaluating the Effect of Cosmetic Ingredients on Sebum Secretion
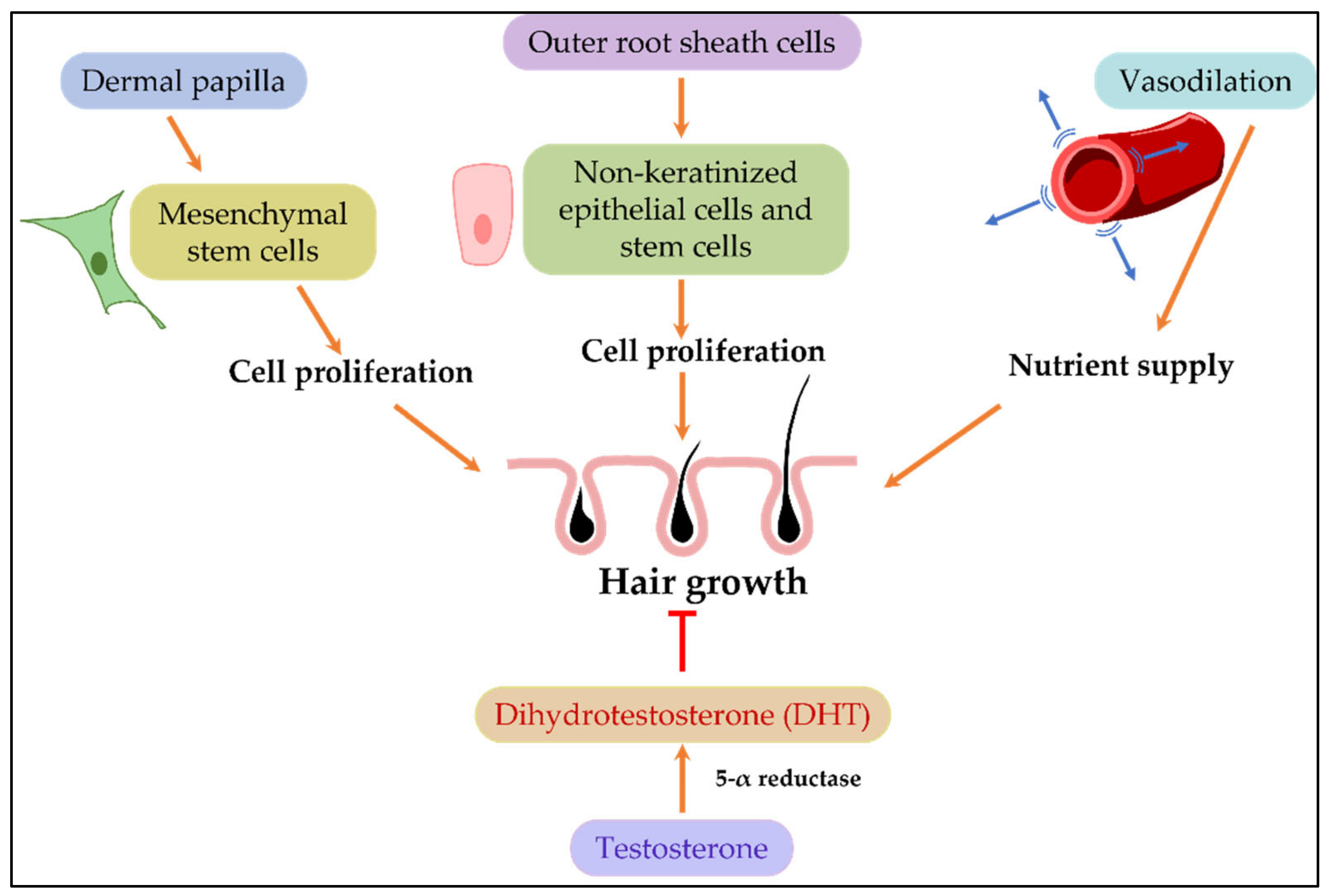
2.3. Evaluation of the Effect of Cosmetic Ingredients on Hair Growth
2.3.1. Assay Systems Based on Proliferation of the Hair Follicle Cells
2.3.2. 5-α Reductase Inhibition
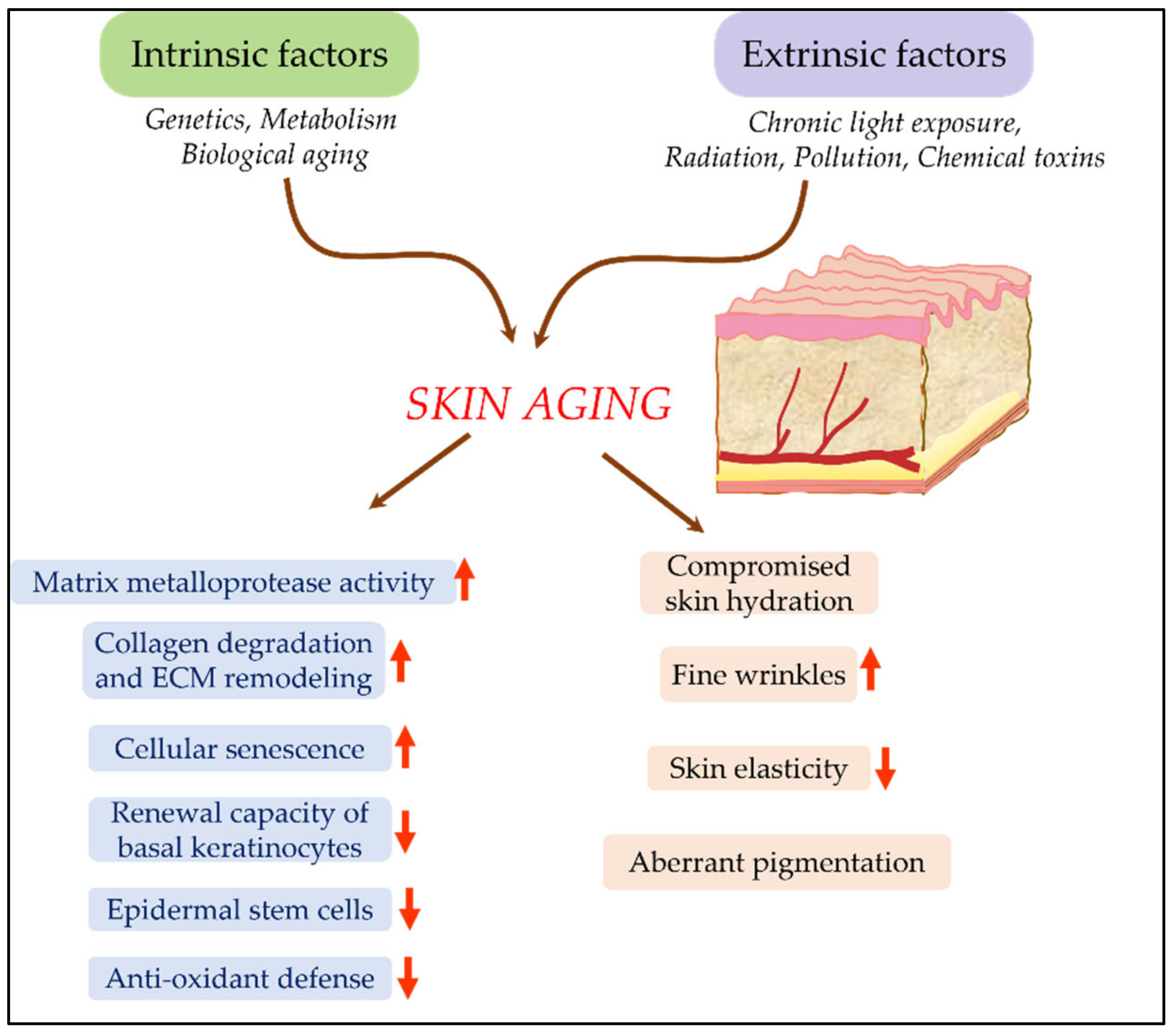
2.4. Evaluation of Effect of Cosmetic Ingredients on Skin-Aging and Rejuvenation
2.4.1. Skin Aging Models Based on Extra-Cellular Matrix (ECM) Dynamics
2.4.2. Measurement of Hyaluronic Acid (HA) Levels
2.4.3. Cellular Senescence Model for Skin Aging
2.5. Gene Expression Studies to Evaluate Skin Hydration, Skin Barrier Function and Skin Rejuvenation
2.6. Evaluation of the Effect of Cosmetic Ingredients on Psoriasis
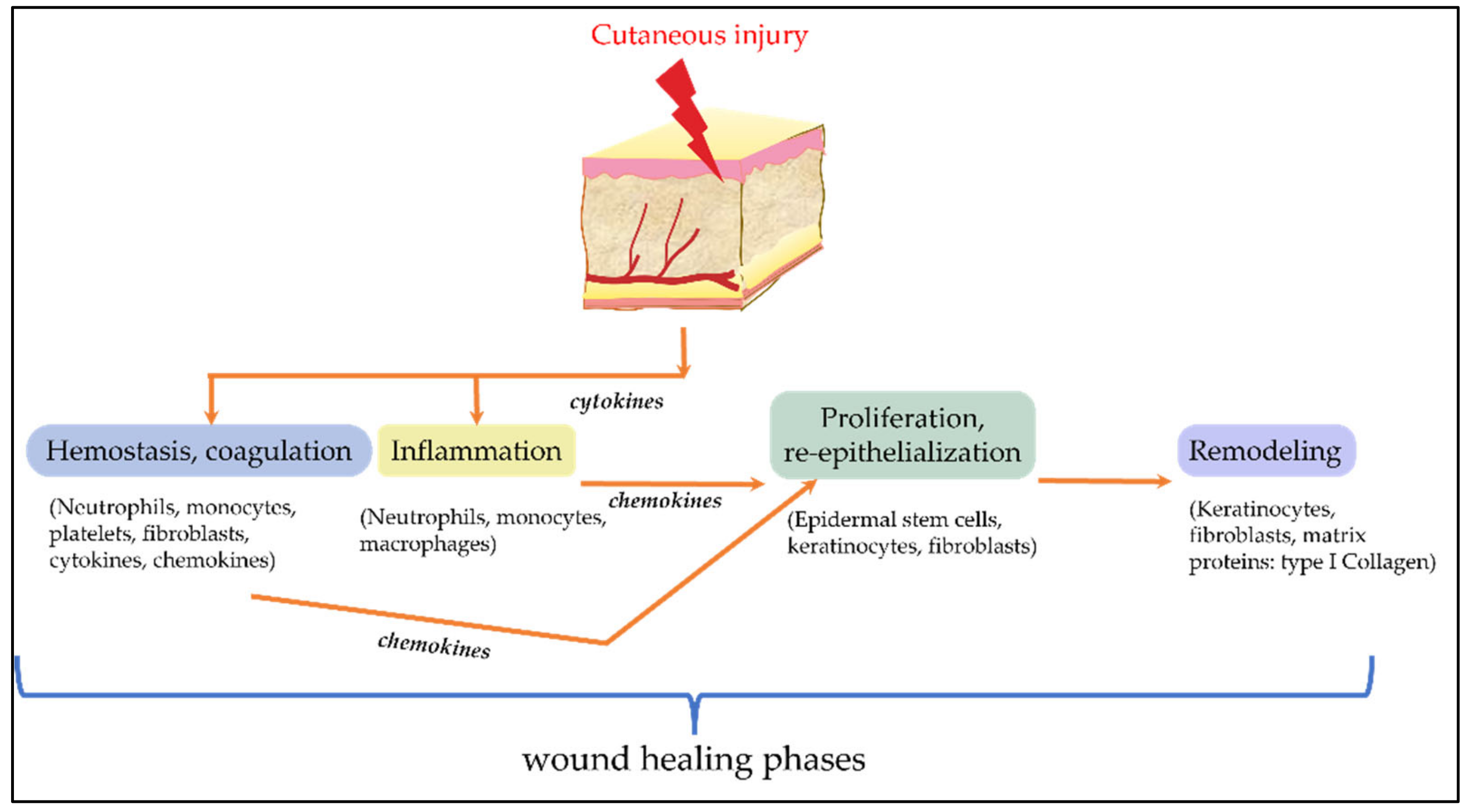
2.7. Cell-Based Systems for Evaluation of Cosmetic Ingredients on Wound-Healing
2.8. Cell-Based Systems for Skin Hyperpigmentation
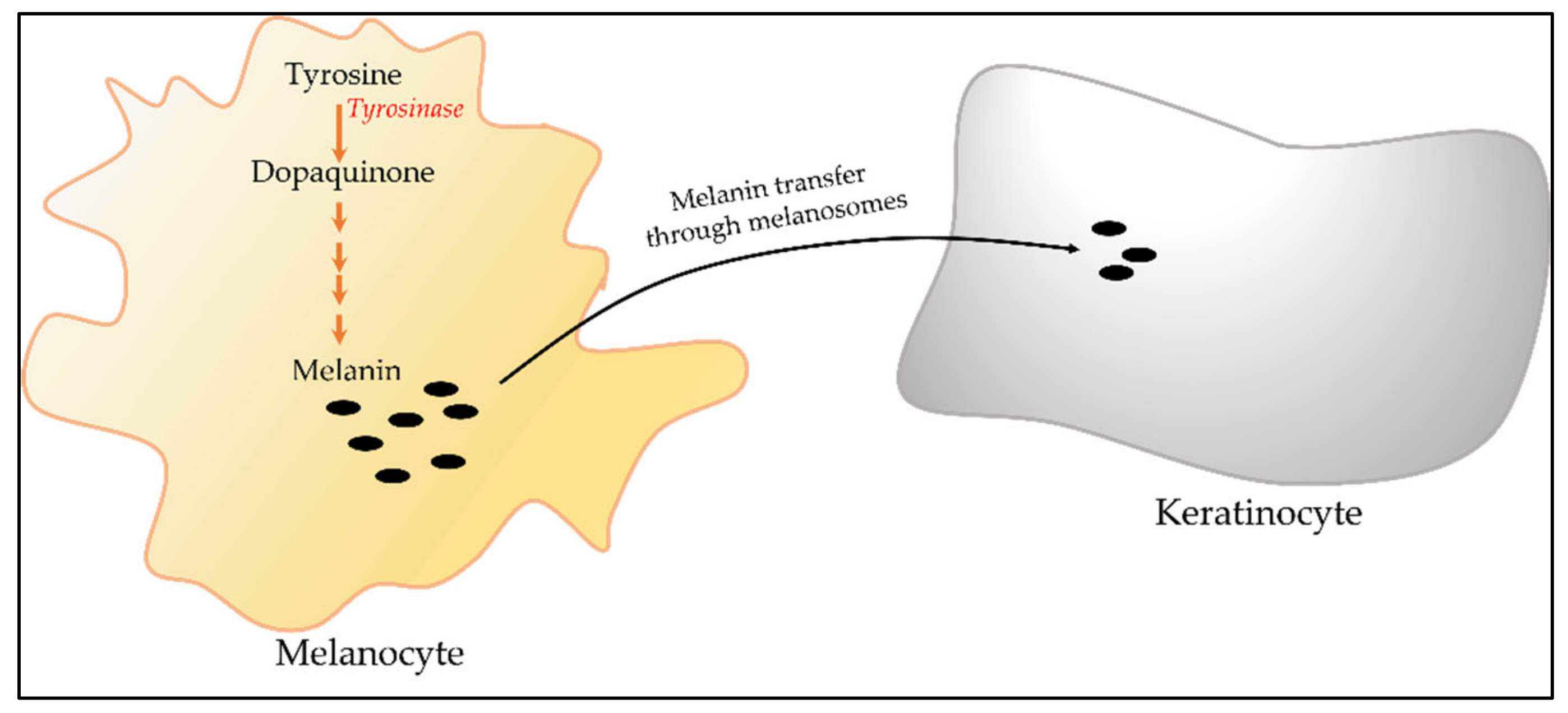
2.8.1. Cell-Based Assay for Melanogenesis
2.8.2. Tyrosinase Inhibition Assay
2.9. Melanosome Transfer Assay
3. Challenges and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| AMP | anti-microbial peptide |
| AQP3 | aquaporin-3 |
| C. acne | Cutibacterium acnes |
| CFDA | carboxyfluorescein diacetate |
| COL1A | collagen |
| DHT | dihydrotestosterone |
| EC | European Commission |
| ECM | extra-cellular matrix |
| ELISA | enzyme-linked immunoassay |
| ELN | elastin |
| EPSCs | epidermal stem cells |
| ERK | extracellular signal-regulated kinases |
| FDA | Food and Drug Administration |
| FGF | fibroblast growth factor |
| FLG | filaggrin |
| HA | hylauronic acid |
| Hacat | human epidermal keratinocytes |
| HDF | human dermal fibroblast |
| HFDPC | human hair follicle dermal papilla cells |
| HGF | hepatocyte growth factor |
| HPLC | high-performance liquid chromatography |
| IC50 | inhibitory concentration 50 |
| IGF-1 | insulin growth factor 1 |
| IL | interleukins |
| INV | involucrin |
| KGF | keratinocyte growth factor |
