
Vitamin-D Binding Protein Gene Polymorphisms and Serum 25-Hydroxyvitamin-D in a Turkish Population

 , ,
, ,
Abstract
:1. Introduction
2. Results
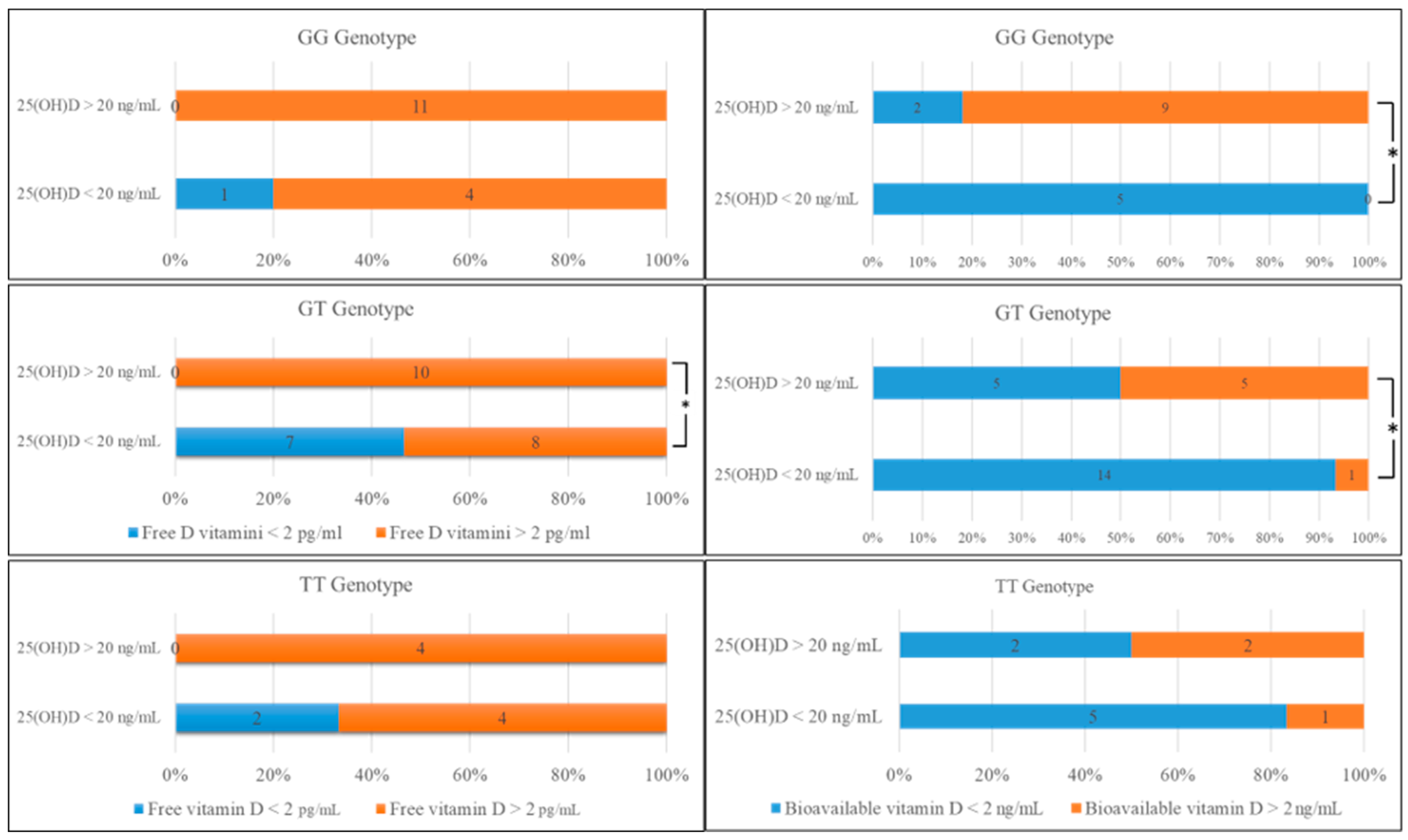
2.1. Comparison of Biochemical Data and Genotype Distribution in rs7041
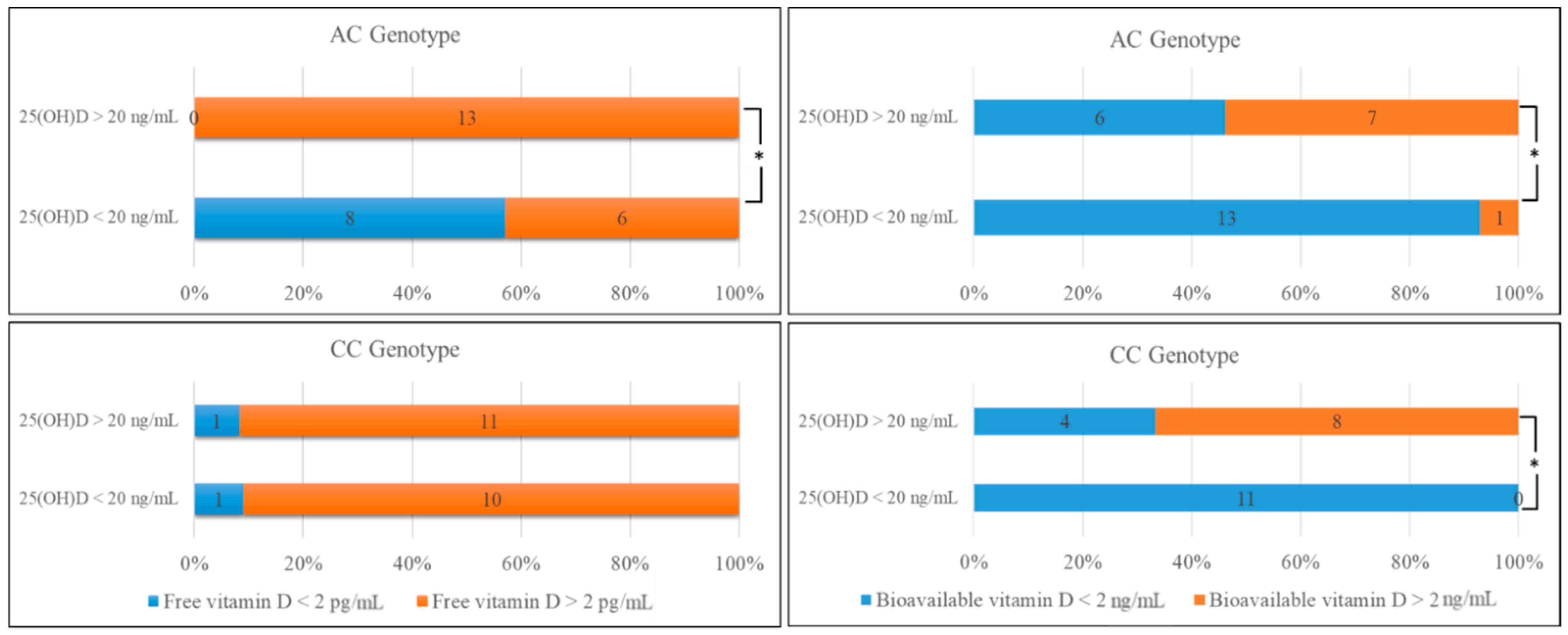
2.2. Comparison of the Biochemical Data and Genotype Distribution in rs4588
2.3. Comparison of Biochemical Data According to DBP Phenotypes
3. Discussion
4. Materials and Methods
4.1. Patient Selection
4.2. Blood Samples
4.3. Measurement of Serum 25(OH)D, Albumin, and Vitamin D-Binding Protein Concentrations
4.4. LC-MS/MS Method
4.5. DNA Isolation
4.6. Real-Time PCR
4.7. Calculation of Free 25(OH)D Levels
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powe, C.E.; Ricciardi, C.; Berg, A.H.; Erdenesanaa, D.; Collerone, G.; Ankers, E.; Wenger, J.; Karumanchi, S.A.; Thadhani, R.; Bhan, I. Vitamin D–binding protein modifies the vitamin D–bone mineral density relationship. J. Bone Miner. Res. 2011, 26, 1609–1616. [Google Scholar] [CrossRef]
