
Dietary Fat and Protein Intake in Relation to Plasma Sphingolipids as Determined by a Large-Scale Lipidomic Analysis

 , , , ,
, , , ,
Abstract
:
1. Introduction
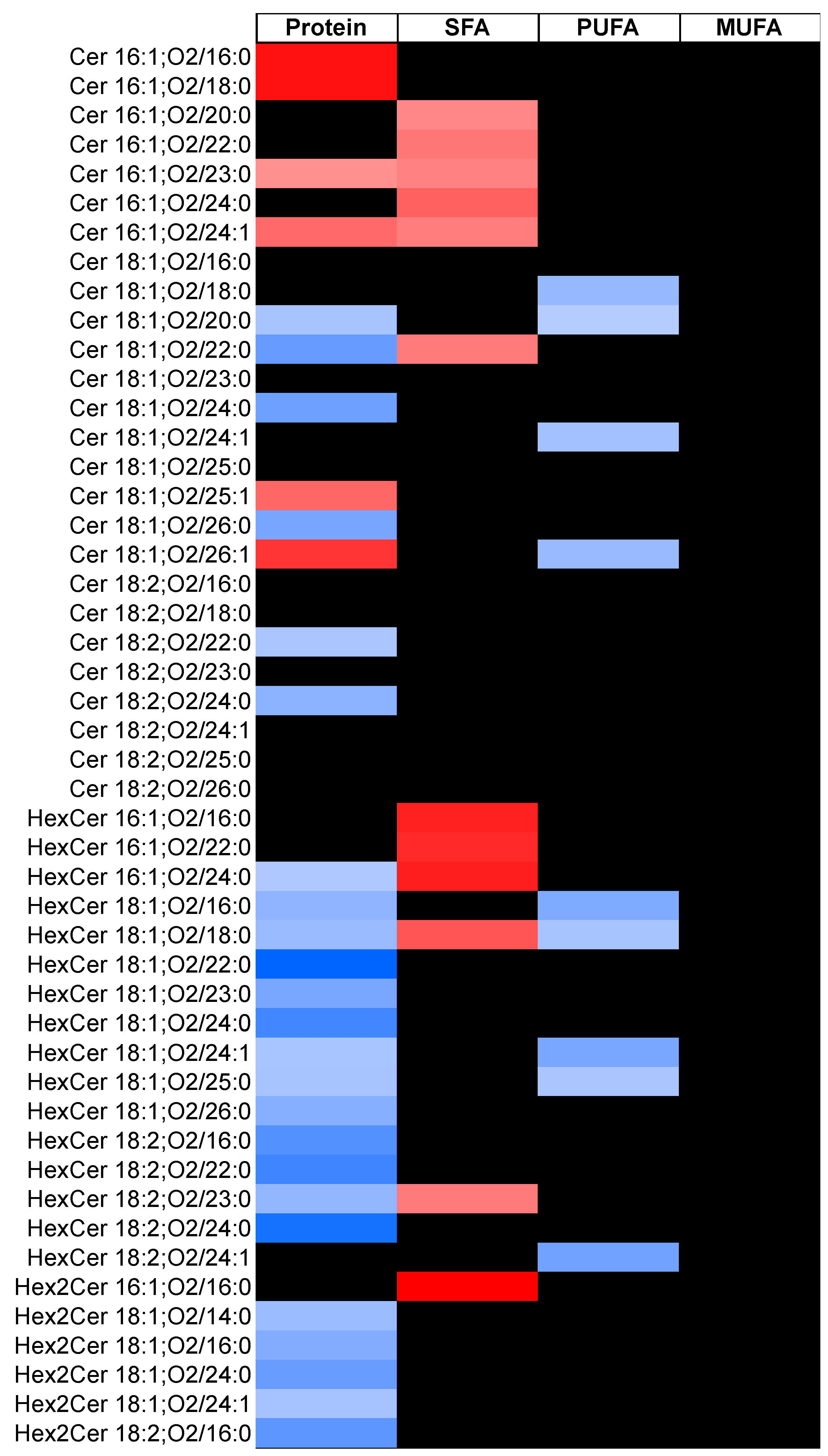
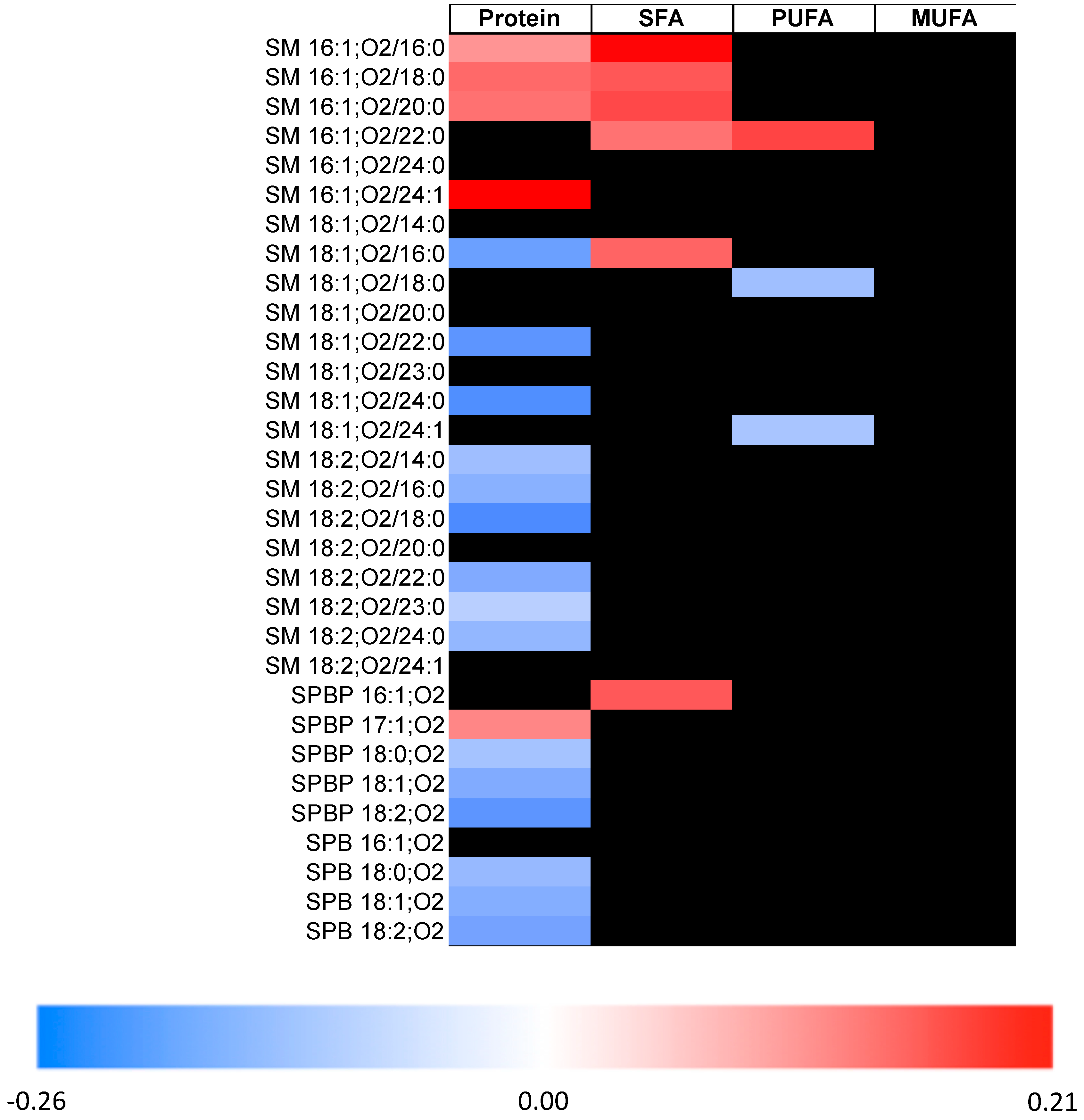
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Lipid Extraction and Quantification
4.3. Assessment of Diet and Covariates
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Chew, W.S.; Torta, F.; Ji, S.S.; Choi, H.; Begum, H.; Sim, X.; Khoo, C.M.; Khoo, E.Y.H.; Ong, W.Y.; Van Dam, R.M.; et al. Large-scale lipidomics identifies associations between plasma sphingolipids and T2DM incidence. JCI Insight 2019, 4, e126925. [Google Scholar] [CrossRef]
- Seah, J.Y.H.; Chew, W.S.; Torta, F.; Khoo, C.M.; Wenk, M.R.; Herr, D.R.; Choi, H.; Tai, E.S.; van Dam, R.M. Plasma sphingolipids and risk of cardiovascular diseases: A large-scale lipidomic analysis. Metabolomics 2020, 16, 89. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, R.N.; Fretts, A.M.; Sitlani, C.M.; Biggs, M.L.; Mukamal, K.; King, I.B.; Song, X.L.; Djousse, L.; Siscovick, D.S.; McKnight, B.; et al. Plasma phospholipid very-long-chain saturated fatty acids and incident diabetes in older adults: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2015, 101, 1047–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigruener, A.; Kleber, M.E.; Heimerl, S.; Liebisch, G.; Schmitz, G.; Maerz, W. Glycerophospholipid and Sphingolipid Species and Mortality: The Ludwigshafen Risk and Cardiovascular Health (LURIC) Study. PLoS ONE 2014, 9, e85724. [Google Scholar] [CrossRef] [Green Version]
- Mundra, P.A.; Barlow, C.K.; Nestel, P.J.; Barnes, E.H.; Kirby, A.; Thompson, P.; Sullivan, D.R.; Alshehry, Z.H.; Mellett, N.A.; Huynh, K.; et al. Large-scale plasma lipidomic profiling identifies lipids that predict cardiovascular events in secondary prevention. JCI Insight 2018, 3, e121326. [Google Scholar] [CrossRef]
- Wang, D.D.; Toledo, E.; Hruby, A.; Rosner, B.A.; Willett, W.C.; Sun, Q.; Razquin, C.; Zheng, Y.; Ruiz-Canela, M.; Guasch-Ferre, M.; et al. Plasma Ceramides, Mediterranean Diet, and Incident Cardiovascular Disease in the PREDIMED Trial ( Prevencion con Dieta Mediterranea). Circulation 2017, 135, 