| LC-MS | liquid chromatography–mass spectrometry |
| L-DOPA | l-3,4-dihydroxyphenylalanine |
| MEF | mouse primary embryonic fibroblasts |
| MFAP4 | microfibril-associated glycoprotein 4 |
| MMPs | matrix metalloprotein |
| MTT/XTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OECD | Organisation for Economic Co-operation and Development |
| ORSc | outer root sheath cells |
| PCR | polymerase chain reaction |
| PDGF-C | platelet-derived growth factor C |
| PTU | phenyl thiourea |
| ROS | reactive oxygen species |
| SA-β-gal | senescence-associated beta-galactosidase |
| TGF-β1 | transforming growth factor beta |
| TGMs | transglutaminase 1, 3, and 5 |
| TIMP1 | tissue inhibitors of metalloproteinases |
| TLC | thin-layer chromatography |
| TNF-alpha | tumor necrosis factor alpha |
| UV | ultraviolet |
| VEGF | vascular endothelial growth factor |
| α-MSH | alpha melanocyte-stimulating hormones |
References
- Cosmetics. Available online: https://cdsco.gov.in/opencms/opencms/en/Cosmetics/cosmetics/ (accessed on 8 September 2022).
- Importing Cosmetics|FDA. Available online: https://www.fda.gov/industry/importing-fda-regulated-products/importing-cosmetics#cosmetic (accessed on 8 September 2022).
- Regulation (EC) No 1223/2009 of the European Parliament and of the Council of 30 November 2009 On cosmetic products (recast) (Text with EEA relevance). Off. J. Eur. Union. 2009. Available online: https://ec.europa.eu/health/sites/health/files/endocrine_disruptors/docs/cosmetic_1223_2009_regulation_en.pdf (accessed on 8 September 2022).
- Chanchal, D.; Swarnlata, S. Novel approaches in herbal cosmetics. J. Cosmet. Derm. 2008, 7, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Jatana, G.K.; Sonthalia, S. Cosmeceuticals. Adv. Integr. Dermatol. 2019, 393–411. [Google Scholar] [CrossRef]