- Bikle, D.D.; Schwartz, J. Vitamin D binding protein, total and free Vitamin D levels in different physiological and pathophysiological conditions. Front. Endocrinol. 2019, 10, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, C.M. The free hormone hypothesis: A physiologically based mathematical model. Endocr. Rev. 1989, 10, 232–274. [Google Scholar] [CrossRef]
- Bikle, D.D.; Gee, E. Free, and not total, 1,25-dihydroxyvitamin d regulates 25-hydroxyvitamin d metabolism by keratinocytes. Endocrinology 1989, 124, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Safadi, F.F.; Thornton, P.; Magiera, H.; Hollis, B.W.; Gentile, M.; Haddad, J.G.; Liebhaber, S.A.; Cooke, N.E. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J. Clin. Investig. 1999, 103, 239–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.; Hyppönen, E. Determinants of vitamin D status: Focus on genetic variations. Curr. Opin. Nephrol. Hypertens. 2011, 20, 331–336. [Google Scholar] [CrossRef]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Al-Daghri, N.M.; Mohammed, A.K.; Bukhari, I.; Rikli, M.; Abdi, S.; Ansari, M.G.A.; Sabico, S.; Hussain, S.D.; Alenad, A.; Al-Saleh, Y.; et al. Efficacy of vitamin D supplementation according to vitamin D-binding protein polymorphisms. Nutrition 2019, 63-64, 148–154. [Google Scholar] [CrossRef]
- Chun, R.; Peercy, B.E.; Orwoll, E.S.; Nielson, C.M.; Adams, J.S.; Hewison, M. Vitamin D and DBP: The free hormone hypothesis revisited. J. Steroid Biochem. Mol. Biol. 2014, 144, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, R.A.; Larsen, L.H.; Damsgaard, C.T.; Sørensen, L.B.; Hjorth, M.F.; Andersen, R.; Tetens, I.; Krarup, H.; Ritz, C.; Astrup, A.; et al. Common genetic variants are associated with lower serum 25-hydroxyvitamin D concentrations across the year among children at northern latitudes. Br. J. Nutr. 2017, 117, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leblanc, E.; Chou, R.; Zakher, B.; Daeges, M.; Pappas, M. Screening for Vitamin D Deficiency: Systematic Review for the U.S. Preventive Services Task Force Recommendation; Evidence Syntheses, No. 119; Agency for Healthcare Research and Quality (US): Rockville, MD, USA, 2014. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK263420/ (accessed on 11 August 2020).
