Distinct Calcium Binding and Structural Properties of Two Centrin Isoforms from Toxoplasma gondii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Production
2.2. Nuclear Magnetic Resonance Spectroscopy
2.3. Circular Dichroism Spectroscopy
2.4. Limited Proteolysis
2.5. Size Exclusion Chromatography
2.6. Fluorescence Spectroscopy
2.7. Isothermal Titration Calorimetry
2.8. Turbidity Measurements
2.9. Dynamic Light Scattering
3. Results
3.1. Sequence Analysis and Protein Production
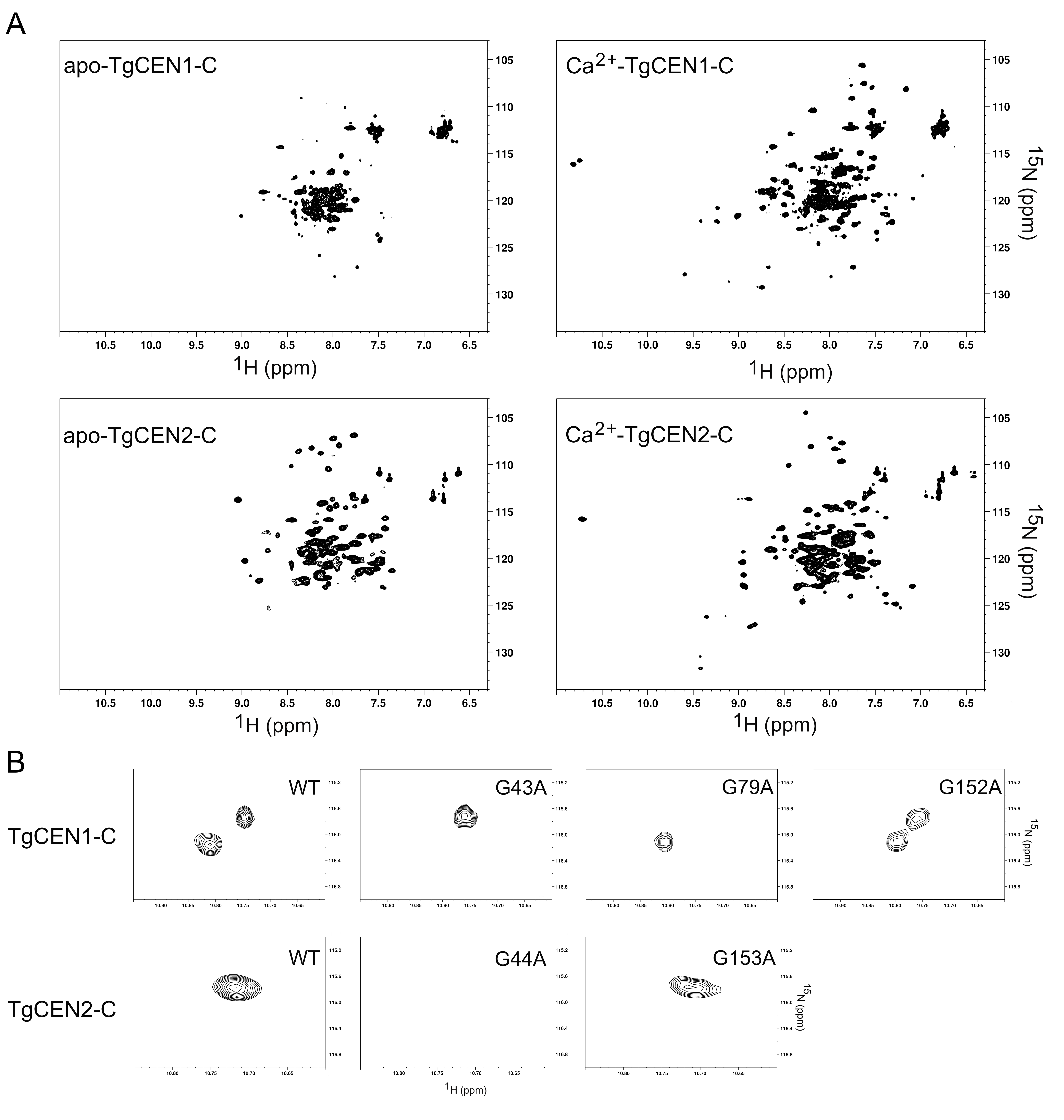
3.2. NMR Spectroscopy
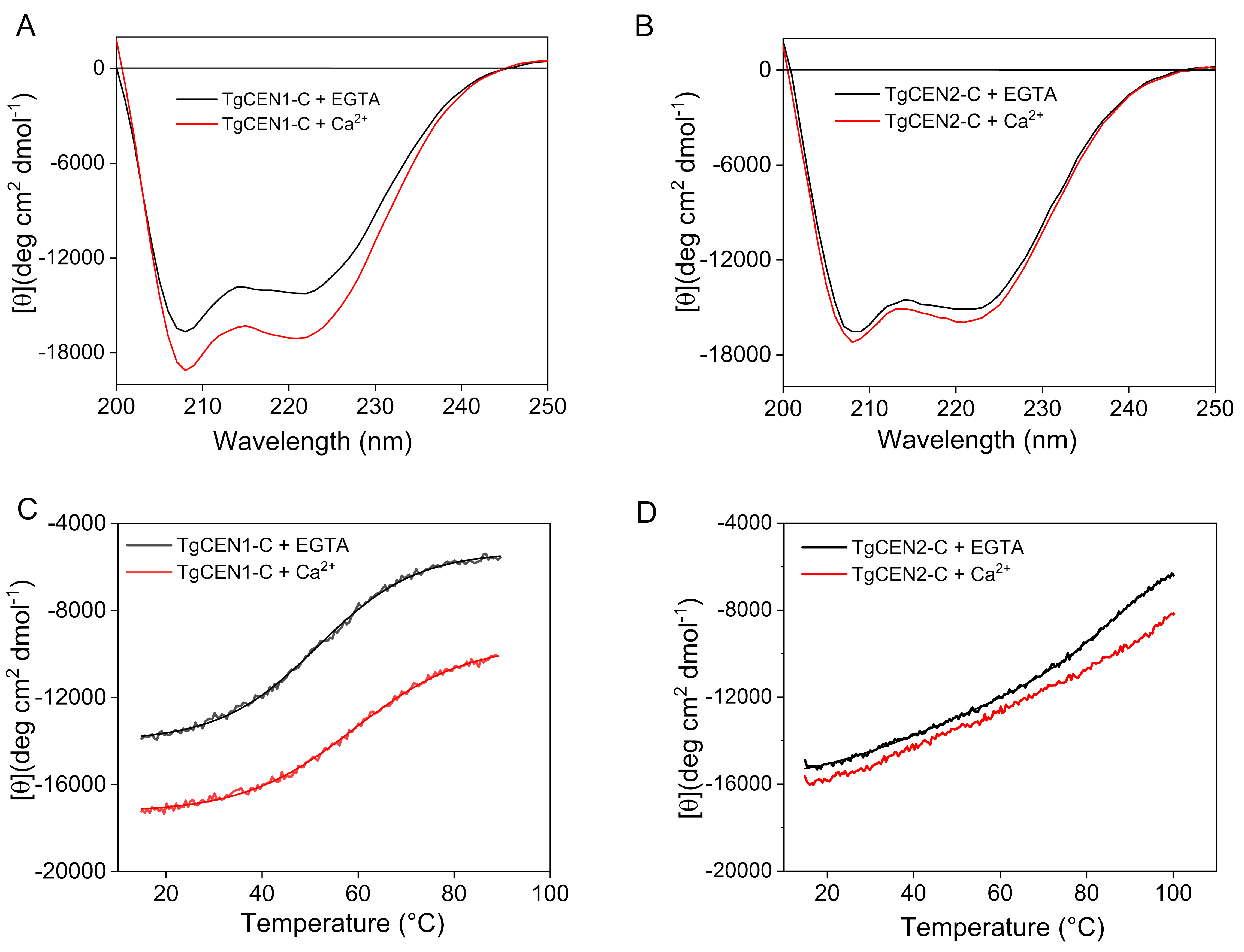
3.3. Far-UV CD Spectroscopy
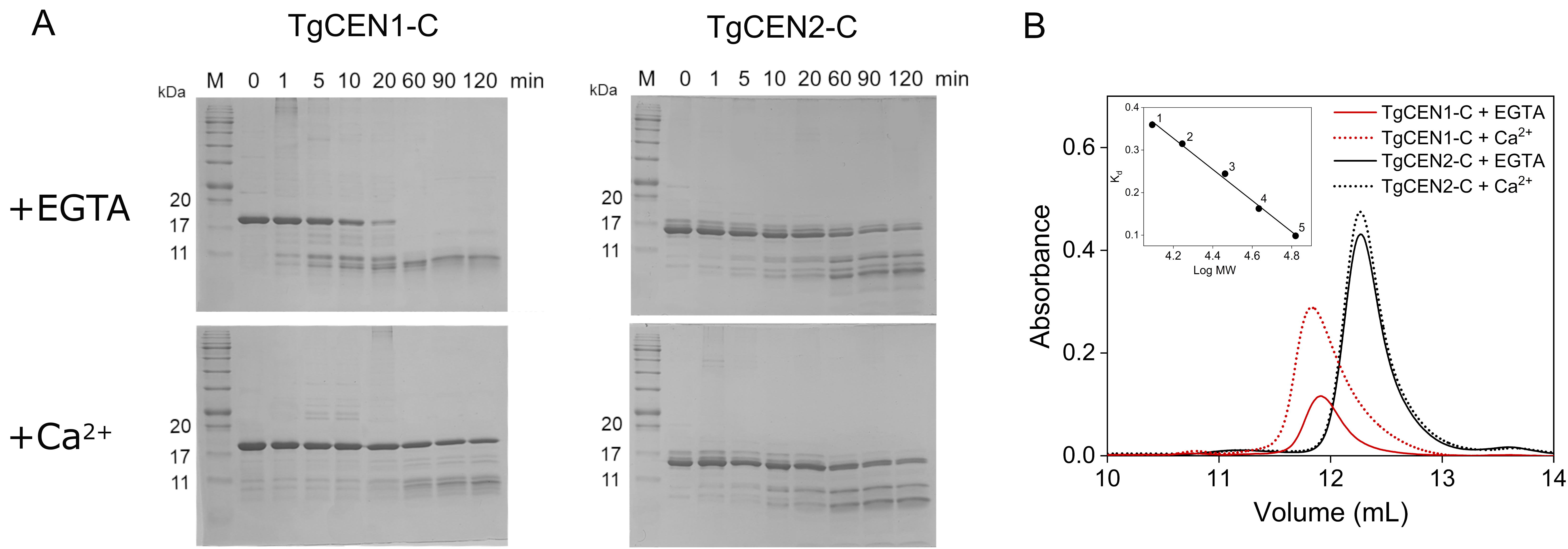
3.4. Limited Proteolysis and Size Exclusion Chromatography
3.5. Interaction of TgCEN1-C and TgCEN2-C with ANS
3.6. Energetics of Ca2+ Binding
3.7. Turbidity Measurements
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paoletti, A.; Moudjou, M.; Paintrand, M.; Salisbury, J.L.; Bornens, M. Most of centrin in animal cells is not centrosome-associated and centrosomal centrin is confined to the distal lumen of centrioles. J. Cell Sci. 1996, 109, 3089–3102. [Google Scholar] [PubMed]
- Nishi, R.; Okuda, Y.; Watanabe, E.; Mori, T.; Iwai, S.; Masutani, C.; Sugasawa, K.; Hanaoka, F. Centrin 2 stimulates nucleotide excision repair by interacting with xeroderma pigmentosum group C protein. Mol. Cell. Biol. 2005, 25, 5664–5674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Flury, S.; Kalck, V.; Hohn, B.; Molinier, J. CENTRIN2 interacts with the Arabidopsis homolog of the human XPC protein (AtRAD4) and contributes to efficient synthesis-dependent repair of bulky DNA lesions. Plant. Mol. Biol. 2006, 61, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miron, S.; Durand, D.; Chilom, C.; Perez, J.; Craescu, C.T. Binding of calcium, magnesium, and target peptides to Cdc31, the centrin of yeast Saccharomyces cerevisiae. Biochemistry 2011, 50, 6409–6422. [Google Scholar] [CrossRef]
- Jani, D.; Lutz, S.; Marshall, N.J.; Fischer, T.; Kohler, A.; Ellisdon, A.M.; Hurt, E.; Stewart, M. Sus1, Cdc31, and the Sac3 CID region form a conserved interaction platform that promotes nuclear pore association and mRNA export. Mol. Cell 2009, 33, 727–737. [Google Scholar] [CrossRef]
