
Translational Application of Fluorescent Molecular Probes for the Detection of Reactive Oxygen and Nitrogen Species Associated with Intestinal Reperfusion Injury

 ,
,  , and
, and
Abstract
:1. Introduction
2. Discussion
2.1. Historical Context
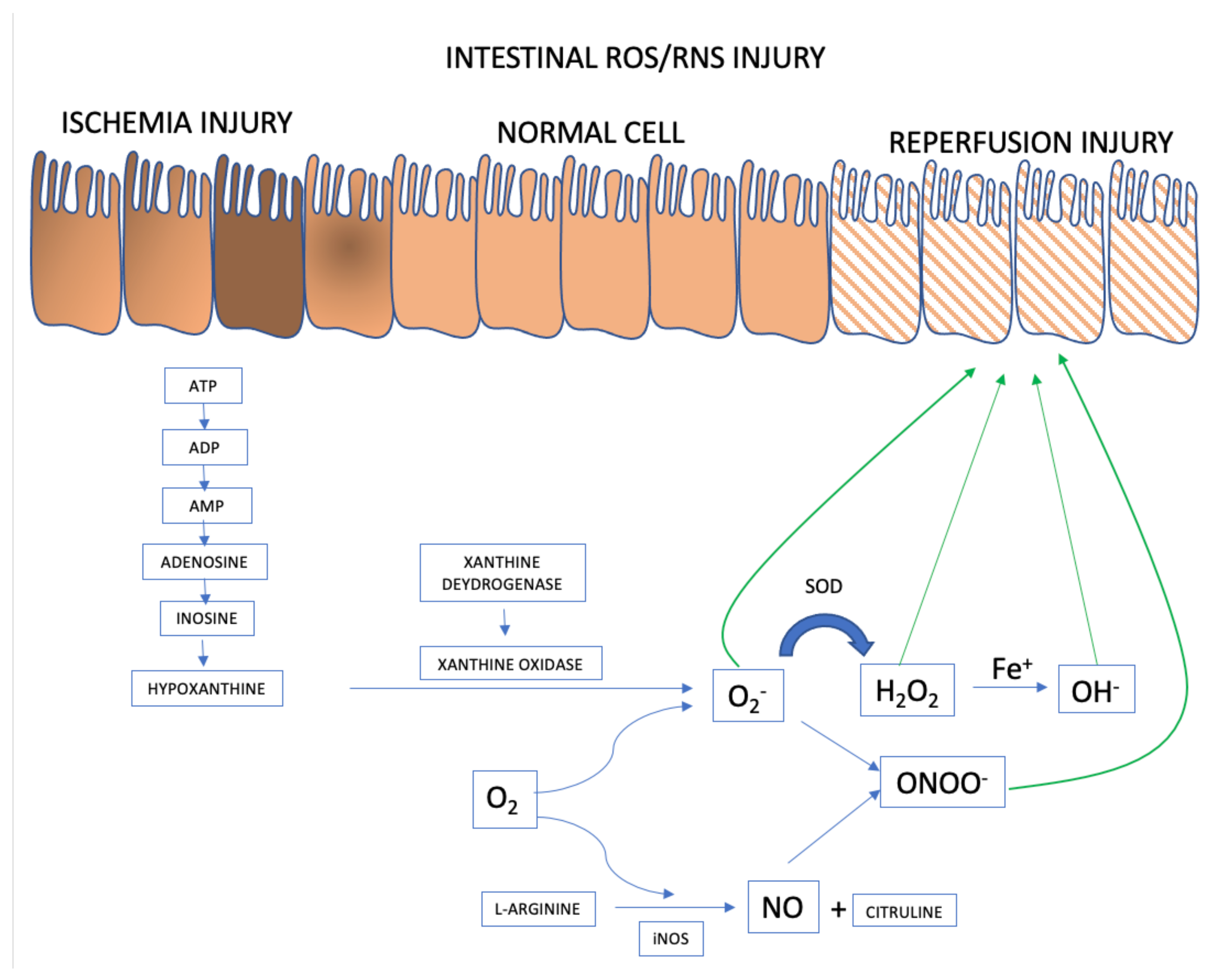
2.2. Free Radical Synthesis and the Pathophysiology of Ischemia/Reperfusion
Intestinal Epithelial Cells
2.3. Molecular Probe Fundamentals
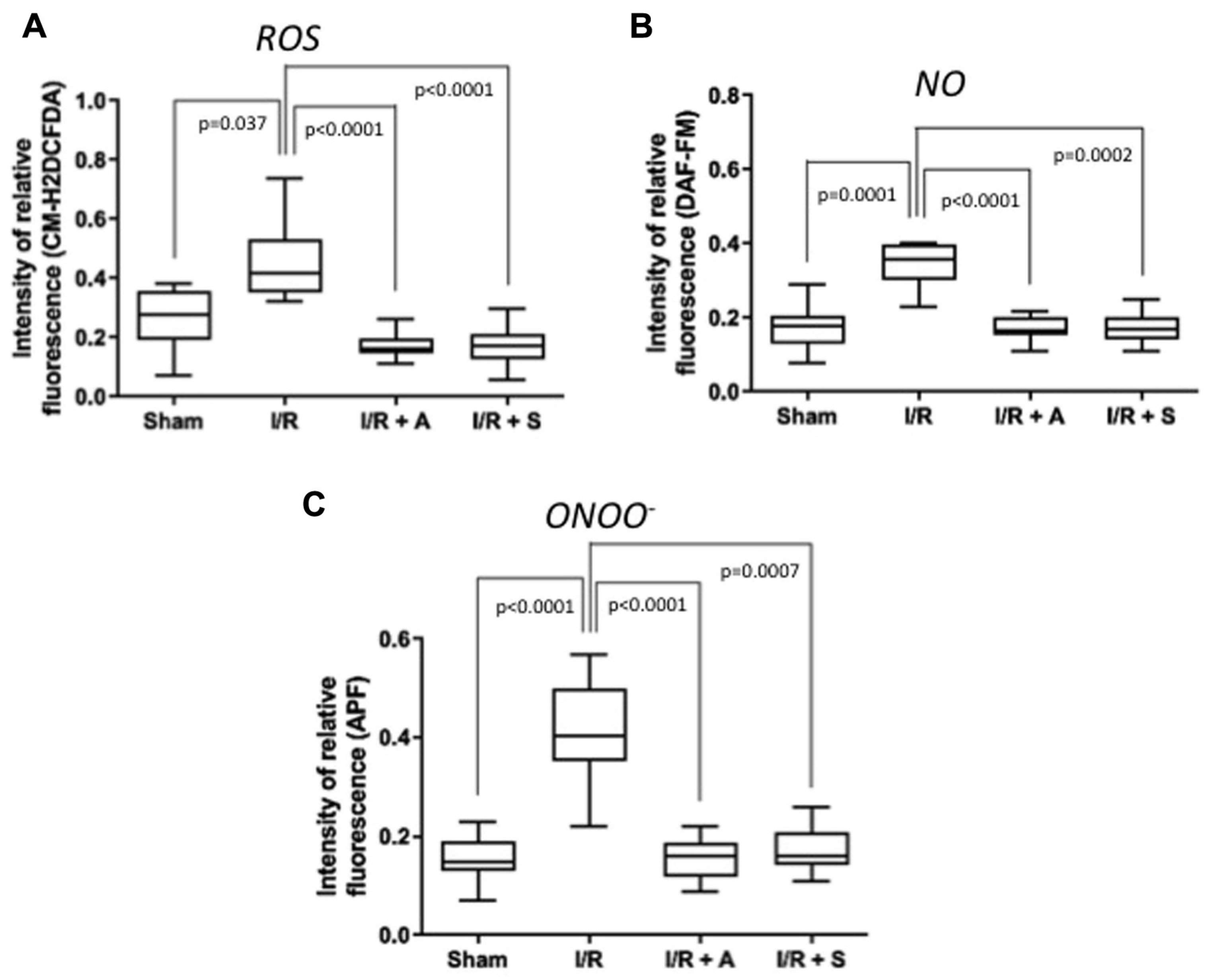
2.4. Translational Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuhn, F.; Schiergens, T.S.; Klar, E. Acute mesenteric ischemia. Visc. Med. 2020, 36, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, B.A. Acute gut ischemia. Surg. Clin. North Am. 2018, 98, 995–1004. [Google Scholar] [CrossRef]
- Karkkainen, J.M. Acute mesenteric ischemia: A challenge for the acute care surgeon. Scand. J. Surg. 2021, 14574969211007590. [Google Scholar] [CrossRef]
- Gnanapandithan, K.; Feuerstadt, P. Review article: Mesenteric ischemia. Curr. Gastroenterol. Rep. 2020, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Memet, O.; Zhang, L.; Shen, J. Serological biomarkers for acute mesenteric ischemia. Ann. Transl. Med. 2019, 7, 394. [Google Scholar] [CrossRef]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.-G. The role of reactive oxygen species (ROS) in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhatt, H.S.; Behr, S.C.; Miracle, A.; Wang, Z.J.; Yeh, B.M. Radiological evaluation of bowel ischemia. Radiol. Clin. N. Am. 2015, 53, 1241–1254. [Google Scholar] [CrossRef] [Green Version]
- Peoc’h, K.; Corcos, O. Biomarkers for acute mesenteric ischemia diagnosis: State of the art and perspectives. Ann. Biol. Clin. 2019, 77, 415–421. [Google Scholar] [CrossRef]
- Haber, F.; Willstätter, R. Unpaarigkeit und Radikalketten im Reaktionsmechanismus organischer und enzymatischer Vorgänge. Ber. Der Dtsch. Chem. Ges. A B Ser. 1931, 64, 2844–2856. [Google Scholar] [CrossRef]
- Haber, F.; Weiss, J.; Pope, W.J. The catalytic decomposition of hydrogen peroxide by iron salts. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1934, 147, 332–351. [Google Scholar]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- De Flora, S.; Grassi, C.; Carati, L. Attenuation of influenza-like symptomatology and improvement of cell-mediated immunity with long-term N-acetylcysteine treatment. Eur. Respir. J. 1997, 10, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Acosta, S. Mesenteric ischemia. Curr. Opin. Crit. Care. 2015, 21, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Karkkainen, J.M.; Acosta, S. Acute mesenteric ischemia (part I)—Incidence, etiologies, and how to improve early diagnosis. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 15–25. [Google Scholar] [CrossRef]
