
Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications



1
Department of Obstetrics and Gynaecology, Shenzhen Baoan Women’s and Children’s Hospital, Shenzhen University, Shenzhen 518133, China
2
Department of Obstetrics and Gynaecology, Faculty of Medicine, The Chinese University of Hong Kong, Hong Kong, China
3
Department of Reproductive Health, Shenzhen Baoan Women’s and Children’s Hospital, Shenzhen University, Shenzhen 518133, China
4
Department of Central Lab, Shenzhen Baoan Women’s and Children’s Hospital, Shenzhen University, Shenzhen 518133, China
*
Authors to whom correspondence should be addressed.
†
X.G. and H.Y. contributed equally to this work.
Biomolecules 2021, 11(2), 253; https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020253
Submission received: 30 December 2020
/
Revised: 6 February 2021
/
Accepted: 8 February 2021
/
Published: 10 February 2021
(This article belongs to the Special Issue Vascular Endothelial Growth Factor (VEGF): From Basic Mechanisms to Clinical Significance and Drug Development)
Abstract
:Vascular endothelial growth factor (VEGF) is a well-known angiogenic factor that plays a critical role in various physiological and pathological processes. VEGF also contributes to the process of embryo implantation by enhancing embryo development, improving endometrial receptivity, and facilitating the interactions between the developing embryo and the endometrium. There is a correlation between the alteration of VEGF expression and reproductive failure, including recurrent implantation failure (RIF) and recurrent miscarriage (RM). In order to clarify the role of VEGF in embryo implantation, we reviewed recent literature concerning the expression and function of VEGF in the reproductive system around the time of embryo implantation and we provide a summary of the findings reported so far. We also explored the effects and the possible underlying mechanisms of action of VEGF in embryo implantation.
1. Introduction
Vascular endothelial growth factor (VEGF) is a multi-functional factor primarily involved in the regulation of proliferation, differentiation and survival of endothelial cells as well as in vascular permeability [1]. The family of VEGF consists of a group of growth proteins including VEGF-A–VEGF-F, placental growth factor (PlGF), and endocrine gland-derived vascular endothelial growth factor (EG-VEGF) [2]. VEGF-A (also called VEGF), which was firstly described by Senger et al. in 1983 [3], has been proved to be the most important and potent factor in angiogenesis [2]. PlGF, on the other hand, is thought to be selectively involved in pathological angiogenesis, for instance, in tumors and in ischemic and inflammatory processes [4,5]. VEGF-B is more involved in the growth, differentiation, and survival of certain types of cells [6,7], while VEGF-C and VEGF-D are primarily implicated in lymphangiogenesis [8,9].
VEGFs exert their effects mainly through binding to tyrosine kinase receptors: fms-like tyrosine kinase 1 (Flt-1, also termed VEGFR-1), kinase insert domain receptor (KDR, also termed VEGFR-2), and Flt-4 (also termed VEGFR-3) [2]. VEGFR-2, which has the strongest pro-angiogenic activity, is mainly expressed in vascular endothelial cells and can bind to VEGF [2]. Compared with VEGFR-1, VEGFR-2 has a higher tyrosine kinase activity but a lower affinity for VEGF [10,11]. Besides the expression in endothelial cells, VEGFR-1 is also expressed in macrophage-lineage cells [11]. VEGFR-1 can interact with VEGF, VEGF-B, and PlGF [12,13]. With a higher affinity but lower kinase activity, VEGFR-1 acts more like a decoy, a negative regulator of VEGF [14,15]. sVEGFR-1 (sFlt-1), a soluble form of VEGFR-1, can also trap VEGF, VEGF-B, and PlGF and therefore block their binding to membrane receptors [16,17], and its activity has been proven to be strongly correlated with unexplained infertility [18,19], recurrent miscarriage [20], and adverse pregnancy outcomes [21]. PlGF, which shows lower affinity for VEGFR-1 than VEGF [22], can replace VEGF in the VEGFR-1 “sink” and thus potentiate the angiogenic effect of VEGF, since VEGFR-2 is the main receptor with a pro-angiogenesis effect [4,12]. VEGFR-3 is expressed in lymphatic endothelia and high endothelial venules [23]. Through binding with VEGF-C or VEGF-D, VEGFR-3 transduces signals for lymphangiogenesis [2]. In addition to these three tyrosine kinase receptors, VEGF can also bind to neuropilins which act as co-receptors [24].
VEGF is implicated in a wide variety of physiological and pathological conditions. During the process of embryo development, VEGF participates in embryonic vasculogenesis and angiogenesis [25,26]. Moreover, in postnatal development, there is accumulating evidence showing the crucial role of VEGF in body growth and organ development [27,28,29]. In inflammation, VEGFR-1 plays a role in the recruitment and activation of monocytes and macrophages [30,31,32]. In oncogenesis, VEGF is responsible for tumor growth and metastasis, based on which anti-angiogenic therapy has achieved great progress in tumor treatment [1]. In the field of reproduction, extensive efforts have been made to clarify the role of VEGF in embryo implantation. However, there is a lack of a comprehensive summary of the existing data. In this review article, we summarized recent literature concerning the expression and function of VEGF in the reproductive system around the time of embryo implantation. We also explored the effects and the possible underlying mechanisms of action of VEGF in embryo implantation.
2. The Role of VEGF in the Reproductive System during Embryo Implantation
2.1. VEGF in Human Endometrium
Endometrium lines the inside of the uterus and undergoes cyclic breakdown and remodeling, which are accompanied with the reconstruction of the vascular system. During the menstrual cycle, there is only a short and critical period of time allowing an embryo to implant, when the endometrium becomes a well-vascularized tissue characterized by increased vascular permeability, oedema, and angiogenesis [33]. Although a myriad of factors and cytokines are thought to be involved in this transformation, VEGF, as a potent angiogenic factor, plays a central role.
Extensive efforts have been made to explore the spatial and temporal expression change of VEGF in the endometrium across the menstrual cycle. Several studies have found the expression of VEGF in the endometrium throughout the menstrual cycle, with a significant increase in the mid-luteal phase, suggesting a role of endometrial VEGF around the time of embryo implantation [34,35,36]. In addition, adequate expression of VEGF is essential for successful pregnancy, based on previous finding that endometrial VEGF expression is impaired in the peri-implantation period in infertile patients [37,38]. Functional experiments have shown that the addition of VEGF could promote embryo outgrowth as well as the adhesive capacity of endometrial epithelial cells [39]. In women with recurrent miscarriage, VEGF showed a lower expression level in the endometrium during the mid-luteal phase [40]. However, one of our previous studies has identified elevated VEGF levels in women with hampered pregnancy potential [41]. In that study, endometrial tissues were obtained from women with elevated progesterone level. Progesterone and estrogen are two main regulators of endometrium changes across the menstrual cycle and have significant promoting effects on VEGF production [42,43]. Therefore, the analysis of VEGF levels should take into account the effects of estrogen, progesterone, and other potential influential factors to understand the role of VEGF during embryo implantation.