- Brandt, F.S.; Cazzaniga, A.; Hann, M. Cosmeceuticals: Current trends and market analysis. Semin. Cutan. Med. Surg. 2011, 30, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Kligman, A. The future of cosmeceuticals: An interview with Albert Kligman, MD, PhD. Interview by Zoe Diana Draelos. Dermatol. Surg. 2005, 31, 890–891. [Google Scholar] [PubMed]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Baell, J.B.; Holloway, G.A. New substructure filters for removal of pan assay interference compounds (PAINS) from screening libraries and for their exclusion in bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [Green Version]
- Baell, J.B. Feeling Nature’s PAINS: Natural Products, Natural Product Drugs, and Pan Assay Interference Compounds (PAINS). J. Nat. Prod. 2016, 79, 616–628. [Google Scholar] [CrossRef]
- The Role of Inflammation in the Pathology of Acne. Available online: https://pubmed.ncbi.nlm.nih.gov/24062871/ (accessed on 28 July 2022).
- Dodou, K. Special Issue “Current and Evolving Practices in the Quality Control of Cosmetics”. Cosmetics 2021, 8, 100. [Google Scholar] [CrossRef]
- Dréno, B. What is new in the pathophysiology of acne, an overview. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Picardo, M.; Ottaviani, M.; Camera, E. Mastrofrancesco A. Sebaceous gland lipids. Dermatoendocrinology 2009, 1, 68. [Google Scholar] [CrossRef]
- Ottaviani, M.; Camera, E.; Picardo, M. Lipid Mediators in Acne. Mediat. Inflamm. 2010, 2010, 858176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandéz, J.R.; Rouzard, K.; Voronkov, M.; Feng, X.; Stock, J.B.; Stock, M.; Gordon, J.S.; Shroot, B.; Christensen, M.S.; Pérez, E. SIG1273: A new cosmetic functional ingredient to reduce blemishes and Propionibacterium acnes in acne prone skin. J. Cosmet. Dermatol. 2012, 11, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, J.B. Licorice (Glycyrrhiza glabra Linn) as a Valuable Medicinal Plant. Available online: https://www.researchgate.net/publication/268502890 (accessed on 29 July 2022).
- Joshi, M.D.; Damle, M. Glycyrrhiza glabra (Liquorice)-a potent medicinal herb. Int. J. Herb. Med. 2014, 2, 132–136. Available online: https://www.researchgate.net/publication/305465442 (accessed on 29 July 2022).