- Karkkainen, J.M.; Acosta, S. Acute mesenteric ischemia (Part II)—Vascular and endovascular surgical approaches. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 27–38. [Google Scholar] [CrossRef]
- Prakash, V.S.; Marin, M.; Faries, P.L. Acute and chronic ischemic disorders of the small bowel. Curr. Gastroenterol. Rep. 2019, 21, 27. [Google Scholar] [CrossRef]
- Granger, D.N. Intestinal microcirculation and transmucosal fluid transport. Am. J. Physiol. 1981, 240, G343–G349. [Google Scholar] [CrossRef]
- Granger, D.N.; Hollwarth, M.E.; Parks, D.A. Ischemia-reperfusion injury: Role of oxygen-derived free radicals. Acta Physiol. Scand. Suppl. 1986, 548, 47–63. [Google Scholar]
- Kajino-Sakamoto, R.; Omori, E.; Nighot, P.K.; Blikslager, A.T.; Matsumoto, K.; Ninomiya-Tsuji, J. TGF-beta-activated kinase 1 signaling maintains intestinal integrity by preventing accumulation of reactive oxygen species in the intestinal epithelium. J. Immunol. 2010, 185, 4729–4737. [Google Scholar] [CrossRef] [Green Version]
- VanDeinse, W.H.; Zawacki, J.K.; Phillips, D. Treatment of acute mesenteric ischemia by percutaneous transluminal angioplasty. Gastroenterology 1986, 91, 475–478. [Google Scholar] [CrossRef]
- Beaulieu, R.J.; Arnaoutakis, K.D.; Abularrage, C.J.; Efron, D.T.; Schneider, E.; Black, J.H., 3rd. Comparison of open and endovascular treatment of acute mesenteric ischemia. J. Vasc. Surg. 2014, 59, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Xie, Y.; Li, J. A colorimetric and fluorescent probe based on rhodamine B for detection of Fe(3+) and Cu(2+) ions. J. Fluoresc. 2019, 29, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Nadatani, Y.; Watanabe, T.; Shimada, S.; Otani, K.; Tanigawa, T.; Fujiwara, Y. Microbiome and intestinal ischemia/reperfusion injury. J. Clin. Biochem. Nutr. 2018, 63, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Bertoni, S.; Ballabeni, V.; Barocelli, E.; Tognolini, M. Mesenteric ischemia-reperfusion: An overview of preclinical drug strategies. Drug Discov. Today 2018, 23, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Mester, A.; Magyar, Z.; Sogor, V.; Tanczos, B.; Stark, Y.; Cherniavsky, K.; Laszlo, B.; Katalin, P.; Norbert, N. Intestinal ischemia-reperfusion leads to early systemic micro-rheological and multiorgan microcirculatory alterations in the rat. Clin. Hemorheol. Microcirc. 2018, 68, 35–44. [Google Scholar] [CrossRef]