- La Verde, V.; Trande, M.; D’Onofrio, M.; Dominici, P.; Astegno, A. Binding of calcium and target peptide to calmodulin-like protein CML19, the centrin 2 of Arabidopsis thaliana. Int. J. Biol. Macromol. 2018, 108, 1289–1299. [Google Scholar] [CrossRef]
- Pedretti, M.; Conter, C.; Dominici, P.; Astegno, A. SAC3B is a target of CML19, the centrin 2 of Arabidopsis thaliana. Biochem. J. 2020, 477, 173–189. [Google Scholar] [CrossRef]
- Baum, P.; Furlong, C.; Byers, B. Yeast gene required for spindle pole body duplication: Homology of its product with Ca2+-binding proteins. Proc. Natl. Acad. Sci. USA 1986, 83, 5512–5516. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Mengersen, A.; Lee, V.D. Molecular cloning of cDNA for caltractin, a basal body-associated Ca2+-binding protein: Homology in its protein sequence with calmodulin and the yeast CDC31 gene product. J. Cell Biol. 1988, 107, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Madeddu, L.; Klotz, C.; Le Caer, J.P.; Beisson, J. Characterization of centrin genes in Paramecium. Eur. J. Biochem. 1996, 238, 121–128. [Google Scholar] [CrossRef]
- Del Vecchio, A.J.; Harper, J.D.I.; Vaughn, K.C.; Baron, A.T.; Salisbury, J.L.; Overall, R.L. Centrin homologues in higher plants are prominently associated with the developing cell plate. Protoplasma 1997, 196, 224–234. [Google Scholar] [CrossRef]
- Hu, H.; Chazin, W.J. Unique features in the C-terminal domain provide caltractin with target specificity. J. Mol. Biol. 2003, 330, 473–484. [Google Scholar] [CrossRef]
- Cox, J.A.; Tirone, F.; Durussel, I.; Firanescu, C.; Blouquit, Y.; Duchambon, P.; Craescu, C.T. Calcium and magnesium binding to human centrin 3 and interaction with target peptides. Biochemistry 2005, 44, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Sheehan, J.H.; Chazin, W.J. The Mode of Action of Centrin: Binding of Ca2+ and a peptide fragment of Kar1p to the C-terminal domain. J. Biol. Chem. 2004, 279, 50895–50903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, F.; Ye, K.; Zhang, J.; Liao, S.; Zhang, X.; Xu, C.; Tu, X. Solution structure of TbCentrin4 from Trypanosoma brucei and its interactions with Ca2+ and other centrins. Biochem. J. 2018, 475, 3763–3778. [Google Scholar] [CrossRef] [PubMed]