In accordance with the upregulation of VEGF, VEGFR-1 and VEGFR-2 are also upregulated in the glandular epithelium in the secretory phase [44]. sFlt-1, a soluble antagonist of VEGF and PlGF, is increased distinctively in the proliferative phase but decreased to almost 25% of its proliferative phase peak value in the endometrium during the luteal phase [45]. The analysis of serum sFlt-1 level showed that infertile women had higher serum sFlt-1 levels in the secretory phase than in the proliferative phase [19]. Experimentally, sFlt-1 expression in human primary endometrial stromal cells is also found to be turned off during decidualization, whereas VEGF expression showed an opposite change [46]. The upregulation of VEGF and the coordinated downregulation of sFlt-1 in the luteal phase may serve to increase endometrial angiogenesis and vascular permeability, which are crucial for endometrium preparation for embryo implantation.
Histochemistry staining reported that PlGF, which exists in glandular and luminal epithelial cells, decidual stromal cells, and uterine fluid, showed increased staining intensity in the endometrium around the time of embryo implantation [47]. Santi et al. also demonstrated that PlGF expression was higher in the endometrium of women with successful embryo implantation compared to those who failed to conceive [48]. In that study, the authors firstly reported that PlGF gene expression was positively correlated with the hysteroscopic appearance of the endometrium based on the Sakumoto-Masamoto grading system. In contrast, one of our previous studies found that PlGF was upregulated in women with a high serum progesterone level, and this increase might be a cause of impaired receptivity of the endometrium in those women [41]. As described before, PlGF and VEGF are two synergistic molecules acting on angiogenesis. The upregulation of these two cytokines in the endometrium during the time of embryo implantation suggests that angiogenesis is crucial for successful embryo implantation. Moreover, the upregulation of VEGF and its receptors in the secretory phase might aim to keep pace with the rapidly thickening endometrium. Another study also showed that stromal VEGF intensity and stromal microvessel density (MVD) could be positive predictors of pregnancy outcomes in preparation of frozen embryo transfer (FET) cycles [49].
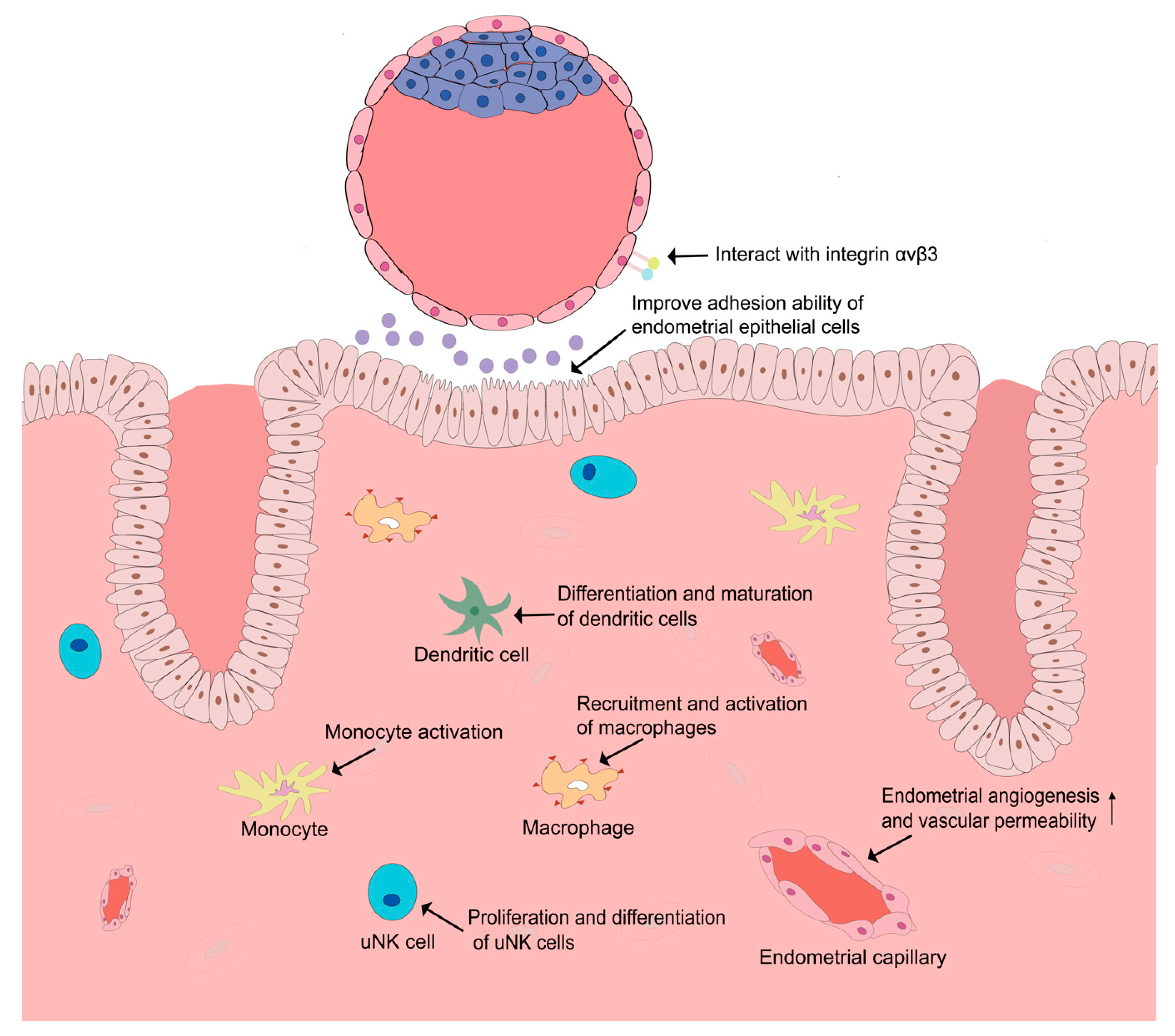
It is worth noting that besides an angiogenic effect, VEGF and PlGF are also functionally related to immune regulation. An embryo is an allograft to the maternal immune system, and immunologic tolerance is critical for the successful establishment of embryo implantation. During the establishment of immuno-tolerance, numerous immunosuppressive mechanisms and various immunocytes, including uterine natural killer (uNK) cells, macrophages, and dendritic cells, are thought to play significant roles [50,51]. Several studies have demonstrated that VEGF and PlGF might serve as immune modulators and mediate the immuno-tolerance of the maternal immune system during the time of embryo implantation. Monocytes, well-characterized immunocytes [52], could be activated by VEGF and PlGF [53,54]. VEGF and PlGF also promote the recruitment and activation of macrophages [32,55,56]. PlGF is also found expressed in uNK cells and plays a significant role in the proliferation and differentiation of uNK cells [57]. PlGF is also implicated in the regulation of the differentiation and maturation of dendritic cells and is able to skew type 1 T helper immune response to the Th2 phenotype [58]. All these immunoregulatory functions of VEGF and PlGF might be favorable for the establishment of pregnancy (Figure 1, Table 1).