- Papezikova, I.; Lojek, A.; Cizova, H.; Ciz, M. Alterations in plasma antioxidants during reperfusion of the ischemic small intestine in rats. Res. Vet. Sci. 2006, 81, 140–147. [Google Scholar] [CrossRef]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Ischemia/reperfusion. Compr. Physiol. 2016, 7, 113–170. [Google Scholar] [PubMed] [Green Version]
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox. Biol. 2015, 6, 524–551. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Wang, H.; Fang, S.; Xu, C. Roles of endoplasmic reticulum stress and autophagy on H2O2 induced oxidative stress injury in HepG2 cells. Mol. Med. Rep. 2018, 18, 4163–4174. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Morris, H.; Cronin, M.T. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef] [Green Version]
- Liskova, A.; Samec, M.; Koklesova, L.; Kudela, E.; Kubatka, P.; Golubnitschaja, O. Mitochondriopathies as a clue to systemic disorders-analytical tools and mitigating measures in context of predictive, preventive, and personalized (3P) medicine. Int. J. Mol. Sci. 2021, 22, 2007. [Google Scholar] [CrossRef]
- Battelli, M.G.; Polito, L.; Bolognesi, A. Xanthine oxidoreductase in atherosclerosis pathogenesis: Not only oxidative stress. Atherosclerosis 2014, 237, 562–567. [Google Scholar] [CrossRef] [Green Version]
- Kleinert, H.; Schwarz, P.M.; Forstermann, U. Regulation of the expression of inducible nitric oxide synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef]
- Valenca, S.S.; Pimenta, W.A.; Rueff-Barroso, C.R.; Ferreira, T.S.; Resende, A.C.; Moura, R.S.; Porto, L.C. Involvement of nitric oxide in acute lung inflammation induced by cigarette smoke in the mouse. Nitric Oxide 2009, 20, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Pires, K.M.; Lanzetti, M.; Rueff-Barroso, C.R.; Castro, P.; Abrahao, A.; Koatz, V.L.; Valença, S.S.; Porto, L.C. Oxidative damage in alveolar macrophages exposed to cigarette smoke extract and participation of nitric oxide in redox balance. Toxicol. In Vitro 2012, 26, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Nesi, R.T.; Barroso, M.V.; Souza Muniz, V.; de Arantes, A.C.; Martins, M.A.; Brito Gitirana, L.; Neves, J.S.; Benjamim, C.F.; Lanzetti, M.; Valenca, S.S. Pharmacological modulation of reactive oxygen species (ROS) improves the airway hyperresponsiveness by shifting the Th1 response in allergic inflammation induced by ovalbumin. Free Radic Res. 2017, 51, 708–722. [Google Scholar] [CrossRef]