- Gifford, J.L.; Walsh, M.P.; Vogel, H.J. Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem. J. 2007, 405, 199–221. [Google Scholar] [CrossRef]
- Carlström, G.; Chazin, W.J. Two-dimensional 1H Nuclear Magnetic Resonance Studies of the Half-saturated (Ca2+)1 State of Calbindin D9k: Further Implications for the Molecular Basis of Cooperative Ca2+ binding. J. Mol. Biol. 1993, 231, 415–430. [Google Scholar] [CrossRef]
- Nagamune, K.; Sibley, L.D. Comparative genomic and phylogenetic analyses of calcium ATPases and calcium-regulated proteins in the apicomplexa. Mol. Biol. Evol. 2006, 23, 1613–1627. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Johnson, J.; Florens, L.; Fraunholz, M.; Suravajjala, S.; DiLullo, C.; Yates, J.; Roos, D.S.; Murray, J.M. Cytoskeletal components of an invasion machine—The apical complex of Toxoplasma gondii. PLoS Pathog. 2006, 2, e13. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, J.; Hu, K.; He, C.Y.; Pelletier, L.; Roos, D.S.; Warren, G. Golgi and centrosome cycles in Toxoplasma gondii. Mol. Biochem. Parasitol. 2006, 145, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Lentini, G.; Dubois, D.J.; Maco, B.; Soldati-Favre, D.; Frénal, K. The roles of Centrin 2 and Dynein Light Chain 8a in apical secretory organelles discharge of Toxoplasma gondii. Traffic 2019, 20, 583–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, J.M.; Liu, J.; Wetzel, L.A.; Hu, K. Centrin2 from the human parasite Toxoplasma gondii is required for its invasion and intracellular replication. J. Cell Sci. 2019, 132, jcs228791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar]
- Vallone, R.; La Verde, V.; D’Onofrio, M.; Giorgetti, A.; Dominici, P.; Astegno, A. Metal binding affinity and structural properties of calmodulin-like protein 14 from Arabidopsis thaliana. Protein Sci. 2016, 25, 1461–1471. [Google Scholar] [CrossRef] [Green Version]