2.2. VEGF in the Development of Human Oocytes and Embryo
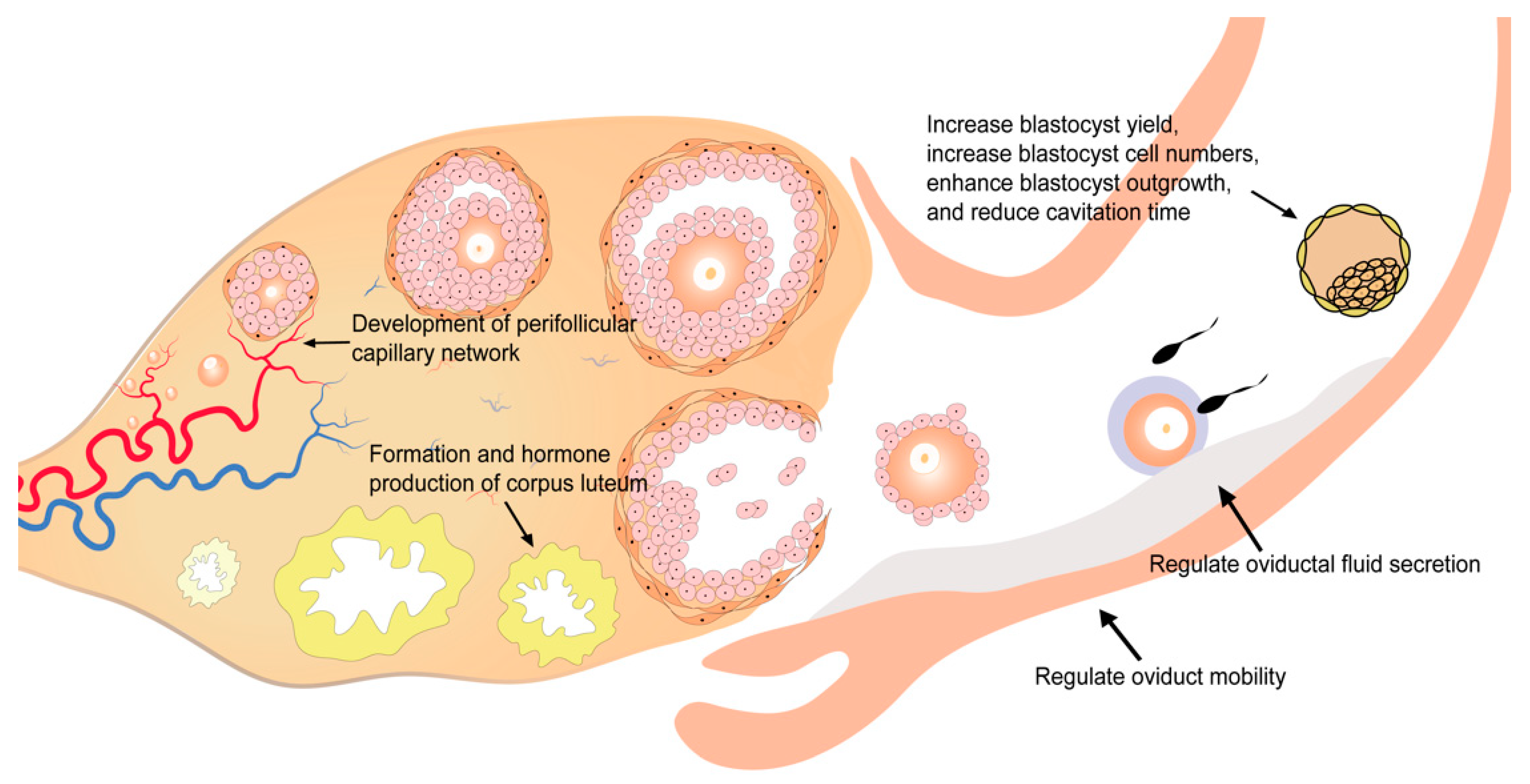
The growth and development of follicles are dependent on an adequate blood supply to obtain nutrients and oxygen. Insufficient blood supply could lead to a low-oxygen status in follicular fluid and severely disrupt the development of oocytes and, subsequently, of the embryo [71]. The inhibition of angiogenesis could lead to disruption of follicular development, ovulation, and endocrine functions of the ovary [72,73]. VEGF is secreted by granulosa cells and theca cells and can be detected in the follicular fluid [74,75]. A high concentration of VEGF in the follicular fluid is correlated with increased perifollicular vascularity, higher fertilization rates, better embryo quality, and higher pregnancy rates [61]. As VEGF in the follicular fluid is mainly secreted from granulosa and theca cells, it is likely that a higher VEGF level indicates better functioning of these cells and thus the presence of a better microenvironment for follicle development. In contrast, there are several studies indicating that VEGF concentration in follicular fluid is negatively correlated with in vitro fertilization (IVF) pregnancy outcomes [76,77]. However, in these studies, women with polycystic ovary syndrome (PCOS) were included. It is well recognized that PCOS patients have a higher level of VEGF and compromised IVF outcomes [78]. The inclusion of PCOS patients could be a remarkable influential factor in these analyses.
VEGF also acts in the formation and function of corpus luteum (Table 1, Figure 2). Corpus luteum is a transient endocrine gland that develops from residual follicular tissues after ovulation in the presence of active angiogenesis [79]. The ruptured follicle is under hypoxia conditions after ovulation, when hypoxia-inducible factor-1 (HIF1) is induced and promotes the angiogenesis of corpus luteum via VEGF [62]. Besides the formation of corpus luteum, VEGF also contributes to progesterone production by corpus luteum [63]. The reduction of VEGF and VEGFR-2 levels in corpus luteum is related to the impairment of the luteal circulation [80]. Blocking of VEGF signaling via VEGFR inhibition is proved to induce decreased luteal endothelial networks, vascular endothelial cell detachment, apoptosis of luteal steroid-producing epithelial cells, disrupted luteal function, embryonic development arrest, and preterm birth [63,80,81].
With a highest level during the periovulatory period, VEGF shows dynamic changes in the human oviduct across the menstrual cycle and is thought to be implicated in the secretion of oviductal fluid via regulating the vascular permeability of the oviduct (Table 1, Figure 2) [65,66]. Furthermore, VEGF also plays a role in oviduct mobility (Table 1, Figure 2). VEGF regulates oviduct contraction by stimulating the biosynthesis and release of prostaglandin E2 (PGE2), prostaglandin F2α (PGF2α), and endothelin-1 (ET-1) in bovine oviductal epithelial cells, which is important for gametes transport, fertilization, and embryo transport [64].