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: What clinicians need to know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- Hekimsoy, Z.; Dinç, G.; Kafesçiler, S.; Onur, E.; Güvenç, Y.; Pala, T.; Güçlü, F.; Özmen, B. Vitamin D status among adults in the Aegean region of Turkey. BMC Public Health 2010, 10, 782. [Google Scholar] [CrossRef] [Green Version]
- Chishimba, L.; Thickett, D.; Stockley, R.A.; Wood, A.M. The vitamin D axis in the lung: A key role for vitamin D-binding protein. Thorax 2010, 65, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, A.M.; Bassford, C.; Webster, D.; Newby, P.; Rajesh, P.; Stockley, R.A.; Thickett, D. Vitamin D-binding protein contributes to COPD by activation of alveolar macrophages. Thorax 2011, 66, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, J.; Constans, J. Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum. Genet. 1993, 92, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.J.; Allison, R.; Close, G.L. Vitamin D and the Athlete: Current Perspectives and New Challenges. Sports Med. 2018, 48, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloia, J.; Mikhail, M.; Dhaliwal, R.; Shieh, A.; Usera, G.; Stolberg, A.; Ragolia, L.; Islam, S. Free 25(OH)D and the Vitamin D paradox in African Americans. J. Clin. Endocrinol. Metab. 2015, 100, 3356–3363. [Google Scholar] [CrossRef]
- Santos, B.R.; Mascarenhas, L.P.G.; Boguszewski, M.C.; Spritzer, P.M. Variations in the vitamin d-binding protein (DBP) gene are related to lower 25-hydroxyvitamin d levels in healthy girls: A cross-sectional study. Horm. Res. Paediatr. 2013, 79, 162–168. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, M.; Zhao, J.; Ren, F.; Chen, Y.; Li, J.-F.; Zhang, J.-Y.; Qu, F.; Zhang, J.-L.; Duan, Z.-P.; et al. Serum vitamin D3does not correlate with liver fibrosis in chronic hepatitis C. World J. Gastroenterol. 2015, 21, 11152–11159. [Google Scholar] [CrossRef]
- Miki, D.; Ochi, H.; Hayes, C.N.; Abe, H.; Yoshima, T.; Aikata, H.; Ikeda, K.; Kumada, H.; Toyota, J.; Morizono, T.; et al. Variation in the DEPDC5 locus is associated with progression to hepatocellular carcinoma in chronic hepatitis C virus carriers. Nat. Genet. 2011, 43, 797–800. [Google Scholar] [CrossRef]
- Ladero, J.M.; Torrejón, M.J.; Sánchez-Pobre, P.; Suárez, A.; Cuenca, F.; de la Orden, V.; Devesa, M.J.; Rodrigo, M.; Estrada, V.; López-Alonso, G.; et al. Vitamin D deficiency and vitamin D therapy in chronic hepatitis C. Ann. Hepatol. 2013, 12, 199–204. [Google Scholar] [CrossRef]
- Shehab, H.; Sabry, D.; Mukhtar, M.; Elakel, W.; Mabrouk, M.; Esmat, G.; El Kazaz, A.; Zahra, A. Vitamin D Receptor FokI Gene Polymorphism Predicted Poor Response to Treatment in Chronic HCV Genotype 4. Int. J. Biomed. 2016, 6, 265–270. [Google Scholar] [CrossRef]
- Shao, B.; Jiang, S.; Muyiduli, X.; Wang, S.; Mo, M.; Li, M.; Wang, Z.; Yu, Y. Vitamin D pathway gene polymorphisms influenced vitamin D level among pregnant women. Clin. Nutr. 2018, 37 Pt A, 2230–2237. [Google Scholar] [CrossRef]