- Garcia-Jares, C.; Rubio, L.; Baby, A.R.; Freire, T.B.; De Argollo Marques, G.; Rijo, P.; Lima, F.V.; Carlos, J.; De Carvalho, M.; Rojas, J.; et al. Azadirachta indica (Neem) as a Potential Natural Active for Dermocosmetic and Topical Products: A Narrative Review. Cosmetics 2022, 9, 58. [Google Scholar] [CrossRef]
- Chainani-Wu, N. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcuma longa). J. Altern. Complement. Med. 2003, 9, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Park, W. Anti-Inflammatory Effect of Myristicin on RAW 264.7 Macrophages Stimulated with Polyinosinic-Polycytidylic Acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, P.; Besra, S.E.; Gomes, A.; Das, M.; Sur, P.; Mitra, S.; Vedasiromoni, J.R. Anti-inflammatory activity of tea (Camellia sinensis) root extract. Life Sci. 2004, 74, 1839–1849. [Google Scholar] [CrossRef]
- Yogesh, H.R.; Gajjar, T.; Patel, N.; Kumawat, R. Clinical study to assess efficacy and safety of Purifying Neem Face Wash in prevention and reduction of acne in healthy adults. J. Cosmet. Dermatol. 2021, 21, 2849–2858. [Google Scholar] [CrossRef]
- Okoro, O.E.; Adenle, A.; Ludovici, M.; Truglio, M.; Marini, F.; Camera, E. Lipidomics of facial sebum in the comparison between acne and non-acne adolescents with dark skin. Sci. Rep. 2021, 11, 16591. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Seltmann, H.; Neitzel, H.; Orfanos, C.E. Establishment and characterization of an immortalized human sebaceous gland cell line (SZ95). J. Investig. Dermatol. 1999, 113, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Lo Celso, C.; Berta, M.A.; Braun, K.M.; Frye, M.; Lyle, S.; Zouboulis, C.C.; Watt, F.M. Characterization of bipotential epidermal progenitors derived from human sebaceous gland: Contrasting roles of c-Myc and beta-catenin. Stem Cells 2008, 26, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Thiboutot, D.; Jabara, S.; McAllister, J.M.; Sivarajah, A.; Gilliland, K.; Cong, Z.; Clawson, G. Human skin is a steroidogenic tissue: Steroidogenic enzymes and cofactors are expressed in epidermis, normal sebocytes, and an immortalized sebocyte cell line (SEB-1). J. Investig. Dermatol. 2003, 120, 905–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrault, C.; Dichamp, I.; Garnier, J.; Pedretti, N.; Juchaux, F.; Deguercy, A.; Agius, G.; Bernard, F.X. Immortalized sebocytes can spontaneously differentiate into a sebaceous-like phenotype when cultured as a 3D epithelium. Exp. Dermatol. 2012, 21, 314–316. [Google Scholar] [CrossRef]
- Barrault, C.; Garnier, J.; Pedretti, N.; Cordier-Dirikoc, S.; Ratineau, E.; Deguercy, A.; Bernard, F.X. Androgens induce sebaceous differentiation in sebocyte cells expressing a stable functional androgen receptor. J. Steroid Biochem. Mol. Biol. 2015, 152, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Motoyoshi, K. Enhanced comedo formation in rabbit ear skin by squalene and oleic acid peroxides. Br. J. Dermatol. 1983, 109, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Yoshizawa, K.; Makino, I.; Kawakami, K.; Onoue, M. Comedogenicity of squalene monohydroperoxide in the skin after topical application. J. Toxicol. Sci. 2000, 25, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman, S.U.; Akhtar, N. Effect of Turmeric (Curcuma longa Zingiberaceae) Extract Cream on Human Skin Sebum Secretion. Trop. J. Pharm. Res. 2013, 12, 665–669. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Li, X.; Zhang, W.; Shen, X.; Lu, Y.; Li, H. Efficacy of a New Non-drug Acne Therapy: Aloe Vera Gel Combined With Ultrasound and Soft Mask for the Treatment of Mild to Severe Facial Acne. Front. Med. 2021, 8, 662640. [Google Scholar] [CrossRef]
- Nam, C.; Kim, S.; Sim, Y.; Chang, I. Anti-Acne Effects of Oriental Herb Extracts: A Novel Screening Method to Select Anti-Acne Agents. Ski. Pharmacol. Physiol. 2003, 16, 84–90. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2015, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Kim, K.Y. Inhibition of Proinflammatory Cytokines in Cutibacterium acnes-Induced Inflammation in HaCaT Cells by Using Buddleja davidii Aqueous Extract. Int. J. Inflamm. 2020, 2020, 8063289. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Chuang LTe Lien, T.J.; Liing, Y.R.; Chen, W.Y.; Tsai, P.J. Rosmarinus officinalis Extract Suppresses Propionibacterium acnes–Induced Inflammatory Responses. J. Med. Food 2013, 16, 324–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koseki, J.; Matsumoto, T.; Matsubara, Y.; Tsuchiya, K.; Mizuhara, Y.; Sekiguchi, K.; Nishimura, H.; Watanabe, J.; Kaneko, A.; Hattori, T.; et al. Inhibition of Rat 5α-Reductase Activity and Testosterone-Induced Sebum Synthesis in Hamster Sebocytes by an Extract of Quercus acutissima Cortex. Evid Based Complement. Alternat. Med. 2015, 2015, 853846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilforoushzadeh, M.A.; Aghdami, N.; Taghiabadi, E. Human Hair Outer Root Sheath Cells and Platelet-Lysis Exosomes Promote Hair Inductivity of Dermal Papilla Cell. Tissue Eng. Regen. Med. 2020, 17, 525–536. [Google Scholar] [CrossRef]
- Won, C.H.; Jeong, Y.M.; Kang, S.; Koo, T.S.; Park, S.H.; Park, K.Y.; Sung, Y.K.; Sung, J.H. Hair-growth-promoting effect of conditioned medium of high integrin α6 and low CD 71 (α6bri/CD71dim) positive keratinocyte cells. Int. J. Mol. Sci. 2015, 16, 4379–4391. [Google Scholar] [CrossRef] [Green Version]
- Watabe, Y.; Tomioka, M.; Watabe, A.; Aihara, M.; Shimba, S.; Inoue, H. The clock gene brain and muscle Arnt-like protein-1 (BMAL1) is involved in hair growth. Arch. Dermatol. Res. 2013, 305, 755–761. [Google Scholar] [CrossRef]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Lin, Z.L.; Lin, B.B.; Huang, Y.; Zhang, H.L.; et al. Fibroblast growth factors stimulate hair growth through β-catenin and Shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar] [CrossRef] [Green Version]
- Iida, M.; Ihara, S.; Matsuzaki, T. Hair cycle-dependent changes of alkaline phosphatase activity in the mesenchyme and epithelium in mouse vibrissal follicles. Dev. Growth Differ. 2007, 49, 185–195. [Google Scholar] [CrossRef]
- Shin, J.Y.; Kim, J.; Choi, Y.H.; Kang, N.G.; Lee, S. Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells. Curr. Issues Mol. Biol. 2021, 43, 1361–1373. [Google Scholar] [CrossRef]
- Saewan, N. Effect of Coffee Berry Extract on Anti-Aging for Skin and Hair—In Vitro Approach. Cosmetics 2022, 9, 66. [Google Scholar] [CrossRef]
- Emblica (Phyllanthus emblica Linn.) Fruit Extract Promotes Proliferation in Dermal Papilla Cells of Human Hair Follicle. Available online: https://scialert.net/fulltext/?doi=rjmp.2011.95.100 (accessed on 30 July 2022).