2028–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laaksonen, R.; Ekroos, K.; Sysi-Aho, M.; Hilvo, M.; Vihervaara, T.; Kauhanen, D.; Suoniemi, M.; Hurme, R.; Marz, W.; Scharnagl, H.; et al. Plasma ceramides predict cardiovascular death in patients with stable coronary artery disease and acute coronary syndromes beyond LDL-cholesterol. Eur. Heart J. 2016, 37, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K. Mechanisms of disease—Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [Green Version]
- Pickup, J.C. Inflammation and activated innate immunity in the pathogenesis of type 2 diabetes. Diabetes Care 2004, 27, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Munoz, A.; Presa, N.; Gomez-Larrauri, A.; Rivera, I.G.; Trueba, M.; Ordonez, M. Control of inflammatory responses by ceramide, sphingosine 1-phosphate and ceramide 1-phosphate. Prog. Lipid Res. 2016, 61, 51–62. [Google Scholar] [CrossRef]
- Gao, D.; Pararasa, C.; Dunston, C.R.; Bailey, C.J.; Griffiths, H.R. Palmitate promotes monocyte atherogenicity via de novo ceramide synthesis. Free Radic. Biol. Med. 2012, 53, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Yang, X.Y.; Xing, S.S.; Bian, F.; Yao, W.J.; Bai, X.L.; Zheng, T.; Wu, G.J.; Jin, S. Endogenous Ceramide Contributes to the Transcytosis of oxLDL across Endothelial Cells and Promotes Its Subendothelial Retention in Vascular Wall. Oxidative Med. Cell. Longev. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Et Biophys. Acta-Mol. Cell Biol. Lipids 2002, 1585, 114–125. [Google Scholar] [CrossRef]
- Boslem, E.; Meikle, P.J.; Biden, T.J. Roles of ceramide and sphingolipids in pancreatic beta-cell function and dysfunction. Islets 2012, 4, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Tomita, T. Apoptosis in pancreatic beta-islet cells in Type 2 diabetes. Bosn. J. Basic Med Sci. 2016, 16, 162–179. [Google Scholar]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. Sphingolipids as Signaling and Regulatory Molecules; Chalfant, C., DelPoeta, M., Eds.; Springer: New York, NY, USA, 2010; pp. 1–23. [Google Scholar]
- Luukkonen, P.K.; Sadevirta, S.; Zhou, Y.; Kayser, B.; Ali, A.; Ahonen, L.; Lallukka, S.; Pelloux, V.; Gaggini, M.; Jian, C.; et al. Saturated Fat Is More Metabolically Harmful for the Human Liver Than Unsaturated Fat or Simple Sugars. Diabetes Care 2018, 41, 1732–1739. [Google Scholar] [CrossRef] [Green Version]