- Scalioni, L.D.P.; dos Santos, B.R.; Spritzer, P.M.; Villela-Nogueira, C.A.; Lewis-Ximenez, L.L.; Pollo-Flores, P.; Esberard, E.B.C.; Brandão-Mello, C.E.; Lampe, E.; Villar, L.M. Impact of vitamin D receptor and binding protein gene polymorphisms in clinical and laboratory data of HCV patients. Medicine 2018, 97, e9881. [Google Scholar] [CrossRef] [PubMed]
- Uitterlinden, A.G.; Fang, Y.; van Meurs, J.B.; Pols, H.A.; van Leeuwen, J.P. Genetics and biology of vitamin D receptor polymorphisms. Gene 2004, 338, 143–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looker, A.; Dawson-Hughes, B.; Calvo, M.; Gunter, E.; Sahyoun, N. Serum 25-hydroxyvitamin D status of adolescents and adults in two seasonal subpopulations from NHANES III. Bone 2002, 30, 771–777. [Google Scholar] [CrossRef]
- van der Wielen, R.; de Groot, L.; van Staveren, W.; Löwik, M.; Berg, H.V.D.; Haller, J.; Moreiras, O. Serum vitamin D concentrations among elderly people in Europe. Lancet 1995, 346, 207–210. [Google Scholar] [CrossRef]
- Hyppönen, E.; Power, C. Hypovitaminosis D in British adults at age 45 y: Nationwide cohort study of dietary and lifestyle predictors. Am. J. Clin. Nutr. 2007, 85, 860–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, O.M.; Farwell, W.R.; Kermah, D.; Taylor, E.N. Racial differences in the relationship between vitamin D, bone mineral density, and parathyroid hormone in the National Health and Nutrition Examination Survey. Osteoporos. Int. 2010, 22, 1745–1753. [Google Scholar] [CrossRef] [Green Version]
- Powe, C.E.; Evans, M.K.; Wenger, J.; Zonderman, A.B.; Berg, A.H.; Nalls, M.; Tamez, H.; Zhang, D.; Bhan, I.; Karumanchi, S.A.; et al. Vitamin D–Binding Protein and Vitamin D Status of Black Americans and White Americans. N. Engl. J. Med. 2013, 369, 1991–2000. [Google Scholar] [CrossRef] [Green Version]
- Nielson, C.M.; Jones, K.S.; Chun, R.F.; Jacobs, J.M.; Wang, Y.; Hewison, M.; Adams, J.S.; Swanson, C.M.; Lee, C.G.; Vanderschueren, D.; et al. Free 25-Hydroxyvitamin D: Impact of Vitamin D Binding Protein Assays on Racial-Genotypic Associations. J. Clin. Endocrinol. Metab. 2016, 101, 2226–2234. [Google Scholar] [CrossRef] [Green Version]
- Henderson, C.M.; Lutsey, P.L.; Misialek, J.R.; Laha, T.J.; Selvin, E.; Eckfeldt, J.H.; Hoofnagle, A.N. Measurement by a novel LC-MS/MS methodology reveals similar serum concentrations of vitamin D–binding protein in blacks and whites. Clin. Chem. 2016, 62, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Törölä, T.; Li, C.-X.; Ohlmeier, S.; Toljamo, T.; Nieminen, P.; Hattori, N.; Pulkkinen, V.; Iwamoto, H.; Mazur, W. Sputum Vitamin D Binding Protein (VDBP) GC1S/1S Genotype Predicts Airway Obstruction: A Prospective Study in Smokers with COPD. Int. J. Chronic Obstr. Pulm. Dis. 2020, 15, 1049–1059. [Google Scholar] [CrossRef]