- Valenca, S.S.; Rueff-Barroso, C.R.; Pimenta, W.A.; Melo, A.C.; Nesi, R.T.; Silva, M.A.; Porto, L.C. L-NAME and L-arginine differentially ameliorate cigarette smoke-induced emphysema in mice. Pulm. Pharmacol. Ther. 2011, 24, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Lanzetti, M.; da Costa, C.A.; Nesi, R.T.; Barroso, M.V.; Martins, V.; Victoni, T.; Lagente, V.; Pires, K.M.P.; Silva, P.M.R.e.; Resende, A.C.; et al. Oxidative stress and nitrosative stress are involved in different stages of proteolytic pulmonary emphysema. Free Radic. Biol. Med. 2012, 53, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Forstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Moller, M.N.; Rios, N.; Trujillo, M.; Radi, R.; Denicola, A.; Alvarez, B. Detection and quantification of nitric oxide-derived oxidants in biological systems. J. Biol. Chem. 2019, 294, 14776–14802. [Google Scholar] [CrossRef] [Green Version]
- Barzilai, A.; Yamamoto, K. DNA damage responses to oxidative stress. DNA Repair. 2004, 3, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.C.; Huang, C.S.; Ming, Y.C.; Chu, S.M.; Chao, H.C. Calcitonin gene-related peptide downregulates expression of inducible nitride oxide synthase and caspase-3 after intestinal ischemia-reperfusion injury in rats. Pediatr. Neonatol. 2016, 57, 474–479. [Google Scholar] [CrossRef] [Green Version]
- Eiserich, J.P.; Patel, R.P.; O’Donnell, V.B. Pathophysiology of nitric oxide and related species: Free radical reactions and modification of biomolecules. Mol. Aspects Med. 1998, 19, 221–357. [Google Scholar] [CrossRef]
- Adams, L.; Franco, M.C.; Estevez, A.G. Reactive nitrogen species in cellular signaling. Exp. Biol. Med. 2015, 240, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Harris, C.M.; Sanders, S.A.; Massey, V. Role of the flavin midpoint potential and NAD binding in determining NAD versus oxygen reactivity of xanthine oxidoreductase. J. Biol. Chem. 1999, 274, 4561–4569. [Google Scholar] [CrossRef] [Green Version]
- Laursen, J.B.; Somers, M.; Kurz, S.; McCann, L.; Warnholtz, A.; Freeman, B.A.; Tarpey, M.; Fukai, T.; Harrison, D.G. Endothelial regulation of vasomotion in apoE-deficient mice: Implications for interactions between peroxynitrite and tetrahydrobiopterin. Circulation 2001, 103, 1282–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocker, R.; Perrella, M.A. Heme oxygenase-1: A novel drug target for atherosclerotic diseases? Circulation 2006, 114, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wu, Z.; Riwanto, M.; Gao, S.; Levison, B.S.; Gu, X.; Fu, X.; Wagner, M.A.; Besler, C.; Gerstenecker, G.; et al. Myeloperoxidase, paraoxonase-1, and HDL form a functional ternary complex. J. Clin. Investig. 2013, 123, 3815–3828. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L. Remodeling of resistance arteries in essential hypertension and effects of antihypertensive treatment. Am. J. Hypertens. 2004, 17, 1192–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauseef, W.M. Assembly of the phagocyte NADPH oxidase. Histochem. Cell Biol. 2004, 122, 277–291. [Google Scholar] [CrossRef]