- Tiampasook, P.; Chaiyasut, C.; Sivamaruthi, B.S.; Timudom, T.; Nacapunchai, D. Effect of Phyllanthus emblica Linn. on Tensile Strength of Virgin and Bleached Hairs. Appl. Sci. 2020, 10, 6305. [Google Scholar] [CrossRef]
- Datta, K.; Singh, A.T.; Mukherjee, A.; Bhat, B.; Ramesh, B.; Burman, A.C. Eclipta alba extract with potential for hair growth promoting activity. J. Ethnopharmacol. 2009, 124, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Effect of Trigonella Foenum-Graecum Linn (seeds) and Butea Monosperma Lam (flowers) on Chemotherapy-Induced Alopecia. Available online: https://www.researchgate.net/publication/312163056_Effect_of_Trigonella_foenum-graecum_Linn_seeds_and_Butea_monosperma_Lam_flowers_on_chemotherapy-induced_alopecia (accessed on 30 July 2022).
- Adhirajan, N.; Ravi Kumar, T.; Shanmugasundaram, N.; Babu, M. In Vivo and in vitro evaluation of hair growth potential of Hibiscus rosa-sinensis Linn. J. Ethnopharmacol. 2003, 88, 235–239. [Google Scholar] [CrossRef]
- Koch, W.; Zagórska, J.; Marzec, Z.; Kukula-Koch, W. Applications of Tea (Camellia sinensis) and Its Active Constituents in Cosmetics. Molecules 2019, 24, 4277. [Google Scholar] [CrossRef] [Green Version]
- Evaluation of Clinical Efficacy and Safety of “Anti Dandruff Hair Cream” for the Treatment of Dandruff|Request PDF. Available online: https://www.researchgate.net/publication/284654042_Evaluation_of_clinical_efficacy_and_safety_of_anti_dandruff_hair_cream_for_the_treatment_of_dandruff (accessed on 30 July 2022).
- Clinical Evaluation of Herbal Hair Loss Cream in Management of Alopecia Aerata: An Open Study|Semantic Scholar. Available online: https://www.semanticscholar.org/paper/Clinical-evaluation-of-herbal-Hair-Loss-Cream-in-of-Ravichandran-Consultant/b923b7ec4c9eb41f09d37d671d926863de6fb3d5 (accessed on 30 July 2022).
- Thiboutot, D.; Harris, G.; Iles, V.; Cimis, G.; Gilliland, K.; Hagari, S. Activity of the type 1 5 alpha-reductase exhibits regional differences in isolated sebaceous glands and whole skin. J. Investig. Dermatol. 1995, 105, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Seiffert, K.; Seltmann, H.; Fritsch, M.; Zouboulis, C.C. Inhibition of 5alpha-reductase activity in SZ95 sebocytes and HaCaT keratinocytes in vitro. Horm. Metab. Res. 2007, 39, 141–148. [Google Scholar] [CrossRef]
- Jang, S.; Lee, Y.; Hwang, S.L.; Lee, M.H.; Park, S.J.; Lee, I.H.; Kang, S.; Roh, S.S.; Seo, Y.J.; Park, J.K.; et al. Establishment of type II 5alpha-reductase over-expressing cell line as an inhibitor screening model. J. Steroid Biochem. Mol. Biol. 2007, 107, 245–252. [Google Scholar] [CrossRef]
- Srivilai, J.; Rabgay, K.; Khorana, N.; Waranuch, N.; Nuengchamnong, N.; Ingkaninan, K. A new label-free screen for steroid 5α-reductase inhibitors using LC-MS. Steroids 2016, 116, 67–75. [Google Scholar] [CrossRef]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Vogeley, C.; Esser, C.; Tüting, T.; Krutmann, J.; Haarmann-Stemmann, T. Role of the Aryl Hydrocarbon Receptor in Environmentally Induced Skin Aging and Skin Carcinogenesis. Int. J. Mol. Sci. 2019, 20, 6005. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Ale-Agha, N.; Haendeler, J.; Ventura, N.; Vogeley, C.; Esser, C.; Tüting, T.; Krutmann, J.; Haarmann-Stemmann, T. The Aryl Hydrocarbon Receptor (AhR) in the Aging Process: Another Puzzling Role for This Highly Conserved Transcription Factor. Front. Physiol. 2020, 10, 1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.; Lee, S.; Lee, M.J.; Lee, D.H.; Won, C.H.; Kim, S.M.; Chung, J.H. Dietary Aloe Vera Supplementation Improves Facial Wrinkles and Elasticity and It Increases the Type I Procollagen Gene Expression in Human Skin in vivo. Ann. Dermatol. 2009, 21, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.K.; Kim, H.I.; Kim, S.Y.; Seo, H.H.; Song, J.; Kim, J.; Shin, D.S.; Jo, Y.; Choi, H.; Lee, J.H.; et al. Anti-Aging Effects of Leontopodium alpinum (Edelweiss) Callus Culture Extract through Transcriptome Profiling. Genes 2020, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.; Rana, A. Withania somnifera (Ashwagandha): A Review. Pharmacogn. Rev. 2007, 1, 129–136. [Google Scholar]
- Varma, S.R.; Sivaprakasam, T.O.; Mishra, A.; Kumar, L.M.S.; Prakash, N.S.; Prabhu, S.; Ramakrishnan, S. Protective Effects of Triphala on Dermal Fibroblasts and Human Keratinocytes. PLoS ONE 2016, 11, e0145921. [Google Scholar] [CrossRef]
- Chuarienthong, P.; Lourith, N.; Leelapornpisid, P. Clinical efficacy comparison of anti-wrinkle cosmetics containing herbal flavonoids. Int. J. Cosmet. Sci. 2010, 32, 99–106. [Google Scholar] [CrossRef]
- Benameur, T.; Soleti, R.; Panaro, M.A.; La Torre, M.E.; Monda, V.; Messina, G.; Porro, C. Curcumin as Prospective Anti-Aging Natural Compound: Focus on Brain. Molecules 2021, 26, 4794. [Google Scholar] [CrossRef]
- Bielak-Zmijewska, A.; Grabowska, W.; Ciolko, A.; Bojko, A.; Mosieniak, G.; Bijoch, Ł.; Sikora, E. The Role of Curcumin in the Modulation of Ageing. Int. J. Mol. Sci. 2019, 20, 1239. [Google Scholar] [CrossRef] [Green Version]
- Bylka, W.; Znajdek-Awizeń, P.; Studzińska-Sroka, E.; Brzezińska, M. Centella asiatica in cosmetology. Adv. Dermatol. Allergol. 2013, 1, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Little, E.; Quan, H.; Qin, Z.; Voorhees, J.J.; Fisher, G.J. Elevated matrix metalloproteinases and collagen fragmentation in photodamaged human skin: Impact of altered extracellular matrix microenvironment on dermal fibroblast function. J. Investig. Dermatol. 2013, 133, 1362–1366. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Jung, E.; Kim, Y.; Park, J.; Park, J.; Hong, S.; Kim, J.; Hyun, C.; Kim, Y.S.; Park, D. Asiaticoside induces human collagen I synthesis through TGFbeta receptor I kinase (TbetaRI kinase)-independent Smad signaling. Planta Med. 2006, 72, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.Y.; Choi, J.S.; Choi, Y.J.; Shin, S.Y.; Kang, S.W.; Han, S.J.; Kang, Y.H. (-)Epigallocatechin gallate hampers collagen destruction and collagenase activation in ultraviolet-B-irradiated human dermal fibroblasts: Involvement of mitogen-activated protein kinase. Food Chem. Toxicol. 2008, 46, 1298–1307. [Google Scholar] [CrossRef]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Dermatoendocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Choi, D.; Park, T. Decanal Protects against UVB-Induced Photoaging in Human Dermal Fibroblasts via the cAMP Pathway. Nutrients 2020, 12, 1214. [Google Scholar] [CrossRef]