VEGF can be detected throughout the process of embryo development [82,83]. The analysis of embryos cultured in vitro also demonstrated that the expression of VEGF and two VEGF receptors (VEGFR-1 and VEGFR-2) was maintained during embryo development [84]. Functional in vitro experiments in embryos suggest that the addition of VEGF could increase blastocyst yield and blastocyst cell numbers, enhance blastocyst outgrowth, and reduce cavitation time [39,67,68]. As another member of the VEGF family, PlGF shows a similar effect on embryo development: increased blastocyst cell numbers and enhanced blastocyst outgrowth [47]. However, the molecular mechanisms underlying these effects remain unclear.
2.3. VEGF in the Interaction between Endometrium and Embryos
After an embryo enters the uterus lumen, the interaction between embryo and endometrium starts in the uterine fluid through locally produced soluble mediators. Uterine fluid, also called uterine secretion, is mainly produced by uterine glands. During the peri-implantation period, endometrial glands secrete important mediators that facilitate pre-implantation embryo development and embryo implantation. Several VEGF family members such as VEGF and PlGF have been identified in uterine fluid [39,47]. It is demonstrated that VEGF concentration was upregulated in mid-luteal uterine fluid [39]. The significant role of VEGF in the dialogue between embryo and endometrium is further evidenced by functional experiments where enhanced blastocyst outgrowth, improved endometrial epithelial cell adhesion ability, reduced cavitation time, increased blastocyst cell numbers, increased implantation rates, and enhanced fetal limb development were observed after VEGF treatment [39,67]. PlGF is found upregulated in endometrial glands, with strong corresponding staining on the apical surface of the epithelium in mid-secretory phase [47]. Functional studies of PlGF showed similar results, indicating that PlGF increased blastocyst cell numbers, enhanced blastocyst outgrowth, and improved endometrial epithelial cells’ adhesion ability [47]. In addition to the effect of endometrium-derived VEGFs on the embryo, embryo-derived factors and cytokines are also involved in the regulation of the uterine microenvironment. For example, embryo-derived VEGF-A stimulates endometrial angiogenesis, which enables embryos to induce angiogenesis directly at the implantation site [69]. Human chorionic gonadotropin (hCG), produced by the embryo, is also a stimulator of VEGF action in the endometrium [85]. Taken these results together, although less is known about the effect of the embryo on the endometrium, the crosstalk of embryo and endometrium might be extensive and influence embryo implantation in a more complex manner (Figure 1, Table 1).
Furthermore, VEGFR-2 could interact with integrin αvβ3 [70], which is an important adhesion molecule during embryo implantation [86], in VEGF-induced angiogenesis. Integrin αvβ3 is found to be expressed in endometrial pinopodes and blastocyst trophectoderm [87], and the ligand of integrin αvβ3, the glycoprotein osteopontin (OPN), is also expressed in the endometrium. The interaction of embryonic integrin αvβ3 and endometrial OPN is thought to be involved in embryo adhesion to the luminal epithelium of the endometrium, while the binding of VEGFR-2 and integrin could shift the cell surface localization of VEGFR-2 to focal adhesions and induce endothelial cell polarization, which could also be a mechanism facilitating embryo implantation [88]. However, further studies are still necessary to explore how this interaction between VEGFR-2 and integrin αvβ3 influences embryo implantation.
3. VEGF in Reproductive Failure
3.1. Recurrent Implantation Failure (RIF)
RIF occurs when a woman under the age of 40 fails to achieve a clinical pregnancy after transfer of at least four good-quality embryos in a minimum of three fresh or frozen embryo transfer cycles [89]. There are numerous factors that could lead to RIF, such as uterine abnormalities, advanced maternal age, elevated body mass index, immunological factors, and abnormal angiogenesis [90,91].
One earlier study has reported an increased serum VEGF level in women with RIF compared to fertile controls [92], whereas one of our earlier studies demonstrated decreased VEGF expression in all regions of the endometrium at the time of embryo implantation [91]. The discrepancy between these two studies might derive from different sampling time and different sample types. Further studies are needed to explore the expression pattern of VEGF in RIF patients. Furthermore, some studies have explored the association between VEGF polymorphisms and the occurrence of embryo implantation failure. These results suggest polymorphisms of the VEGF gene could impact fertilization rate, embryo implantation rate, and pregnancy rate [93]. In addition, VEGF polymorphisms such as VEGF-1154A/A, which are related to altered VEGF expression, increase the risk of RIF [94,95,96,97]. However, whether VEGF influences embryo implantation in RIF patients remains largely unknown.
3.2. Recurrent Miscarriage (RM)
RM is defined as three or more consecutive miscarriages before 24 weeks of gestation [98]. Although several known causes of RM, including parental chromosomal anomalies, uterine malformation, endocrinological disorders, and immunological abnormality, are known, around half of RM cases remain unexplained [99]. Recent evidence has indicated that altered expression of VEGF family members might be a contributory factor to RM [20,100].
Lash et al. explored the expression of VEGF and its receptors in women with unexplained recurrent miscarriage [38]. The results demonstrated that, during mid-late secretory phase, the expression of VEGF-A in vascular smooth muscle cells (VSMCs), endothelial cells (ECs), and glandular epithelial cells was decreased, while the expression of VEGFR-1 in stromal cells, VSMCs, ECs, and glandular epithelial cells was increased in RM patients. This study also showed a decreased expression of VEGFR-2 in VSMCs and stromal cells, an increased expression of VEGFR-3 in glandular epithelial cells, and a greater proportion of mature vessels in the endometrium around the time of embryo implantation in women with RM. Banerjee et al. identified lower VEGF expression in endometrial tissues from patients with idiopathic recurrent miscarriage (IRM) in the peri-implantation period [101], and the observed downregulation of VEGF might be a product of the downregulation of angiogenic cytokines including interleukin (IL)-2, IL-6, and IL-8. In another previous study, Amirchaghmaghi et al. analyzed the gene expression of VEGF, VEGFR-1, and VEGFR-2 and found that unexplained RM (URM) patients had a lower VEGF level and higher VEGFR-1 and VEGFR-2 levels in the endometrium between day 19th and 24th of the menstrual cycle when compared to fertile controls [100]. In contrast, in one of our earlier studies, RM patients were found to have a higher VEGF level in luminal epithelium, glandular epithelium, and stroma of the endometrium around the time of embryo implantation [91]. RM patients were also found to have higher HIF1α expression and increased number of micro-blood vessels in the endometrium [102]. These contradictory results imply the multifactorial nature of the etiology of RM, although all patients were diagnosed with unexplained RM at the time of sampling.