- Rosqvist, F.; Kullberg, J.; Ståhlman, M.; Cedernaes, J.; Heurling, K.; Johansson, H.E.; Iggman, D.; Wilking, H.; Larsson, A.; Eriksson, O.; et al. Overeating Saturated Fat Promotes Fatty Liver and Ceramides Compared With Polyunsaturated Fat: A Randomized Trial. J. Clin. Endocrinol. Metab. 2019, 104, 6207–6219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankinen, M.; Schwab, U.; Kolehmainen, M.; Paananen, J.; Nygren, H.; Seppanen-Laakso, T.; Poutanen, K.; Hyotylainen, T.; Riserus, U.; Savolainen, M.J.; et al. A Healthy Nordic Diet Alters the Plasma Lipidomic Profile in Adults with Features of Metabolic Syndrome in a Multicenter Randomized Dietary Intervention. J. Nutr. 2016, 146, 662–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankinen, M.; Schwab, U.; Erkkila, A.; Seppanen-Laakso, T.; Hannila, M.L.; Mussalo, H.; Lehto, S.; Uusitupa, M.; Gylling, H.; Oresic, M. Fatty Fish Intake Decreases Lipids Related to Inflammation and Insulin Signaling-A Lipidomics Approach. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [Green Version]
- Lankinen, M.; Schwab, U.; Kolehmainen, M.; Paananen, J.; Poutanen, K.; Mykkanen, H.; Seppanen-Laakso, T.; Gylling, H.; Uusitupa, M.; Oresic, M. Whole Grain Products, Fish and Bilberries Alter Glucose and Lipid Metabolism in a Randomized, Controlled Trial: The Sysdimet Study. PLoS ONE 2011, 6, e22646. [Google Scholar] [CrossRef] [Green Version]
- Ottestad, I.; Hassani, S.; Borge, G.I.; Kohler, A.; Vogt, G.; Hyotylainen, T.; Oresic, M.; Bronner, K.W.; Holven, K.B.; Ulven, S.M.; et al. Fish Oil Supplementation Alters the Plasma Lipidomic Profile and Increases Long-Chain PUFAs of Phospholipids and Triglycerides in Healthy Subjects. PLoS ONE 2012, 7, e42550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornemann, T.; Penno, A.; Rutti, M.F.; Ernst, D.; Kivrak-Pfiffner, F.; Rohrer, L.; von Eckardstein, A. The SPTLC3 Subunit of Serine Palmitoyltransferase Generates Short Chain Sphingoid Bases. J. Biol. Chem. 2009, 284, 26322–26330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachnio-Zabielska, A.; Baranowski, M.; Zabielski, P.; Gorski, J. Effect of High Fat Diet Enriched With Unsaturated and Diet Rich in Saturated Fatty Acids on Sphingolipid Metabolism in Rat Skeletal Muscle. J. Cell. Physiol. 2010, 225, 786–791. [Google Scholar] [CrossRef]