- Lago, J.C.; Puzzi, M.B. The effect of aging in primary human dermal fibroblasts. PLoS ONE 2019, 14, e0219165. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.H.; Rhie, S.J.; Kim, Y.C. Antioxidant and Skin Anti-Aging Effects of Marigold Methanol Extract. Toxicol. Res. 2018, 34, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Tan, P.L.; Rajagopal, M.; Chinnappan, S.; Selvaraja, M.; Leong, M.Y.; Tan, L.F.; Yap, V.L. Formulation and Physicochemical Evaluation of Green Cosmeceutical Herbal Face Cream Containing Standardized Mangosteen Peel Extract. Cosmetics 2022, 9, 46. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlomagno, F.; Roveda, G.; Michelotti, A.; Ruggeri, F.; Tursi, F. Anti-Skin-Aging Effect of a Treatment with a Cosmetic Product and a Food Supplement Based on a New Hyaluronan: A Randomized Clinical Study in Healthy Women. Cosmetics 2022, 9, 54. [Google Scholar] [CrossRef]
- Foolad, N.; Vaughn, A.R.; Rybak, I.; Burney, W.A.; Chodur, G.M.; Newman, J.W.; Steinberg, F.M.; Sivamani, R.K. Prospective randomized controlled pilot study on the effects of almond consumption on skin lipids and wrinkles. Phyther. Res. 2019, 33, 3212–3217. [Google Scholar] [CrossRef] [Green Version]
- Hwang, E.; Lee, D.G.; Park, S.H.; Oh, M.S.; Kim, S.Y. Coriander Leaf Extract Exerts Antioxidant Activity and Protects Against UVB-Induced Photoaging of Skin by Regulation of Procollagen Type I and MMP-1 Expression. J. Med. Food 2014, 17, 985–995. [Google Scholar] [CrossRef]
- Bralley, E.; Greenspan, P.; Hargrove, J.L.; Hartle, D.K. Pharmaceutical Biology Inhibition of Hyaluronidase Activity by Vitis rotundifolia. (Muscadine) Berry Seeds and Skins Inhibition of Hyaluronidase Activity by Vitis rotundifolia (Muscadine) Berry Seeds and Skins. Pharm. Biol. 2007, 45, 667–673. [Google Scholar] [CrossRef]
- Wlaschek, M.; Maity, P.; Makrantonaki, E.; Scharffetter-Kochanek, K. Connective Tissue and Fibroblast Senescence in Skin Aging. J. Investig. Dermatol. 2021, 141, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Ng, S.C.; Hsu, J.Y.; Liu, H.; Chen, C.J.; Huang, C.Y.; Kuo, W.W. Galangin Reverses H 2 O 2-Induced Dermal Fibroblast Senescence via SIRT1-PGC-1α/Nrf2 Signaling. Int. J. Mol. Sci. 2022, 23, 1387. [Google Scholar] [CrossRef]
- Ho, C.Y.; Dreesen, O. Faces of cellular senescence in skin aging. Ageing Dev. 2021, 198, 111525. [Google Scholar] [CrossRef]
- Fuhrmann-Stroissnigg, H.; Santiago, F.E.; Grassi, D.; Ling, Y.Y.; Niedernhofer, L.J.; Robbins, P.D. SA-β-Galactosidase-Based Screening Assay for the Identification of Senotherapeutic Drugs. J. Vis. Exp. 2019, 148, e58133. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.; Shin, S.; Cho, E.; Ryu, D.; Garandeau, D.; Chajra, H.; Fréchet, M.; Park, D.; Jung, E. Senotherapeutic-like effect of Silybum marianum flower extract revealed on human skin cells. PLoS ONE 2021, 16, e0260545. [Google Scholar] [CrossRef]
- Jeon, S.; Choi, M. Anti-inflammatory and anti-aging effects of hydroxytyrosol on human dermal fibroblasts (HDFs). Biomed. Dermatol. 2018, 2, 21. [Google Scholar] [CrossRef]
- Kim, H.J.; Song, S.B.; Choi, J.M.; Kim, K.M.; Cho, B.K.; Cho, D.H.; Park, H.J. IL-18 downregulates collagen production in human dermal fibroblasts via the ERK pathway. J. Investig. Dermatol. 2010, 130, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Kabashima, T.; Rahman, M.S.; Shibata, T.; Kai, M. Amplified and selective assay of collagens by enzymatic and fluorescent reactions. Sci. Rep. 2014, 4, 4950. [Google Scholar] [CrossRef] [Green Version]
- Fosang, A.J.; Hey, N.J.; Carney, S.L.; Hardinghami, T.E. An Elisa Plate Based Assay for Hyaluronan Using Biotinylated Proteoglycan G1 Domain (HA-Binding Region). Matrix 1990, 10, 306–313. [Google Scholar] [CrossRef]
- Thring, T.S.A.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genc, Y.; Dereli, F.T.G.; Saracoglu, I.; Akkol, E.K. The inhibitory effects of isolated constituents from Plantago major subsp. major L. on collagenase, elastase and hyaluronidase enzymes: Potential wound healer. Saudi Pharm. J. 2019, 28, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O.; et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Furue, M. Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic Implications in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Eckert, R.L.; Sturniolo, M.T.; Broome, A.M.; Ruse, M.; Rorke, E.A. Transglutaminase Function in Epidermis. J. Investig. Dermatol. 2005, 124, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Boury-Jamot, M.; Sougrat, R.; Tailhardat, M.; Le Varlet, B.; Bonté, F.; Dumas, M.; Verbavatz, J.-M. Expression and function of aquaporins in human skin: Is aquaporin-3 just a glycerol transporter? Biochim. Biophys. Acta Biomembr. 2006, 1758, 1034–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochénska, K.; Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Gabig-Cimińska, M. Models in the Research Process of Psoriasis. Int. J. Mol. Sci. 2017, 18, 2514. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, B.; Ceribelli, A.; Massarotti, M.S.; Selmi, C. The Th17 axis in psoriatic disease: Pathogenetic and therapeutic implications. AutoImmun. Highlights 2014, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varma, S.R.; Sivaprakasam, T.O.; Mishra, A.; Prabhu, S.; Rafiq, M.; Rangesh, P. Imiquimod-induced psoriasis-like inflammation in differentiated Human keratinocytes: Its evaluation using curcumin. Eur. J. Pharmacol. 2017, 813, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Morizane, S.; Gallo, R.L. Antimicrobial peptides in the pathogenesis of psoriasis. J. Dermatol. 2012, 39, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Ekman, A.K.; Vegfors, J.; Bivik Eding, C.; Enerbäck, C. Overexpression of psoriasin (S100A7) contributes to dysregulated differentiation in psoriasis. Acta Derm. Venereol. 2017, 97, 441–448. [Google Scholar] [CrossRef] [Green Version]
- New Biologics in Psoriasis: An Update on IL-23 and IL-17 Inhibitors. Available online: https://pubmed.ncbi.nlm.nih.gov/28319618/ (accessed on 29 July 2022).