- Hoofnagle, A.N.; Eckfeldt, J.H.; Lutsey, P.L. Vitamin D–Binding Protein Concentrations Quantified by Mass Spectrometry. N. Engl. J. Med. 2015, 373, 1480–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heureux, N.; Lindhout, E.; Swinkels, L. A direct assay for measuring free 25-hydroxyvitamin D. J. AOAC Int. 2017, 100, 1318–1322. [Google Scholar] [CrossRef]
- Jorde, R.; Schirmer, H.; Wilsgaard, T.; Bøgeberg Mathiesen, E.; Njølstad, I.; Løchen, M.L.; Joakimsen, R.M.; Grimnes, G. The DBP phenotype Gc-1f/Gc-1f is associated with reduced risk of cancer. The Tromsø Study. PLoS ONE 2015, 10, e0126359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekkinen, M.; Saarnio, E.; Viljakainen, H.T.; Kokkonen, E.; Jakobsen, J.; Cashman, K.; Mäkitie, O.; Lamberg-Allardt, C. Vitamin D binding protein genotype is associated with serum 25-hydroxyvitamin D and PTH concentrations, as well as bone health in children and adolescents in finland. PLoS ONE 2014, 9, e87292. [Google Scholar] [CrossRef] [Green Version]
- Sobczynska-Malefora, A.; Harbige, L.; Steed, D.; MacMahon, E.; Shankar-Hari, M.; Harrington, D. Vitamin-D deficiency is more common in UK black people and a possible COVID-19 risk factor. Ann. Nutr. Metab. 2021, 21, 451–452. [Google Scholar] [CrossRef]
- Mondul, A.M.; Weinstein, S.J.; Virtamo, J.; Albanes, D. Influence of vitamin D binding protein on the association between circulating vitamin D and risk of bladder cancer. Br. J. Cancer 2012, 107, 1589–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peršić, V.; Raljević, D.; Markova-Car, E.; Cindrić, L.; Miškulin, R.; Žuvić, M.; Pavelić, S.K. Vitamin D-binding protein (rs4588) T/T genotype is associated with anteroseptal myocardial infarction in coronary artery disease patients. Ann. Transl. Med. 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genotypes X ± SD | p Values | |||
|---|---|---|---|---|
| GG (n = 16) | GT (n = 25) | TT (n = 10) | ||
| LC-MS/MS-25(OH)D (ng/mL) | 27.68 ± 13.69 | 17.33 ± 10.71 * | 12.95 ± 5.52 * | 0.005 |
| DBP (mg/L) | 391.83 ± 91.10 | 409.64 ± 162.41 | 337.4 ± 98.45 | 0.134 |
| Free 25(OH)D (pg/mL) | 5.73 ± 2.58 | 3.62 ± 2.20 * | 2.74 ± 1.36 * | 0.006 |
| Bioavailable 25(OH)D (ng/mL) | 2.35 ± 1.03 | 1.43 ± 0.85 * | 1.27 ± 0.65 * | 0.004 |
| n (%) | Free 25(OH)D < 2 pg/mL (n = 10) | Free 25(OH)D > 2 pg/mL (n = 41) | p Values | Bioavailable 25(OH)D < 2 ng/mL (n = 33) | Bioavailable 25(OH)D > 2 ng/mL (n = 18) | p Values | |
|---|---|---|---|---|---|---|---|
| Genotype distribution | GG | 1 (6) | 15 (94) | 0.052 | 7 (43.75) | 9 (56.25) * | 0.017 |
| GT | 7 (28) | 18 (72) * | 0.033 | 19 (76) | 6 (24) * | 0.049 | |
| TT | 2 (20) | 8 (80) | 0.486 | 7 (70) | 3 (30) | 0.348 | |
| Allele frequency | G | 0.80 | 0.80 | 0.259 | 0.79 | 0.83 | 0.155 |
| T | 0.90 | 0.63 | 0.79 | 0.50 | |||
| Genotypes X ± SD | |||
|---|---|---|---|
| AC (n = 27) | CC (n = 23) | p Values | |
| LC-MS/MS-25(OH)D (ng/mL) | 16.03 ± 8.91 | 24.58 ± 13.04 * | 0.032 |
| DBP (mg/L) | 372.28 ± 144.83 | 409.82 ± 119.30 | 0.150 |
| Free 25(OH)D (pg/mL) | 2.99 ± 1.76 | 4.35 ± 2.53 * | 0.043 |