- Lee, M.Y.; Griendling, K.K. Redox signaling, vascular function, and hypertension. Antioxid. Redox Signal. 2008, 10, 1045–1059. [Google Scholar] [CrossRef]
- Liao, G.; Chen, S.; Cao, H.; Wang, W.; Gao, Q. Review: Acute superior mesenteric artery embolism: A vascular emergency cannot be ignored by physicians. Medicine 2019, 98, e14446. [Google Scholar] [CrossRef]
- Singh, M.; Long, B.; Koyfman, A. Mesenteric ischemia: A deadly miss. Emerg. Med. Clin. North Am. 2017, 35, 879–888. [Google Scholar] [CrossRef]
- Li, R.; Jia, Z.; Zhu, H. Regulation of Nrf2 signaling. React. Oxyg. Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Duanghathaipornsuk, S.; Farrell, E.J.; Alba-Rubio, A.C.; Zelenay, P.; Kim, D.S. Detection technologies for reactive oxygen species: Fluorescence and electrochemical methods and their applications. Biosensors 2021, 11, 30. [Google Scholar] [CrossRef]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and methods to measure oxidative stress in clinical samples: Research applications in the cancer field. Oxid. Med. Cell Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Wang, L.; Carroll, S.L.; Chen, J.; Wang, M.C.; Wang, J. Challenges and opportunities for small-molecule fluorescent probes in redox biology applications. Antioxid. Redox Signal. 2018, 29, 518–540. [Google Scholar] [CrossRef] [PubMed]
- Hempel, S.L.; Buettner, G.R.; O’Malley, Y.Q.; Wessels, D.A.; Flaherty, D.M. Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: Comparison with 2′,7′-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 1999, 27, 146–159. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, J., II; Ischiropoulss, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: Challenges and limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Han, H.; Zhang, H.; Song, S.; Shuang, S.; Dong, C. Boronate based sensitive fluorescent probe for the detection of endogenous peroxynitrite in living cells. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 243, 118683. [Google Scholar] [CrossRef]
- Deshwal, S.; Antonucci, S.; Kaludercic, N.; di Lisa, F. Measurement of mitochondrial rOS Formation. Methods Mol. Biol. 2018, 1782, 403–418. [Google Scholar] [PubMed]
- Fluorescence imaging of oxidative stress in live cells. BioProbes J. Cell Biol. Appl. 2011, 65, 10.
- Schenk, B.; Fulda, S. Reactive oxygen species regulate Smac mimetic/TNFalpha-induced necroptotic signaling and cell death. Oncogene 2015, 34, 5796–5806. [Google Scholar] [CrossRef]