- Rønholt, K.; Iversen, L. Old and New Biological Therapies for Psoriasis. Int. J. Mol. Sci. 2017, 18, 2297. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.L.; Schinella, G.R.; Andújar, I. Antipsoriatic Medicinal Plants: From Traditional Use to Clinic. In Ethnobotany, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 158–186. [Google Scholar] [CrossRef]
- Wadher, K.; Dabre, S.; Gaidhane, A.; Trivedi, S.; Umekar, M. Evaluation of antipsoriatic activity of gel containing Pongamia pinnata extract on Imiquimod-induced psoriasis. Clin. Phytosci. 2021, 7, 1–6. [Google Scholar] [CrossRef]
- Divakara, P.; Nagaraju, B.; Buden, R.P.; Sekhar, H.S.; Ravi, C.M. Antipsoriatic activity of ayurvedic ointment containing aqueous extract of the bark of Pongamia Pinnata using the rat ultraviolet ray photodermatitis model. Adv. Med. Plant Res. 2013, 1, 8–16. [Google Scholar]
- Sangiovanni, E.; Di Lorenzo, C.; Piazza, S.; Manzoni, Y.; Brunelli, C.; Fumagalli, M.; Magnavacca, A.; Martinelli, G.; Colombo, F.; Casiraghi, A.; et al. Vitis vinifera L. Leaf Extract Inhibits In Vitro Mediators of Inflammation and Oxidative Stress Involved in Inflammatory-Based Skin Diseases. Antioxidants 2019, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [Green Version]
- Yue, P.Y.K.; Leung, E.P.Y.; Mak, N.K.; Wong, R.N.S. A simplified method for quantifying cell migration/wound healing in 96-well plates. J. Biomol. Screen 2010, 15, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Wang, Y.L.; Ren, F.; Lele, T.P. Stamp wound assay for studying coupled cell migration and cell debris clearance. Langmuir 2010, 26, 16672–16676. [Google Scholar] [CrossRef]
- Hettler, A.; Werner, S.; Eick, S.; Laufer, S.; Weise, F. A New In Vitro Model to Study Cellular Responses after Thermomechanical Damage in Monolayer Cultures. PLoS ONE 2013, 8, e82635. [Google Scholar] [CrossRef]
- Keese, C.R.; Wegener, J.; Walker, S.R.; Giaever, I. Electrical wound-healing assay for cells in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 1554–1559. [Google Scholar] [CrossRef] [Green Version]
- Zordan, M.D.; Mill, C.P.; Riese, D.J.; Leary, J.F. A high throughput, interactive imaging, bright-field wound healing assay. Cytom. Part A 2011, 79, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Fronza, M.; Heinzmann, B.; Hamburger, M.; Laufer, S.; Merfort, I. Determination of the wound healing effect of Calendula extracts using the scratch assay with 3T3 fibroblasts. J. Ethnopharmacol. 2009, 126, 463–467. [Google Scholar] [CrossRef]
- Fox, L.T.; Mazumder, A.; Dwivedi, A.; Gerber, M.; du Plessis, J.; Hamman, J.H. In Vitro wound healing and cytotoxic activity of the gel and whole-leaf materials from selected aloe species. J. Ethnopharmacol. 2017, 200, 1–7. [Google Scholar] [CrossRef]
- Razia, S.; Park, H.; Shin, E.; Shim, K.S.; Cho, E.; Kang, M.C.; Kim, S.Y. Synergistic effect of Aloe vera flower and Aloe gel on cutaneous wound healing targeting MFAP4 and its associated signaling pathway: In-vitro study. J. Ethnopharmacol. 2022, 290, 115096. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Razzell, W.; Martin, P. ‘White wave’ analysis of epithelial scratch wound healing reveals how cells mobilise back from the leading edge in a myosin-II-dependent fashion. J. Cell Sci. 2011, 124, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Zaja-Milatovic, S.; Richmond, A. CXC chemokines and their receptors: A case for a significant biological role in cutaneous wound healing. Histol. Histopathol. 2008, 23, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Eckes, B.; Mauch, C.; Hartmann, K.; Krieg, T. Monocyte Chemoattractant Protein-1 Enhances Gene Expression and Synthesis of Matrix Metalloproteinase-1 in Human Fibroblasts by an Autocrine IL-1α Loop. J. Immunol. 2000, 164, 6174–6179. [Google Scholar] [CrossRef] [Green Version]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The role of chemokines in wound healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.; Tsou, R.; Wallace, K.; Gibran, N.; Isik, F. Early gene expression profile of human skin to injury using high-density cDNA microarrays. Wound Repair Regen. 2001, 9, 360–370. [Google Scholar] [CrossRef]
- Wiegand, C.; Hipler, U.C.; Elsner, P.; Tittelbach, J. Keratinocyte and fibroblast wound healing in vitro is repressed by non-optimal conditions but the reparative potential can be improved by water-filtered infrared A. Biomedicines 2021, 9, 1802. [Google Scholar] [CrossRef]
- Nowinski, D.; Lysheden, A.S.; Gardner, H.; Rubin, K.; Gerdin, B.; Ivarsson, M. Analysis of Gene Expression in Fibroblasts in Response to Keratinocyte-Derived Factors In Vitro: Potential Implications for the Wound Healing Process. J. Investig. Dermatol. 2004, 122, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Leng, H.; Pu, L.; Xu, L.; Shi, X.; Ji, J.; Chen, K. Effects of aloe polysaccharide, a polysaccharide extracted from Aloe vera, on TNF-α-induced HaCaT cell proliferation and the underlying mechanism in psoriasis. Mol. Med. Rep. 2018, 18, 3537–3543. [Google Scholar] [CrossRef] [Green Version]
- Wahedi, H.M.; Jeong, M.; Chae, J.K.; Do, S.G.; Yoon, H.; Kim, S.Y. Aloesin from Aloe vera accelerates skin wound healing by modulating MAPK/Rho and Smad signaling pathways in vitro and in vivo. Phytomedicine 2017, 28, 19–26. [Google Scholar] [CrossRef]
- Ju Woo, H.; Jun, D.Y.; Lee, J.Y.; Park, H.S.; Woo, M.H.; Park, S.J.; Kim, S.C.; Yang, C.H.; Kim, Y.H. Anti-inflammatory action of 2-carbomethoxy-2,3-epoxy-3-prenyl-1,4-naphthoquinone (CMEP-NQ) suppresses both the MyD88-dependent and TRIF-dependent pathways of TLR4 signaling in LPS-stimulated RAW264.7 cells. J. Ethnopharmacol. 2017, 205, 103–115. [Google Scholar] [CrossRef]
- Evaluation of Antioxidant and Anti-Melanogenic Activities of Different Extracts from Aerial Parts of Nepeta Binaludensis Jamzad in Murine Melanoma B16F10 Cells. Available online: https://pubmed.ncbi.nlm.nih.gov/27482348/ (accessed on 29 July 2022).