- Russo, S.B.; Tidhar, R.; Futerman, A.H.; Cowart, L.A. Myristate-derived d16:0 Sphingolipids Constitute a Cardiac Sphingolipid Pool with Distinct Synthetic Routes and Functional Properties. J. Biol. Chem. 2013, 288, 13397–13409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesper, H.; Schmelz, E.M.; Nikolova-Karakashian, M.N.; Dillehay, D.L.; Lynch, D.V.; Merrill, A.H. Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J. Nutr. 1999, 129, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Norris, G.H.; Blesso, C.N. Dietary and Endogenous Sphingolipid Metabolism in Chronic Inflammation. Nutrients 2017, 9, 1180. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, A.; Duan, R.D. Absorption and lipoprotein transport of sphingomyelin. J. Lipid Res. 2006, 47, 154–171. [Google Scholar] [CrossRef] [Green Version]
- Zendzian-Piotrowska, M.; Baranowski, M.; Zabielski, P.; Gorski, J. Effects of Pioglitazone and high-fat diet on ceramide metabolism in rat skeletal muscles. J. Physiol. Pharmacol. 2006, 57, 101–114. [Google Scholar]
- Helge, J.W.; Tobin, L.; Drachmann, T.; Hellgren, L.I.; Dela, F.; Galbo, H. Muscle ceramide content is similar after 3 weeks’ consumption of fat or carbohydrate diet in a crossover design in patients with type 2 diabetes. Eur. J. Appl. Physiol. 2012, 112, 911–918. [Google Scholar] [CrossRef]
- Deurenberg-Yap, M.; Li, T.; Tan, W.L.; van Staveren, W.A.; Deurenberg, P. Validation of a semiquantitative food frequency questionnaire for estimation of intakes of energy, fats and cholesterol among Singaporeans. Asia Pac. J. Clin. Nutr. 2000, 9, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Burla, B.; Muralidharan, S.; Wenk, M.R.; Torta, F. Sphingolipid Analysis in Clinical Research. Methods Mol. Biol. 2018, 1730, 135–162. [Google Scholar] [PubMed]
- Narayanaswamy, P.; Shinde, S.; Sulc, R.; Kraut, R.; Staples, G.; Thiam, C.H.; Grimm, R.; Sellergren, B.; Torta, F.; Wenk, M.R. Lipidomic “Deep Profiling”: An Enhanced Workflow to Reveal New Molecular Species of Signaling Lipids. Anal. Chem. 2014, 86, 3043–3047. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.H.X.; Tan, L.W.L.; Sim, X.; Tai, E.S.; Lee, J.J.; Chia, K.S.; van Dam, R.M. Cohort Profile: The Singapore Multi-Ethnic Cohort (MEC) study. Int. J. Epidemiol. 2018, 47, 699j. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, J.; Jiang, X.C.; Tabas, I.; Tall, A.; Shea, S. Plasma sphingomyelin and subclinical atherosclerosis: Findings from the multi-ethnic study of atherosclerosis. Am. J. Epidemiol. 2006, 163, 903–912. [Google Scholar] [CrossRef]