- Kageyama, S.; Hirao, H.; Nakamura, K.; Ke, B.; Zhang, M.; Ito, T.; Aziz, A.; Oncel, D.; Kaldas, F.M.; Bussutil, R.W.; et al. Recipient HO-1 inducibility is essential for posttransplant hepatic HO-1 expression and graft protection: From bench-to-bedside. Am. J. Transplant. 2019, 19, 356–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peshavariya, H.M.; Dusting, G.J.; Selemidis, S. Analysis of dihydroethidium fluorescence for the detection of intracellular and extracellular superoxide produced by NADPH oxidase. Free Radic. Res. 2007, 41, 699–712. [Google Scholar] [CrossRef]
- Zhao, H.; Joseph, J.; Fales, H.M.; Sokoloski, E.A.; Levine, R.L.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc. Natl. Acad. Sci. USA 2005, 102, 5727–5732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal 2014, 20, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, T. Bioimaging probes for reactive oxygen species and reactive nitrogen species. J. Clin. Biochem. Nutr. 2009, 45, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Cohn, C.A.; Simon, S.R.; Schoonen, M.A. Comparison of fluorescence-based techniques for the quantification of particle-induced hydroxyl radicals. Part Fibre Toxicol. 2008, 5, 2. [Google Scholar] [CrossRef]
- Wiederschain, G.Y. The molecular probes handbook. A guide to fluorescent probes and labeling technologies. Biochemistry 2011, 76, 1276. [Google Scholar] [CrossRef] [Green Version]
- Childs, E.W.; Udobi, K.F.; Wood, J.G.; Hunter, F.A.; Smalley, D.M.; Cheung, L.Y. In vivo visualization of reactive oxidants and leukocyte-endothelial adherence following hemorrhagic shock. Shock 2002, 18, 423–427. [Google Scholar] [CrossRef]
- Tang, Y.; Shen, J.; Zhang, F.; Yang, F.Y.; Liu, M. Human serum albumin attenuates global cerebral ischemia/reperfusion-induced brain injury in a Wnt/beta-Catenin/ROS signaling-dependent manner in rats. Biomed. Pharmacother. 2019, 115, 108871. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Costantino, G.; Mazzon, E.; Micali, A.; de Sarro, A.; Caputi, A.P. Beneficial effects of melatonin in a rat model of splanchnic artery occlusion and reperfusion. J. Pineal Res. 2000, 28, 52–63. [Google Scholar] [CrossRef]
- Yan, X.T.; Cheng, X.L.; He, X.H.; Zheng, W.Z.; Xiao-Fang, Y.; Hu, C. The HO-1-expressing bone mesenchymal stem cells protects intestine from ischemia and reperfusion injury. BMC Gastroenterol. 2019, 19, 124. [Google Scholar] [CrossRef] [Green Version]
- Nagira, M.; Tomita, M.; Mizuno, S.; Kumata, M.; Ayabe, T.; Hayashi, M. Ischemia/reperfusion injury in the monolayers of human intestinal epithelial cell line caco-2 and its recovery by antioxidants. Drug Metab. Pharmacokinet. 2006, 21, 230–237. [Google Scholar] [CrossRef]