- Astegno, A.; Giorgetti, A.; Allegrini, A.; Cellini, B.; Dominici, P. Characterization of C-S Lyase from C. diphtheriae: A possible target for new antimicrobial drugs. Biomed. Res. Int. 2013, 2013, 701536. [Google Scholar] [CrossRef] [Green Version]
- La Verde, V.; Dominici, P.; Astegno, A. Determination of Hydrodynamic Radius of Proteins by Size Exclusion Chromatography. BioProtocol 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Trande, M.; Pedretti, M.; Bonza, M.C.; Di Matteo, A.; D’Onofrio, M.; Dominici, P.; Astegno, A. Cation and peptide binding properties of CML7, a calmodulin-like protein from Arabidopsis thaliana. J. Inorg. Biochem. 2019, 199, 110796. [Google Scholar] [CrossRef]
- Astegno, A.; Maresi, E.; Marino, V.; Dominici, P.; Pedroni, M.; Piccinelli, F.; Dell’Orco, D. Structural plasticity of calmodulin on the surface of CaF2 nanoparticles preserves its biological function. Nanoscale 2014, 6, 15037–15047. [Google Scholar] [CrossRef]
- Marino, V.; Astegno, A.; Pedroni, M.; Piccinelli, F.; Dell’Orco, D. Nanodevice-induced conformational and functional changes in a prototypical calcium sensor protein. Nanoscale 2014, 6, 412–423. [Google Scholar] [CrossRef]
- Cuff, J.A.; Clamp, M.E.; Siddiqui, A.S.; Finlay, M.; Barton, G.J. JPred: A consensus secondary structure prediction server. Bioinformatics 1998, 14, 892–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, E.; Sigrist, C.J.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE signature matches and ProRule-associated functional and structural residues in proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef] [PubMed]
- Linse, S.; Forsen, S. Determinants that govern high-affinity calcium binding. Adv. Second Messenger Phosphoprot. Res. 1995, 30, 89–151. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, L.P.; Bujnicki, J.M. MetaDisorder: A meta-server for the prediction of intrinsic disorder in proteins. BMC Bioinform. 2012, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tourbez, M.; Firanescu, C.; Yang, A.; Unipan, L.; Duchambon, P.; Blouquit, Y.; Craescu, C.T. Calcium-dependent Self-assembly of Human Centrin 2. J. Biol. Chem. 2004, 279, 47672–47680. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Martin, S.R.; Newman, R.A.; Hamilton, S.L.; Shea, M.A.; Bayley, P.M.; Beckingham, K.M. Biochemical properties of V91G calmodulin: A calmodulin point mutation that deregulates muscle contraction in Drosophila. Protein Sci. 2004, 13, 3285–3297. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.-i.; Hayashi, H.; Chino, M. Detection of calmodulin and calmodulin-binding proteins in pure phloem sap of rice plants. Soil Sci. Plant Nutr. 2006, 52, 195–202. [Google Scholar] [CrossRef]
- Marmé, D.; Dieter, P. Role of Ca2+ and calmodulin in plants. In Calcium and Cell Function; Cheng, W.Y., Ed.; Academic Press: London, UK, 1983; Volume IV, pp. 263–311. [Google Scholar]
- Veeraraghavan, S.; Fagan, P.A.; Hu, H.; Lee, V.; Harper, J.F.; Huang, B.; Chazin, W.J. Structural independence of the two EF-hand domains of caltractin. J. Biol. Chem. 2002, 277, 28564–28571. [Google Scholar] [CrossRef] [Green Version]
- Astegno, A.; Bonza, M.C.; Vallone, R.; La Verde, V.; D’Onofrio, M.; Luoni, L.; Molesini, B.; Dominici, P. Arabidopsis calmodulin-like protein CML36 is a calcium (Ca2+) sensor that interacts with the plasma membrane Ca2+-ATPase isoform ACA8 and stimulates its activity. J. Biol. Chem. 2017, 292, 15049–15061. [Google Scholar] [CrossRef] [Green Version]
- Osawa, M.; Dace, A.; Tong, K.I.; Valiveti, A.; Ikura, M.; Ames, J.B. Mg2+ and Ca2+ differentially regulate DNA binding and dimerization of DREAM. J. Biol. Chem. 2005, 280, 18008–18014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamniuk, A.P.; Nguyen, L.T.; Hoang, T.T.; Vogel, H.J. Metal ion binding properties and conformational states of calcium- and integrin-binding protein. Biochemistry 2004, 43, 2558–2568. [Google Scholar] [CrossRef] [PubMed]
- Wingard, J.N.; Chan, J.; Bosanac, I.; Haeseleer, F.; Palczewski, K.; Ikura, M.; Ames, J.B. Structural analysis of Mg2+ and Ca2+ binding to CaBP1, a neuron-specific regulator of calcium channels. J. Biol. Chem. 2005, 280, 37461–37470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Mohan, P.M.; Chary, K.V. Magnesium promotes structural integrity and conformational switching action of a calcium sensor protein. Biochemistry 2007, 46, 3835–3845. [Google Scholar] [CrossRef] [PubMed]