| Bioavailable 25(OH)D (ng/mL) | 1.41 ± 0.81 | 2.09 ± 0.90 * | 0.017 |
| n (%) | Free 25(OH)D < 2 pg/mL (n = 10) | Free 25(OH)D > 2 pg/mL (n = 41) | p Values | Bioavailable 25(OH)D < 2 ng/mL (n = 33) | Bioavailable 25(OH)D > 2 ng/mL (n = 18) | p Values | |
|---|---|---|---|---|---|---|---|
| Genotype distribution | AA | 0 (0) | 1 (%100) | 0.309 | 0 (0) | 1 (100) | N/A |
| AC | 8 (%30) | 19 (%70) * | 0.028 | 19 (70.4) | 8 (29.6) | 0.276 | |
| CC | 2 (%9) | 21 (%91) * | 0.038 | 15 (65.2) | 8 (34.8) | 0.421 | |
| Allele frequency | A | 0.80 | 0.49 | 0.194 | 0.56 | 0.53 | 0.495 |
| C | 1.00 | 0.98 | 1.0 | 0.94 | |||
| Phenotypes | X ± SD | |||
|---|---|---|---|---|
| Total 25(OH)D (ng/mL) | DBP (mg/L) | Free 25(OH)D (pg/mL) | Bioavailable 25(OH)D (ng/mL) | |
| Gc1s/1s (n = 16) | 27.69 ± 13.7 | 391.8 ± 91.1 | 5.42 ± 2.35 | 2.44 ± 0.99 |
| Gc1s/1f (n = 7) | 17.27 ± 4.3 | 423.4 ± 163.8 | 3.28 ± 0.93 | 1.55 ± 0.65 |
| Gc1s/2 (n = 18) | 16.48 ± 9.4 * | 361.7 ± 88.6 | 2.56 ± 1.46 ** | 1.34 ± 0.79 ** |
| Gc1f/1f (n = 3) | 24.26 ± 13.0 | 417.0 ± 153.8 | 4.92 ± 2.62 | 2.05 ± 1.17 |
| Gc1f/2 (n = 7) | 13.10 ± 6.5 * | 303.3 ± 46.1 | 2.48 ± 1.26 ** | 1.19 ± 0.59 ** |
| p value | 0.016 | 0.202 | 0.003 | 0.005 |
| Phenotype | Total 25(OH)D < 20 ng/mL (n = 26) | Total 25(OH)D ≥ 20 ng/mL (n = 25) | p Values | Free 25(OH)D < 2 pg/mL (n = 9) | Free 25(OH)D ≥ 2 pg/mL (n = 42) | p Values | Bioavailable 25(OH)D < 2 ng/mL (n = 33) | Bioavailable 25(OH)D ≥ 2 ng/mL (n = 18) | p Values |
|---|---|---|---|---|---|---|---|---|---|
| Gc1s/1s | 5 (31.25) | 11 (68.75) * | 0.028 | 1 (6.25) | 15 (93.75) | 0.074 | 7 (43.75) | 9 (56.25)* | 0.017 |
| Gc1s/1f | 5 (71.43) | 2 (28.57) | 0.122 | 0 (0) | 7 (100) | 0.204 | 6 (85.72) | 1 (14.28) | 0.105 |
| Gc1s/2 | 10 (55.56) | 8 (44.44) | 0.315 | 6 (33.34) | 12 (66.66) * | 0.015 | 13 (61.54) | 5 (38.46) | 0.203 |
| Gc1f/1f | 2 (66.67) | 1 (33.33) | 0.288 | 0 (0) | 3 (100) | 0.204 | 1 (33.34) | 2 (66.66) | 0.121 |
| Gc1f/2 | 4 (57.15) | 3 (42.85) | 0.363 | 2 (28.58) | 5 (71.42) | 0.207 | 6 (83.34) | 1 (16.66) | 0.105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karcıoğlu Batur, L.; Özaydın, A.; Maviş, M.E.; Gürsu, G.G.; Harbige, L.; Hekim, N. Vitamin-D Binding Protein Gene Polymorphisms and Serum 25-Hydroxyvitamin-D in a Turkish Population. Metabolites 2021, 11, 696. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100696
Karcıoğlu Batur L, Özaydın A, Maviş ME, Gürsu GG, Harbige L, Hekim N. Vitamin-D Binding Protein Gene Polymorphisms and Serum 25-Hydroxyvitamin-D in a Turkish Population. Metabolites. 2021; 11(10):696. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100696
Chicago/Turabian StyleKarcıoğlu Batur, Lutfiye, Ahmet Özaydın, Murat Emrah Maviş, Gökçe Göksu Gürsu, Laurence Harbige, and Nezih Hekim. 2021. "Vitamin-D Binding Protein Gene Polymorphisms and Serum 25-Hydroxyvitamin-D in a Turkish Population" Metabolites 11, no. 10: 696. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11100696