In addition to the endometrial tissues, there are also some studies focusing on VEGF expression in other types of tissue from women with recurrent miscarriage. By sampling chorionic villi from RM patients, Pang et al. found that VEGF and sFlt-1 levels were increased when compared to their levels in women with a normal pregnancy [103]. However, other studies demonstrated that VEGF expression in the chorionic villi and decidua from women with recurrent pregnancy loss (RPL) was significantly decreased compared with the expression in women with normal pregnancy [104,105], while in peripheral blood, serum VEGF expression was higher in URM patients between day 19th and 24th of the menstrual cycle when compared to VEGF level in fertile controls [100]. In accordance with these results, Pang et al. also found that the serum levels of VEGF and sFlt-1 in RM women were significantly higher than in women with normal pregnancy [20]. However, several studies reported opposite results, though the time of serum sampling was not mentioned [106,107,108]. Therefore, given the significantly different changes during the menstrual cycle and pregnancy, it is crucial to standardize the timing of sampling in future studies.
Although there is compelling evidence showing dysregulation of VEGF in a number of common diseases, the underlying causes of this dysregulation remain unknown. Genetic variation might be one of the potential causes responsible for dysregulated VEGF expression [109]. A meta-analysis of 10 independent case–control studies revealed that rs1570360, rs3025039, rs2010963, and rs3025020 polymorphisms of VEGF were associated with elevated RM risk [110]. Furthermore, Su et al. reported that KDR polymorphisms were correlated with RPL [111]. Hence, further studies are needed to illustrate the mechanisms by which VEGF is regulated in RM.
3.3. Endometriosis
Endometriosis is a common cause of infertility in women of reproductive age. There is accumulating evidence showing that eutopic endometrium in women with endometriosis displays higher VEGF expression [112,113]. At the same time, estrogen receptor is also upregulated in eutopic endometrium of patients with endometriosis [112]. Thus, the endometrium of endometriosis patients might be hyper-responsive to estrogen stimulation and thus enhance the expression of VEGF. In addition, the dysregulation of VEGFRs (downregulation of VEGFR-1 and upregulation of VEGFR-2) seems to be responsible for endometriosis [114]. Kim et al. have shown that expression of VEGFR-1, VEGFR-2, and VEGFR-3 was higher in the mid-luteal endometrium of patients with endometriosis [115]. The authors also conducted a subgroup analysis and found that in patients with endometriosis, non-pregnant subjects had higher VEGFR-1 and VEGFR-3 expression than pregnant subjects [115]. Moreover, endometriotic mesenchymal stem cells of ectopic lesions from endometriosis patients exhibited unique biological characteristics, with increased production of angiogenic factors including VEGF and platelet-derived growth factor (PDGF), which implies intrinsic defects in these cells [116]. In addition, anti-VEGF therapy could attenuate the progress of endometriosis and may be a novel strategy for endometriosis treatment [117]. All these findings suggest the important role of VEGF in endometriosis.
3.4. PCOS
PCOS is a common endocrine disease characterized by oligomenorrhea or amenorrhea, hyperandrogenism, and the presence of polycystic ovary. In addition to endocrine disorders, ovarian hyperplasia and hypervascularity are also two common features of PCOS and are thought to be related to extensive angiogenesis of the ovary [118,119]. These angiogenic disorders might be associated with the decreased ovulation rates of PCOS patients [120]. Immunohistochemistry analysis found increased VEGF expression in PCOS ovary [121], which might be correlated with increased vascularity in the ovarian stroma and the higher incidence of ovarian hyperstimulation syndrome (OHSS) observed in PCOS patients [122,123,124]. After VEGF inhibitor treatment, the ovary from PCOS rats demonstrated a decreased percentage of primary follicles and improved ovulation and follicular development [125].
It has been proved that VEGF levels in serum and follicular fluid of PCOS patients were significantly higher, while sFlt-1 level was lower than in normal ovulatory women [78,126]. Taken these results together, the upregulation of VEGF and the downregulation of VEGF antagonists may jointly increase VEGF bioavailability in PCOS patients. However, in PCOS patients, the expression of VEGF in the endometrium around the time of embryo implantation is lower than in controls [127]. As VEGF is upregulated in the mid-luteal phase in normal endometrium as described above, the downregulation of VEGF might be a contributory factor to impaired endometrial receptivity in patients with PCOS. Moreover, VEGF polymorphisms are also found to be related to PCOS [128,129].
3.5. Preeclampsia (PE)
PE is a serious pregnancy complication characterized with hypertension and proteinuria. A defect in placental angiogenesis is considered to be one of the factors responsible for the pathogenesis of PE [130]. Significant attention has been given to the alteration of the VEGF system in PE, since it is the main regulator of placental angiogenesis.
sFlt-1 was found to be upregulated [131,132,133,134], whilst VEGF and PlGF were downregulated in serum and placenta of PE patients [131,132,135,136]. VEGFR-1 and VEGFR-2 were found to be overexpressed in the placenta from patients suffering from PE [130]. Placentation is a process with extensive angiogenesis in order to establish an appropriate vascular network between mother and fetus [137,138,139]. VEGF mediates angiogenesis and has an anti-apoptotic effect on vascular endothelium cells [2]. VEGF also plays a role in the proliferation, migration, and endovascular differentiation of trophoblast cells [140,141]. Thus, the alteration of the VEGF system could lead to placental malfunction. Maynard et al. have elucidated the central role of sFlt-1 in the pathogenesis of PE [133]. At an early stage of pregnancy, hypoxia increases sFlt-1 production by placental cytotrophoblasts [142]. Subsequently, excess sFlt-1 produced by the placenta leads to endothelial dysfunction, hypertension, and proteinuria by trapping VEGF and PlGF [133]. Thus, among these changes in the VEGF system, the elevation of sFlt-1 might play a central role, since decreased concentrations of VEGF and PlGF might be a consequence of an elevated level of circulating sFlt-1.
Based on the fact that the process of angiogenesis is regulated by numerous pro-angiogenic and anti-angiogenic factors and cytokines as well as angiogenic receptors, there is an increasing number of studies focusing on the balance between pro-angiogenic and anti-angiogenic factors in preeclampsia [143,144]. In some studies, the ratios sFlt-1/PlGF and VEGF/ sFlt-1 were determined to study the association of angiogenesis with certain pathological conditions [19,21,145].
3.6. Anti-Angiogenic Therapy
Over these years, the development of anti-VEGF agents has achieved great progress [146,147]. A number of anti-angiogenic agents including bevacizumab have been applied clinically in tumor treatment [1]. Associated with conventional chemotherapy, bevacizumab significantly improved the prognosis of patients with metastatic colorectal cancer or non-small-cell lung cancer [148,149,150,151,152]. Anti-angiogenic therapy is also proved to be effective in the treatment of non-tumor diseases, for instance, age-related macular degeneration [153]. However, as stated above, due to the complex role of VEGF in the reproductive system, anti-VEGF therapy has not been applied in clinical practice in the field of reproductive medicine yet. Although some studies have reported the possible therapeutic role of VEGF inhibitors in women with PCOS [125], the clinical application of anti-VEGF therapy should be cautious before we fully understand the underlying mechanism whereby VEGF influences embryo implantation and the pharmacokinetic characteristics of these drugs.