- Gifford, J.L.; Jamshidiha, M.; Mo, J.; Ishida, H.; Vogel, H.J. Comparing the calcium binding abilities of two soybean calmodulins: Towards understanding the divergent nature of plant calmodulins. Plant. Cell 2013, 25, 4512–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Yan, J.; Feng, Y.; Liang, A.; Yang, B. Analysis of the role of Mg2+ on conformational change and target recognition by Ciliate Euplotes octocarinatus centrin. J. Photochem. Photobiol. B Biol. 2011, 105, 60–68. [Google Scholar] [CrossRef]
- Gagne, S.M.; Tsuda, S.; Li, M.X.; Chandra, M.; Smillie, L.B.; Sykes, B.D. Quantification of the calcium-induced secondary structural changes in the regulatory domain of troponin-C. Protein Sci. 1994, 3, 1961–1974. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Song, L.; Liang, A.; Yang, B. Characterization of self-assembly of Euplotes octocarinatus centrin. J. Photochem. Photobiol. B Biol. 2009, 95, 26–32. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, X.; Yang, B. Calcium-induced human centrin 1 self-assembly and double-regulating the binding with peptide R18-Sfi1p. Int. J. Biol. Macromol. 2019, 128, 314–323. [Google Scholar] [CrossRef]
- Arrizabalaga, G.; Boothroyd, J.C. Role of calcium during Toxoplasma gondii invasion and egress. Int. J. Parasitol. 2004, 34, 361–368. [Google Scholar] [CrossRef]
- Chang, L.; Dykes, E.J.; Li, J.; Moreno, S.N.J.; Hortua Triana, M.A. Characterization of Two EF-hand Domain-containing Proteins from Toxoplasma gondii. J. Eukaryot. Microbiol. 2019, 66, 343–353. [Google Scholar] [CrossRef]
- Durussel, I.; Blouquit, Y.; Middendorp, S.; Craescu, C.T.; Cox, J.A. Cation- and peptide-binding properties of human centrin 2. FEBS Lett. 2000, 472, 208–212. [Google Scholar] [CrossRef]
- Ogunrinde, A.; Munro, K.; Davidson, A.; Ubaid, M.; Snedden, W.A. Arabidopsis Calmodulin-Like Proteins, CML15 and CML16 Possess Biochemical Properties Distinct from Calmodulin and Show Non-overlapping Tissue Expression Patterns. Front. Plant Sci. 2017, 8, 2175. [Google Scholar] [CrossRef] [PubMed]