- Begum, H.; Li, B.W.; Shui, G.H.; Cazenave-Gassiot, A.; Soong, R.; Ong, R.T.H.; Little, P.; Teo, Y.Y.; Wenk, M.R. Discovering and validating between-subject variations in plasma lipids in healthy subjects. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gika, H.G.; Theodoridis, G.A.; Wingate, J.E.; Wilson, I.D. Within-day reproducibility of an HPLC-MS-Based method for metabonomic analysis: Application to human urine. J. Proteome Res. 2007, 6, 3291–3303. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Koefeler, H.; et al. Update on LIPID MAPS Classification, Nomenclature and Shorthand Notation for MS-derived Lipid Structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Nang, E.E.K.; Ngunjiri, S.A.G.; Wu, Y.; Salim, A.; Tai, E.S.; Lee, J.; Van Dam, R.M. Validity of the international physical activity questionnaire and the Singapore prospective study program physical activity questionnaire in a multiethnic urban Asian population. BMC Med. Res. Methodol. 2011, 11, 141. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B-Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Age at interview (y) | 48.6 ± 12.0 |
| Male (n (%)) | 1345 (47.0) |
| Cigarette smoking (n (%)) | |
| Never-smoker | 2295 (80.2) |
| Ex-smoker | 247 (8.6) |
| Current smoker | 318 (11.1) |
| Alcohol intake (n (%)) | |
| Never or hardly ever | 2210 (77.3) |
| Mild to moderate | 220 (7.7) |
| Moderate to heavy | 159 (5.6) |
| Heavy | 271 (9.5) |
| Physical activity (MET-h/week) 2 | 108 ± 82 |
| Energy intake (kcal/d) | 2168 ± 920 |
| Nutrient intake (% energy) | |
| Protein | 15.9 ± 2.2 |
| Total fat | 30.5 ± 5.6 |
| Saturated fat | 11.4 ± 2.8 |
| Polyunsaturated fat | 6.4 ± 2.7 |
| Monounsaturated fat | 10.8 ± 2.7 |
| Body mass index (kg/m2) | 22.9 ± 3.8 |
| HDL cholesterol (mmol/L) 3 | 1.48 ± 0.37 |
| LDL cholesterol (mmol/L) 3 | 3.21 ± 0.81 |
| Triglycerides (mmol/L) 3 | 1.31 ± 0.86 |
| Protein | Saturated Fat | Polyunsaturated Fat | Monounsaturated Fat | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β2 | 95% CI | β | 95% CI | β | 95% CI | β | 95% CI | |||||