- Sampaio de Holanda, G.; dos Santos Valenca, S.; Maran Carra, A.; Lopes Lichtenberger, R.C.; Franco, O.B.; Ribeiro, B.E.; Rosas, S.L.P.; Santana, P.T.; Castelo-Branco, M.T.L.; de Souza, H.S.F.; et al. Sulforaphane and albumin attenuate experimental intestinal ischemia-reperfusion injury. J. Surg. Res. 2021, 262, 212–223. [Google Scholar] [CrossRef]
- Fang, Y.; Dehaen, W. Fluorescent probes for selective recognition of hypobromous acid: Achievements and future perspectives. Molecules 2021, 26, 363. [Google Scholar] [CrossRef]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical relevance of biomarkers of oxidative stress. Antioxid. Redox Signal 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Probes | Reactive Species | Chemical Structure |
| DHR123 | Hydrogen peroxide Hypochlorous acid Peroxynitrite anion |  |
| CM-H2DCFDA | Hydrogen peroxide Hydroxyl radical Peroxynitrite anion Peroxyl radical |  |
| CellRox | Hydrogen peroxide Hydroxyl radical Nitric oxide Peroxynitrite anion Superoxide anion | |
| Dihydroethidium | Superoxide anion |  |
| MitoSox | Superoxide anion |  |
| DAF-FM | Nitric oxide | |
| APF and HPF | Hypochlorous acid Peroxynitrite anion |  |
| Hydroxyl radical | ||
| Boronate | Hydrogen peroxide Peroxynitrite anion |  |
| Amplex red | Hydrogen peroxide |  |
| Author | Fluorescent Probe | Results | Graphics |
|---|---|---|---|
| Childs EW, et al. [73] | Dihydrorhodamine 123 i.v. and observes in vivo mesenteric endothelium | Reactive oxygen species production in the mesenteric microvascular endothelium, attributed to hemorrhagic shock and reperfusion injury, after resuscitation, and mediated by the administration of a platelet activating factor antagonist |  Effect of platelet activating factor (anti-LFA-1_ and WEB 2086) on leukocyte adherence ROS given 10 min prior to the shock period versus the hemorrhagic shock alone group. * p < 0.05 compared with the hemmorhagic shock alone group. |
| Tang Y, et al. [74] | DCFH-DA (2′, 7′-dichlorodihydrofluorescein diacetate) used in fresh tissue homogenates | Human albumin intravenous administration, in ROS attenuation, in a global cerebral ischemia reperfusion model by Wnt/β-Catenin pathway signaling |  Effect of human albumin treatment on oxidative stress following global cerebral ischemia/reperfusion, (p < 0.05) in contrast to the Global Cerebral Ischemia/Reperfusion (GCI/R) group, * p < 0.05 in contrast to GCI/R group, # p < 0.05, in contrast to the GCI/R+Human Serum Albumin group. |