- La Verde, V.; Dominici, P.; Astegno, A. Towards Understanding Plant Calcium Signaling through Calmodulin-Like Proteins: A Biochemical and Structural Perspective. Int. J. Mol. Sci 2018, 1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MORENO, S.N.J.; ZHONG, L. Acidocalcisomes in Toxoplasma gondii tachyzoites. Biochem. J. 1996, 313, 655–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radu, L.; Durussel, I.; Assairi, L.; Blouquit, Y.; Miron, S.; Cox, J.A.; Craescu, C.T. Scherffelia dubia centrin exhibits a specific mechanism for Ca2+-controlled target binding. Biochemistry 2010, 49, 4383–4394. [Google Scholar] [CrossRef]
- Sheehan, J.H.; Bunick, C.G.; Hu, H.; Fagan, P.A.; Meyn, S.M.; Chazin, W.J. Structure of the N-terminal calcium sensor domain of centrin reveals the biochemical basis for domain-specific function. J. Biol. Chem. 2006, 281, 2876–2881. [Google Scholar] [CrossRef] [Green Version]
- Cates, M.S.; Teodoro, M.L.; Phillips, G.N., Jr. Molecular mechanisms of calcium and magnesium binding to parvalbumin. Biophys. J. 2002, 82, 1133–1146. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Sites Used for Modelling | n | Ka (M−1) | ΔH (kcal mol−1) | |
|---|---|---|---|---|
| TgCEN1-C | 2 | n1 = 0.7 ± 0.1 | 4.8 × 105 ± 6.1 × 103 | −9.6 ± 1.1 |
| n2 = 1.2 ± 0.3 | 3.9 × 104 ± 4.5 × 103 | −3.2 ± 0.7 | ||
| TgCEN2-C | 1 | 0.8 ± 0.1 | 1.6 × 104 ± 1.5 × 103 | −3.4 ± 0.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bombardi, L.; Pedretti, M.; Conter, C.; Dominici, P.; Astegno, A. Distinct Calcium Binding and Structural Properties of Two Centrin Isoforms from Toxoplasma gondii. Biomolecules 2020, 10, 1142. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10081142
Bombardi L, Pedretti M, Conter C, Dominici P, Astegno A. Distinct Calcium Binding and Structural Properties of Two Centrin Isoforms from Toxoplasma gondii. Biomolecules. 2020; 10(8):1142. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10081142
Chicago/Turabian StyleBombardi, Luca, Marco Pedretti, Carolina Conter, Paola Dominici, and Alessandra Astegno. 2020. "Distinct Calcium Binding and Structural Properties of Two Centrin Isoforms from Toxoplasma gondii" Biomolecules 10, no. 8: 1142. https://0-doi-org.brum.beds.ac.uk/10.3390/biom10081142