| Ceramides | −0.11 | −0.18 | −0.04 | 0.07 | 0.01 | 0.14 | −0.01 | −0.07 | 0.04 | −0.02 | −0.09 | 0.05 |
| Short-chain (C14) | NA | |||||||||||
| Long-chain (C16–C18) | 0.00 | −0.08 | 0.08 | 0.02 | −0.06 | 0.10 | −0.11 | −0.18 | −0.04 | 0.00 | −0.09 | 0.08 |
| Very long chain (C20–C26) | −0.11 | −0.18 | −0.05 | 0.08 | 0.01 | 0.14 | −0.01 | −0.07 | 0.05 | −0.02 | −0.09 | 0.05 |
| 16:1;O2 | 0.06 | −0.01 | 0.13 | 0.13 | 0.06 | 0.20 | 0.05 | −0.01 | 0.12 | −0.10 | −0.17 | −0.03 |
| 18:1;O2 | −0.12 | −0.19 | −0.06 | 0.07 | 0.00 | 0.14 | −0.03 | −0.08 | 0.03 | 0.00 | −0.07 | 0.07 |
| 18:2;O2 | −0.10 | −0.18 | −0.03 | 0.04 | −0.04 | 0.11 | 0.02 | −0.05 | 0.08 | −0.06 | −0.13 | 0.02 |
| HexCer | −0.20 | −0.28 | −0.12 | 0.07 | −0.01 | 0.15 | −0.09 | −0.16 | −0.02 | 0.03 | −0.06 | 0.11 |
| Short-chain (C14) | NA | |||||||||||
| Long-chain (C16–C18) | −0.14 | −0.22 | −0.05 | 0.05 | −0.03 | 0.14 | −0.14 | −0.22 | −0.07 | 0.03 | −0.05 | 0.12 |
| Very long chain (C20–C26) | −0.21 | −0.29 | −0.13 | 0.07 | −0.01 | 0.15 | −0.08 | −0.15 | −0.02 | 0.02 | −0.06 | 0.11 |
| 16:1;O2 | −0.06 | −0.15 | 0.02 | 0.19 | 0.11 | 0.27 | 0.02 | −0.05 | 0.09 | −0.06 | −0.14 | 0.03 |
| 18:1;O2 | −0.20 | −0.29 | −0.12 | 0.06 | −0.02 | 0.14 | −0.10 | −0.16 | −0.03 | 0.03 | −0.05 | 0.11 |
| 18:2;O2 | −0.20 | −0.29 | −0.12 | 0.05 | −0.03 | 0.13 | −0.08 | −0.15 | −0.01 | 0.00 | −0.08 | 0.08 |
| Hex2Cer | −0.15 | −0.24 | −0.07 | 0.06 | −0.02 | 0.15 | 0.01 | −0.06 | 0.08 | 0.01 | −0.07 | 0.10 |
| Short-chain (C14) | −0.11 | −0.20 | −0.03 | 0.06 | −0.02 | 0.14 | 0.05 | −0.02 | 0.12 | −0.04 | −0.13 | 0.04 |
| Long-chain (C16–C18) | −0.15 | −0.23 | −0.06 | 0.05 | −0.03 | 0.13 | 0.01 | −0.06 | 0.08 | 0.01 | −0.07 | 0.10 |
| Very long chain (C20–C26) | −0.15 | −0.23 | −0.06 | 0.09 | 0.01 | 0.18 | −0.01 | −0.08 | 0.06 | 0.03 | −0.06 | 0.11 |
| 16:1;O2 | 0.00 | −0.09 | 0.08 | 0.21 | 0.12 | 0.30 | 0.06 | −0.01 | 0.14 | −0.09 | −0.18 | −0.01 |
| 18:1;O2 | −0.15 | −0.24 | −0.07 | 0.05 | −0.03 | 0.14 | 0.00 | −0.07 | 0.07 | 0.02 | −0.06 | 0.10 |
| 18:2;O2 | −0.18 | −0.26 | −0.10 | 0.09 | 0.00 | 0.17 | 0.00 | −0.07 | 0.07 | 0.00 | −0.08 | 0.08 |
| SM | −0.12 | −0.19 | −0.05 | 0.09 | 0.02 | 0.16 | −0.03 | −0.09 | 0.02 | 0.00 | −0.07 | 0.07 |
| Short-chain (C14) | −0.08 | −0.15 | −0.01 | 0.05 | −0.02 | 0.12 | 0.01 | −0.05 | 0.07 | −0.07 | −0.14 | 0.00 |