- Burger, P.; Landreau, A.; Azoulay, S.; Michel, T.; Fernandez, X. Skin Whitening Cosmetics: Feedback and Challenges in the Development of Natural Skin Lighteners. Cosmetics 2016, 3, 36. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Yotsumoto, H.; Tian, Y.; Sakamoto, K. α-Mangostin suppressed melanogenesis in B16F10 murine melanoma cells through GSK3β and ERK signaling pathway. Biochem. Biophys. Rep. 2021, 26, 100949. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Lee, J.W.; Lee, T.H.; Subedi, L.; Wahedi, H.M.; Do, S.G.; Shin, E.; Moon, E.Y.; Kim, S.Y. UP256 Inhibits Hyperpigmentation by Tyrosinase Expression/Dendrite Formation via Rho-Dependent Signaling and by Primary Cilium Formation in Melanocytes. Int. J. Mol. Sci. 2020, 21, 5341. [Google Scholar] [CrossRef]
- Lajis, A.F.B.; Hamid, M.; Ariff, A.B. Depigmenting effect of Kojic acid esters in hyperpigmented B16F1 melanoma cells. J. Biomed. Biotechnol. 2012, 2012, 952452. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, Y.; Hearing, V.J. Physiological factors that regulate skin pigmentation. BioFactors 2009, 35, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Noh, S.G.; Park, Y.; Kang, D.; Chun, P.; Chung, H.Y.; Jung, H.J.; Moon, H.R. A Potent Tyrosinase Inhibitor, ( E)-3-(2,4-Dihydroxyphenyl)-1-(thiophen-2-yl)prop-2-en-1-one, with Anti-Melanogenesis Properties in α-MSH and IBMX-Induced B16F10 Melanoma Cells. Molecules 2018, 23, 2725. [Google Scholar] [CrossRef] [Green Version]
- Moreiras, H.; Seabra, M.C.; Barral, D.C. Melanin Transfer in the Epidermis: The Pursuit of Skin Pigmentation Control Mechanisms. Int. J. Mol. Sci. 2021, 22, 4466. [Google Scholar] [CrossRef]
- Greatens, A.; Hakozaki, T.; Koshoffer, A.; Epstein, H.; Schwemberger, S.; Babcock, G.; Bissett, D.; Takiwaki, H.; Arase, S.; Wickett, R.R.; et al. Effective inhibition of melanosome transfer to keratinocytes by lectins and niacinamide is reversible. Exp. Dermatol. 2005, 14, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Nizard, C.; Kurfurst, R.; Bonte, F.; Schnebert, S.; Tobin, D.J. The silver locus product (Silv/gp100/Pmel17) as a new tool for the analysis of melanosome transfer in human melanocyte-keratinocyte co-culture. Exp. Dermatol. 2008, 17, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Shieh, B.H.; Lu, M.H.; Chen, J.Y.; Chang, L.T.; Chao, C.F. A method for quantifying melanosome transfer efficacy from melanocytes to keratinocytes in vitro. Pigment Cell Melanoma Res. 2008, 21, 559–564. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Assay | Significance of Assay | Model System Used and Evaluation Strategy | End Point Measured |
|---|---|---|---|
| Anti-microbial activity | Anti-microbial activity against C. acnes is important for the anti-acne effects of the ingredients | C. acnes bacterial culture | Anti-bacterial activity of the ingredients using assays such as agar diffusion assay [35] |
| Anti-inflammatory activity | Inflammation is an integral part of the pathogenesis of acne vulgaris, and the anti-inflammatory activity of the ingredients can offer good protection against acne. | Monocyte cell lines (THP-1), keratinocytes (HaCaT cell line), or human sebocyte cell line. (inflammation induced in the cells by exposure to heat-inactive C. acnes or peptidoglycans [16] | Effect of cosmetic ingredients on C. acnes-induced inflammation as measured by the levels of inflammatory cytokines in the cell culture supernatant or expression of the respective genes in the cells [36,37] |
| Sebum quality and sebostatic activity | Sebum quality and quantity is an important parameter that can contribute to the progression of the pathology of acne vulgaris | Human sebocytic cell lines |
|
| Type of Assay | Significance of Assay | Model System Used and Evaluation Strategy | End Point Measured |
|---|---|---|---|
| Proliferation of the follicle cells | Hair follicle cells are primarily responsible for the growth of hair. | HFPDPC or ORSc cells are used as model systems. The cells are grown in the presence of different doses of the test substance for different time points: 24, 48, and 72 h | Effect of the ingredients on proliferation of HFPDPC or ORSc cells using standard cell-proliferation-based assays [40]. |
| Inhibition of 5-α reductase | 5-α reductase converts testosterone into a more potent form, i.e., DHT. DHT is known to be involved in male pattern baldness. | Keratinocyte and sebocytes that are known to exhibit detectable levels of 5-α reductase activity (should be individually established in the labs since the activity levels may depend on the cell type) | Effect of the ingredients on the enzyme activity of 5-α reductase using testosterone as substrate. The detection of the DHT can be based on radioactive detection, TLC, HPLC, or LC-MS [38,57] |
| Type of Assay | Significance of Assay | Model System Used and Evaluation Strategy | End Point Measured |
|---|---|---|---|
| Estimation of collagen levels | Collagen is important for skin integrity. Its levels are known to reduce with aging, which results in visible phenotypes such as skin wrinkles | Human dermal fibroblasts (e.g., HDF or HS68) are treated with test substances, and collagen levels are estimated by using biochemical methods or ELISA [91,92] | Collagen levels |
| Estimation of hyaluronic acid levels | Hyaluronic acid is an important component of ECM, and it is known to reduce with aging | ELISA-based estimation of HA in human fibroblasts such as HDF in presence of test substances [93] | Hyaluronic acid levels |
| Estimation of enzyme activities of collagenase, elastase, hyaluronidase, and HA synthases | These enzymes affect the levels of key ECM components such as collagen, elastin, and hyaluronic acid | Effect of cosmetic ingredients on the activities of these enzymes are tested by respective biochemical methods. Human fibroblasts (HDF) or keratinocytes (HaCaT) can be employed [94,95] | Enzyme activities |
| Photoaging | Exposure to UV is one of the triggers for age associated changes in the skin | Human dermal fibroblasts (HDF or HS68) cells are exposed to UV light and the effect of the test substance is evaluated [72] |
|
| Cellular senescence | Cellular senescence causes irreversible cessation of cell proliferation and increases with aging | Human dermal fibroblasts are exposed to either UV light or hydrogen peroxide to induce cellular senescence; test substances are evaluated for their effectiveness to regulate or reverse the senescence [87,96] |
|
| Wound Creation Methods | Pros | Cons | |
|---|---|---|---|
| Mechanical wounding (scratch assay) | Scratch introduced mechanically on cell monolayer using a pipette tip or cell scraper or stamping with stamp molds [115,116] | Simple technique, easily available tools | Irregular wound area created, scraped cells may be retained at the edges, impairing the data |
| Thermo-mechanical wounding | Temperature-controlled stamp molds [117] | Thermal and mechanical damage healing can be analyzed | Heat may affect surrounding cells of the wound area, impairing data |
| Electrical wounding | Cells grown on electrodes, scratch made on the cell layer, change in impedance studied | Accurate, reproducible data | Heating of bystander cells surrounding the wound; specialized tools required to perform electrical cell-substrate impedance sensing (ECIS) [118] |
| Optical wounding | Laser beam pulses used to create precise wounds | Precise, high-throughput, reproducible wounds and data created | Specialized equipment required: Laser-enabled analysis and processing (LEAP) instrument [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Bhat, S.S.; Singh, N.; Venkanna, B.U.; Mohamed, R.; Rao, R.P. Cell-Based Model Systems for Validation of Various Efficacy-Based Claims for Cosmetic Ingredients. Cosmetics 2022, 9, 107. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics9050107
Singh P, Bhat SS, Singh N, Venkanna BU, Mohamed R, Rao RP. Cell-Based Model Systems for Validation of Various Efficacy-Based Claims for Cosmetic Ingredients. Cosmetics. 2022; 9(5):107. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics9050107
Chicago/Turabian StyleSingh, Prachi, Shruthi S. Bhat, Nikhil Singh, Babu U. Venkanna, Rafiq Mohamed, and Raghavendra P. Rao. 2022. "Cell-Based Model Systems for Validation of Various Efficacy-Based Claims for Cosmetic Ingredients" Cosmetics 9, no. 5: 107. https://0-doi-org.brum.beds.ac.uk/10.3390/cosmetics9050107