| Cuzzocrea S, et al. [75] | Dihydrorodamine 123 i.v. plasma analysed | Melatonin infusion attenuated the reperfusion injury produced by splanchnic artery occlusion |  Plasma peroxynitrite production assessed by oxidation of dihydrorhodamine 123 to rhodamine. Peroxynitrite production in the Splancnic Arterial Oclusion (SAO)-shocked rats was significantly increased versus sham group. Melatonin-treated rats show a significant reduction of the SAO-induced elevation of the plasma peroxynitrite production. * p < 0.01 versus vehicle. ° p < 0.01 versus SAO. |
| Yan XT, et al. [76] | DCFH-DA used in homogenized intestinal tissue | Heme Oxygenase-1-expressing Bone Marrow Steam Cell after intestinal I/R performed by temporary occlusion of the superior mesenteric artery |  Bone Marrow Steam Cell/HemeOxygenase-1 (BMSC/HO-1) attenuated production of ROS in intestine and serum. Levels of ROS in intestine at 24 h of reperfusion were significantly higher than those in sham group and decreased after treatment of BMSC/HO-1. ** p < 0.01 vs. Sham; # p < 0.05 vs. I/R; + p < 0.05 vs. BMSC. |
| Nagira M, et al. [77] | Rhodamine 123 | Lutein effects In vitro ischemia reperfusion injury, using monolayers of human colon cancer intestinal epithelial cell line |  The effects of lutein and biliverdin on rhodamine 123 permeability in the apical to basal direction in cell monolayers. * p < 0.05 and ** p < 0.01 vs. lipid peroxidation inducer. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Holanda, G.S.; dos Santos Valença, S.; Carra, A.M.; Lichtenberger, R.C.L.; de Castilho, B.; Franco, O.B.; de Moraes, J.A.; Schanaider, A. Translational Application of Fluorescent Molecular Probes for the Detection of Reactive Oxygen and Nitrogen Species Associated with Intestinal Reperfusion Injury. Metabolites 2021, 11, 802. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120802
de Holanda GS, dos Santos Valença S, Carra AM, Lichtenberger RCL, de Castilho B, Franco OB, de Moraes JA, Schanaider A. Translational Application of Fluorescent Molecular Probes for the Detection of Reactive Oxygen and Nitrogen Species Associated with Intestinal Reperfusion Injury. Metabolites. 2021; 11(12):802. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120802
Chicago/Turabian Stylede Holanda, Gustavo Sampaio, Samuel dos Santos Valença, Amabile Maran Carra, Renata Cristina Lopes Lichtenberger, Bianca de Castilho, Olavo Borges Franco, João Alfredo de Moraes, and Alberto Schanaider. 2021. "Translational Application of Fluorescent Molecular Probes for the Detection of Reactive Oxygen and Nitrogen Species Associated with Intestinal Reperfusion Injury" Metabolites 11, no. 12: 802. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120802