| Long-chain (C16–C18) | −0.16 | −0.23 | −0.08 | 0.14 | 0.07 | 0.22 | −0.08 | −0.14 | −0.02 | −0.01 | −0.08 | 0.06 |
| Very long chain (C20–C26) | −0.08 | −0.15 | −0.01 | 0.03 | −0.03 | 0.10 | 0.01 | −0.05 | 0.06 | 0.02 | −0.05 | 0.08 |
| 16:1;O2 | 0.13 | 0.05 | 0.21 | 0.16 | 0.08 | 0.24 | 0.08 | 0.01 | 0.14 | −0.10 | −0.18 | −0.02 |
| 18:1;O2 | −0.15 | −0.22 | −0.08 | 0.08 | 0.01 | 0.15 | −0.06 | −0.12 | 0.00 | 0.03 | −0.04 | 0.10 |
| 18:2;O2 | −0.12 | −0.19 | −0.05 | 0.03 | −0.04 | 0.09 | −0.01 | −0.07 | 0.05 | −0.01 | −0.08 | 0.06 |
| SPB | −0.14 | −0.23 | −0.05 | 0.04 | −0.05 | 0.13 | 0.00 | −0.07 | 0.08 | 0.04 | −0.05 | 0.13 |
| 16:1;O2 | −0.01 | −0.10 | 0.07 | 0.07 | −0.02 | 0.16 | 0.04 | −0.04 | 0.11 | −0.01 | −0.10 | 0.08 |
| 18:1;O2 | −0.14 | −0.23 | −0.05 | 0.03 | −0.06 | 0.12 | −0.01 | −0.09 | 0.07 | 0.06 | −0.04 | 0.15 |
| 18:2;O2 | −0.16 | −0.25 | −0.07 | 0.03 | −0.06 | 0.12 | 0.01 | −0.07 | 0.09 | 0.04 | −0.05 | 0.14 |
| SPBP | −0.15 | −0.24 | −0.06 | 0.00 | −0.09 | 0.09 | 0.03 | −0.05 | 0.10 | 0.09 | 0.00 | 0.18 |
| 16:1;O2 | 0.00 | −0.09 | 0.08 | 0.14 | 0.05 | 0.23 | 0.08 | 0.00 | 0.15 | −0.02 | −0.11 | 0.07 |
| 18:1;O2 | −0.15 | −0.24 | −0.06 | 0.00 | −0.09 | 0.09 | 0.02 | −0.06 | 0.10 | 0.09 | 0.00 | 0.18 |
| 18:2;O2 | −0.18 | −0.27 | −0.09 | −0.03 | −0.12 | 0.06 | −0.01 | −0.08 | 0.07 | 0.11 | 0.02 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seah, J.Y.H.; Chew, W.S.; Torta, F.; Khoo, C.M.; Wenk, M.R.; Herr, D.R.; Tai, E.S.; van Dam, R.M. Dietary Fat and Protein Intake in Relation to Plasma Sphingolipids as Determined by a Large-Scale Lipidomic Analysis. Metabolites 2021, 11, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020093
Seah JYH, Chew WS, Torta F, Khoo CM, Wenk MR, Herr DR, Tai ES, van Dam RM. Dietary Fat and Protein Intake in Relation to Plasma Sphingolipids as Determined by a Large-Scale Lipidomic Analysis. Metabolites. 2021; 11(2):93. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020093
Chicago/Turabian StyleSeah, Jowy Yi Hoong, Wee Siong Chew, Federico Torta, Chin Meng Khoo, Markus R. Wenk, Deron R. Herr, E. Shyong Tai, and Rob M. van Dam. 2021. "Dietary Fat and Protein Intake in Relation to Plasma Sphingolipids as Determined by a Large-Scale Lipidomic Analysis" Metabolites 11, no. 2: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11020093