4. Conclusions
VEGF is an important angiogenic factor in many physiological and pathological conditions. In this review, we summarized recent data on the role of VEGF in embryo implantation and reproductive failure. The existing data show that VEGF plays multifaceted roles in embryo implantation and that the alteration of the expression of VEGF, including VEGF polymorphisms, could lead to infertility and pregnancy complications. However, little is known about the mechanisms whereby VEGF influences embryo implantation or the root causes of the observed alterations in VEGF expression. Further studies are in urgent need to clarify the role of VEGF in successful pregnancy and reproductive failure.
Author Contributions
Conceptualization, X.G. and X.C.; methodology, X.G.; data curation, X.G. and H.Y.; writing—original draft preparation, X.G., H.Y. and Y.W.; writing—review and editing, H.W. and X.C.; supervision, T.C.L., H.W. and X.C.; funding acquisition, H.W. and X.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Hong Kong Health and Medical Research Fund (06170186), National Natural Science Foundation of China (81741004) and Shenzhen Key Medical Discipline Construction Fund (SZXK028).
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Senger, D.R.; Galli, S.J.; Dvorak, A.M.; Perruzzi, C.A.; Harvey, V.S.; Dvorak, H.F. Tumor cells secrete a vascular permeability factor that promotes accumulation of ascites fluid. Science 1983, 219, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Moons, L.; Luttun, A.; Vincenti, V.; Compernolle, V.; De Mol, M.; Wu, Y.; Bono, F.; Devy, L.; Beck, H.; et al. Synergism between vascular endothelial growth factor and placental growth factor contributes to angiogenesis and plasma extravasation in pathological conditions. Nat. Med. 2001, 7, 575–583. [Google Scholar] [CrossRef]
- Van Bergen, T.; Etienne, I.; Cunningham, F.; Moons, L.; Schlingemann, R.O.; Feyen, J.H.M.; Stitt, A.W. The role of placental growth factor (PlGF) and its receptor system in retinal vascular diseases. Prog. Retin. Eye Res. 2019, 69, 116–136. [Google Scholar] [CrossRef] [PubMed]
- Lal, N.; Chiu, A.P.; Wang, F.; Zhang, D.; Jia, J.; Wan, A.; Vlodavsky, I.; Hussein, B.; Rodrigues, B. Loss of VEGFB and its signaling in the diabetic heart is associated with increased cell death signaling. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1163–H1175. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Li, D.; Wang, X.; Jia, J.; Chen, Y.; Yao, Y.; Zhao, C.; Lu, X.; Zhang, S.; Togo, J.; et al. VEGF and VEGFB Play Balancing Roles in Adipose Differentiation, Gene Expression, and Function. Endocrinology 2018, 159, 2036–2049. [Google Scholar] [CrossRef] [Green Version]
- Bui, H.M.; Enis, D.; Robciuc, M.R.; Nurmi, H.J.; Cohen, J.; Chen, M.; Yang, Y.; Dhillon, V.; Johnson, K.; Zhang, H.; et al. Proteolytic activation defines distinct lymphangiogenic mechanisms for VEGFC and VEGFD. J. Clin. Investig. 2016, 126, 2167–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astin, J.W.; Haggerty, M.J.; Okuda, K.S.; Le Guen, L.; Misa, J.P.; Tromp, A.; Hogan, B.M.; Crosier, K.E.; Crosier, P.S. Vegfd can compensate for loss of Vegfc in zebrafish facial lymphatic sprouting. Development 2014, 141, 2680–2690. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef]
- Shibuya, M. Differential roles of vascular endothelial growth factor receptor-1 and receptor-2 in angiogenesis. J. Biochem. Mol. Biol. 2006, 39, 469–478. [Google Scholar] [CrossRef]
- Park, J.E.; Chen, H.H.; Winer, J.; Houck, K.A.; Ferrara, N. Placenta growth factor. Potentiation of vascular endothelial growth factor bioactivity, in vitro and in vivo, and high affinity binding to Flt-1 but not to Flk-1/KDR. J. Biol. Chem. 1994, 269, 25646–25654. [Google Scholar] [CrossRef]
- Olofsson, B.; Korpelainen, E.; Pepper, M.S.; Mandriota, S.J.; Aase, K.; Kumar, V.; Gunji, Y.; Jeltsch, M.M.; Shibuya, M.; Alitalo, K.; et al. Vascular endothelial growth factor B (VEGF-B) binds to VEGF receptor-1 and regulates plasminogen activator activity in endothelial cells. Proc. Natl. Acad. Sci. USA 1998, 95, 11709–11714. [Google Scholar] [CrossRef] [Green Version]
- Hiratsuka, S.; Minowa, O.; Kuno, J.; Noda, T.; Shibuya, M. Flt-1 lacking the tyrosine kinase domain is sufficient for normal development and angiogenesis in mice. Proc. Natl. Acad. Sci. USA 1998, 95, 9349–9354. [Google Scholar] [CrossRef] [Green Version]
- Fong, G.H.; Rossant, J.; Gertsenstein, M.; Breitman, M.L. Role of the Flt-1 receptor tyrosine kinase in regulating the assembly of vascular endothelium. Nature 1995, 376, 66–70. [Google Scholar] [CrossRef]
- Kendall, R.L.; Wang, G.; Thomas, K.A. Identification of a natural soluble form of the vascular endothelial growth factor receptor, FLT-1, and its heterodimerization with KDR. Biochem. Biophys. Res. Commun. 1996, 226, 324–328. [Google Scholar] [CrossRef]
- Shibuya, M. VEGF-VEGFR System as a Target for Suppressing Inflammation and other Diseases. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Edgell, T.A.; Evans, J.; Lazzaro, L.; Boyes, K.; Sridhar, M.; Catt, S.; Rombauts, L.J.F.; Vollenhoven, B.J.; Salamonsen, L.A. Assessment of potential biomarkers of pre-receptive and receptive endometrium in uterine fluid and a functional evaluation of the potential role of CSF3 in fertility. Cytokine 2018, 111, 222–229. [Google Scholar] [CrossRef]
- Wathén, K.A.; Unkila-Kallio, L.; Isaksson, R.; Tiitinen, A.; Stenman, U.H.; Vuorela, P. Is serum-soluble vascular endothelial growth factor receptor-1 of importance in unexplained infertility? Acta Obs. Gynecol. Scand. 2008, 87, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Wei, Z.; Li, O.; Huang, R.; Qin, J.; Chen, H.; Fan, X.; Chen, Z.J. An increase in vascular endothelial growth factor (VEGF) and VEGF soluble receptor-1 (sFlt-1) are associated with early recurrent spontaneous abortion. PLoS ONE 2013, 8, e75759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, I.; Chan, Y.; Sriprasert, I.; Louie, K.; Ingles, S.; Stanczyk, F.; McGinnis, L.K.; Chung, K. The role of angiogenic markers in adverse perinatal outcomes: Fresh versus frozen embryo transfers. J. Assist. Reprod. Genet. 2017, 34, 1639–1643. [Google Scholar] [CrossRef]
- Persico, M.G.; Vincenti, V.; DiPalma, T. Structure, expression and receptor-binding properties of placenta growth factor (PlGF). Curr. Top. Microbiol. Immunol. 1999, 237, 31–40. [Google Scholar] [PubMed]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, F.; Goshima, Y. Structural and functional relation of neuropilins. Adv. Exp. Med. Biol. 2002, 515, 55–69. [Google Scholar]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Theis, V.; Theiss, C. VEGF—A Stimulus for Neuronal Development and Regeneration in the CNS and PNS. Curr. Protein. Pept. Sci. 2018, 19, 589–597. [Google Scholar] [CrossRef]
- Gerber, H.P.; Hillan, K.J.; Ryan, A.M.; Kowalski, J.; Keller, G.A.; Rangell, L.; Wright, B.D.; Radtke, F.; Aguet, M.; Ferrara, N. VEGF is required for growth and survival in neonatal mice. Development 1999, 126, 1149–1159. [Google Scholar]
- Smythe, G. Role of Growth Factors in Modulation of the Microvasculature in Adult Skeletal Muscle. Adv. Exp. Med. Biol. 2016, 900, 161–183. [Google Scholar]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef]
- Zittermann, S.I.; Issekutz, A.C. Endothelial growth factors VEGF and bFGF differentially enhance monocyte and neutrophil recruitment to inflammation. J. Leukoc. Biol. 2006, 80, 247–257. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, Y.H.; Son, J.E.; Lee, J.H.; Kim, S.; Choe, M.S.; Moon, J.H.; Zhong, J.; Fu, K.; Lenglin, F.; et al. Intermittent fasting promotes adipose thermogenesis and metabolic homeostasis via VEGF-mediated alternative activation of macrophage. Cell Res. 2017, 27, 1309–1326. [Google Scholar] [CrossRef]
- Plaisier, M. Decidualisation and angiogenesis. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 259–271. [Google Scholar] [CrossRef]
- Sugino, N.; Kashida, S.; Karube-Harada, A.; Takiguchi, S.; Kato, H. Expression of vascular endothelial growth factor (VEGF) and its receptors in human endometrium throughout the menstrual cycle and in early pregnancy. Reproduction 2002, 123, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Shifren, J.L.; Tseng, J.F.; Zaloudek, C.J.; Ryan, I.P.; Meng, Y.G.; Ferrara, N.; Jaffe, R.B.; Taylor, R.N. Ovarian steroid regulation of vascular endothelial growth factor in the human endometrium: Implications for angiogenesis during the menstrual cycle and in the pathogenesis of endometriosis. J. Clin. Endocrinol. Metab. 1996, 81, 3112–3118. [Google Scholar]
- Torry, D.S.; Holt, V.J.; Keenan, J.A.; Harris, G.; Caudle, M.R.; Torry, R.J. Vascular endothelial growth factor expression in cycling human endometrium. Fertil. Steril. 1996, 66, 72–80. [Google Scholar] [CrossRef]
- Jee, B.C.; Suh, C.S.; Kim, K.C.; Lee, W.D.; Kim, H.; Kim, S.H. Expression of vascular endothelial growth factor-A and its receptor-1 in a luteal endometrium in patients with repeated in vitro fertilization failure. Fertil. Steril. 2009, 91, 528–534. [Google Scholar] [CrossRef]
- Lash, G.E.; Innes, B.A.; Drury, J.A.; Robson, S.C.; Quenby, S.; Bulmer, J.N. Localization of angiogenic growth factors and their receptors in the human endometrium throughout the menstrual cycle and in recurrent miscarriage. Hum. Reprod. 2012, 27, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannan, N.J.; Paiva, P.; Meehan, K.L.; Rombauts, L.J.; Gardner, D.K.; Salamonsen, L.A. Analysis of fertility-related soluble mediators in human uterine fluid identifies VEGF as a key regulator of embryo implantation. Endocrinology 2011, 152, 4948–4956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadekova, O.N.; Nikitina, L.A.; Rashidov, T.N.; Voloschuk, I.N.; Samokhodskaya, L.M.; Demidova, E.M.; Tkachuk, V.A. Luteal phase defect is associated with impaired VEGF mRNA expression in the secretory phase endometrium. Reprod. Biol. 2015, 15, 65–68. [Google Scholar] [CrossRef]
- Chen, X.; Jin, X.; Liu, L.; Man, C.W.; Huang, J.; Wang, C.C.; Zhang, S.; Li, T.C. Differential expression of vascular endothelial growth factor angiogenic factors in different endometrial compartments in women who have an elevated progesterone level before oocyte retrieval, during in vitro fertilization-embryo transfer treatment. Fertil. Steril. 2015, 104, 1030–1036. [Google Scholar] [CrossRef]
- Kim, M.; Park, H.J.; Seol, J.W.; Jang, J.Y.; Cho, Y.S.; Kim, K.R.; Choi, Y.; Lydon, J.P.; Demayo, F.J.; Shibuya, M.; et al. VEGF-A regulated by progesterone governs uterine angiogenesis and vascular remodelling during pregnancy. Embo Mol. Med. 2013, 5, 1415–1430. [Google Scholar] [CrossRef]
- Okada, H.; Tsutsumi, A.; Imai, M.; Nakajima, T.; Yasuda, K.; Kanzaki, H. Estrogen and selective estrogen receptor modulators regulate vascular endothelial growth factor and soluble vascular endothelial growth factor receptor 1 in human endometrial stromal cells. Fertil. Steril. 2010, 93, 2680–2686. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G. Expression of vascular endothelial growth factor and its receptors, flt-1 and kinase insert domain-containing receptor in normal human endometrium during menstrual cycle. Zhonghua Fu Chan Ke Za Zhi 2002, 37, 729–732. [Google Scholar] [PubMed]
- Krüssel, J.S.; Casañ, E.M.; Raga, F.; Hirchenhain, J.; Wen, Y.; Huang, H.Y.; Bielfeld, P.; Polan, M.L. Expression of mRNA for vascular endothelial growth factor transmembraneous receptors Flt1 and KDR, and the soluble receptor sflt in cycling human endometrium. Mol. Hum. Reprod. 1999, 5, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, H.N.; Wu, J.; Rimawi, B.H.; Duran, J.M.; Spencer, J.B.; Sidell, N.; Rajakumar, A. Human endometrial stromal cell plasticity: Reversible sFlt1 expression negatively coincides with decidualization. Hypertens. Pregnancy 2017, 36, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Binder, N.K.; Evans, J.; Salamonsen, L.A.; Gardner, D.K.; Kaitu’u-Lino, T.J.; Hannan, N.J. Placental Growth Factor Is Secreted by the Human Endometrium and Has Potential Important Functions during Embryo Development and Implantation. PLoS ONE 2016, 11, e0163096. [Google Scholar]
- Santi, A.; Felser, R.S.; Mueller, M.D.; Wunder, D.M.; McKinnon, B.; Bersinger, N.A. Increased endometrial placenta growth factor (PLGF) gene expression in women with successful implantation. Fertil. Steril. 2011, 96, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Law, T.S.M.; Cheung, W.C.; Wu, F.; Zhang, R.; Chung, J.P.W.; Wang, C.C.; Chen, X.; Li, T.C. Endometrial Vascularization Characterized by Optical Coherence Tomography and Immunohistochemistry in Women Undergoing In Vitro Fertilization-Embryo Transfer Treatment. Medicina 2019, 55, 81. [Google Scholar] [CrossRef] [Green Version]
- Faas, M.M.; de Vos, P. Uterine NK cells and macrophages in pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Tolerance, suppression and the fetal allograft. J. Mol. Med. 2005, 83, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Immune regulation by monocytes. Semin. Immunol. 2018, 35, 12–18. [Google Scholar] [CrossRef]
- Selvaraj, S.K.; Giri, R.K.; Perelman, N.; Johnson, C.; Malik, P.; Kalra, V.K. Mechanism of monocyte activation and expression of proinflammatory cytochemokines by placenta growth factor. Blood 2003, 102, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Barleon, B.; Sozzani, S.; Zhou, D.; Weich, H.A.; Mantovani, A.; Marmé, D. Migration of human monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood 1996, 87, 3336–3343. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jin, Q.; Yao, Q.; Zhou, Y.; Zou, Y.; Li, Z.; Zhang, S.; Tu, C. Placental Growth Factor Contributes to Liver Inflammation, Angiogenesis, Fibrosis in Mice by Promoting Hepatic Macrophage Recruitment and Activation. Front. Immunol. 2017, 8, 801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, K.C.; Jena, M.K.; Pradhan, B.S.; Nayak, N.; Das, S.; Hsu, C.D.; Wheeler, D.S.; Chen, K.; Nayak, N.R. VEGF may contribute to macrophage recruitment and M2 polarization in the decidua. PLoS ONE 2018, 13, e0191040. [Google Scholar] [CrossRef] [PubMed]
- Tayade, C.; Hilchie, D.; He, H.; Fang, Y.; Moons, L.; Carmeliet, P.; Foster, R.A.; Croy, B.A. Genetic deletion of placenta growth factor in mice alters uterine NK cells. J. Immunol. 2007, 178, 4267–4275. [Google Scholar] [CrossRef]
- Lin, Y.L.; Liang, Y.C.; Chiang, B.L. Placental growth factor down-regulates type 1 T helper immune response by modulating the function of dendritic cells. J. Leukoc. Biol. 2007, 82, 1473–1480. [Google Scholar] [CrossRef]
- Fan, X.; Krieg, S.; Kuo, C.J.; Wiegand, S.J.; Rabinovitch, M.; Druzin, M.L.; Brenner, R.M.; Giudice, L.C.; Nayak, N.R. VEGF blockade inhibits angiogenesis and reepithelialization of endometrium. Faseb J. 2008, 22, 3571–3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockwell, L.C.; Pillai, S.; Olson, C.E.; Koos, R.D. Inhibition of vascular endothelial growth factor/vascular permeability factor action blocks estrogen-induced uterine edema and implantation in rodents. Biol. Reprod. 2002, 67, 1804–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteleone, P.; Giovanni Artini, P.; Simi, G.; Casarosa, E.; Cela, V.; Genazzani, A.R. Follicular fluid VEGF levels directly correlate with perifollicular blood flow in normoresponder patients undergoing IVF. J. Assist. Reprod. Genet. 2008, 25, 183–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, R.; Okuda, K. Multiple roles of hypoxia in bovine corpus luteum. J. Reprod. Dev. 2020, 66, 307–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauli, S.A.; Tang, H.; Wang, J.; Bohlen, P.; Posser, R.; Hartman, T.; Sauer, M.V.; Kitajewski, J.; Zimmermann, R.C. The vascular endothelial growth factor (VEGF)/VEGF receptor 2 pathway is critical for blood vessel survival in corpora lutea of pregnancy in the rodent. Endocrinology 2005, 146, 1301–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijayagunawardane, M.P.; Kodithuwakku, S.P.; Yamamoto, D.; Miyamoto, A. Vascular endothelial growth factor system in the cow oviduct: A possible involvement in the regulation of oviductal motility and embryo transport. Mol. Reprod. Dev. 2005, 72, 511–520. [Google Scholar] [CrossRef]
- Lam, P.M.; Briton-Jones, C.; Cheung, C.K.; Lok, I.H.; Yuen, P.M.; Cheung, L.P.; Haines, C. Vascular endothelial growth factor in the human oviduct: Localization and regulation of messenger RNA expression in vivo. Biol. Reprod. 2003, 68, 1870–1876. [Google Scholar] [CrossRef]
- Lam, P.M.; Briton-Jones, C.; Cheung, C.K.; Po, L.S.; Cheung, L.P.; Haines, C. Increased mRNA expression of vascular endothelial growth factor and its receptor (flt-1) in the hydrosalpinx. Hum. Reprod. 2003, 18, 2264–2269. [Google Scholar] [CrossRef] [Green Version]
- Binder, N.K.; Evans, J.; Gardner, D.K.; Salamonsen, L.A.; Hannan, N.J. Endometrial signals improve embryo outcome: Functional role of vascular endothelial growth factor isoforms on embryo development and implantation in mice. Hum. Reprod. 2014, 29, 2278–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anchordoquy, J.M.; Anchordoquy, J.P.; Testa, J.A.; Sirini, M.; Furnus, C.C. Influence of vascular endothelial growth factor and Cysteamine on in vitro bovine oocyte maturation and subsequent embryo development. Cell Biol. Int. 2015, 39, 1090–1098. [Google Scholar] [CrossRef]
- Kapiteijn, K.; Koolwijk, P.; van der Weiden, R.M.; van Nieuw Amerongen, G.; Plaisier, M.; van Hinsbergh, V.W.; Helmerhorst, F.M. Human embryo-conditioned medium stimulates in vitro endometrial angiogenesis. Fertil. Steril. 2006, 85 (Suppl. 1), 1232–1239. [Google Scholar] [CrossRef]
- Soldi, R.; Mitola, S.; Strasly, M.; Defilippi, P.; Tarone, G.; Bussolino, F. Role of alphavbeta3 integrin in the activation of vascular endothelial growth factor receptor-2. Embo J. 1999, 18, 882–892. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J.; Antczak, M.; Schrader, R. The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: Association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum. Reprod. 1997, 12, 1047–1055. [Google Scholar] [CrossRef]
- Xu, F.; Hazzard, T.M.; Evans, A.; Charnock-Jones, S.; Smith, S.; Stouffer, R.L. Intraovarian actions of anti-angiogenic agents disrupt periovulatory events during the menstrual cycle in monkeys. Contraception 2005, 71, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Wulff, C.; Wiegand, S.J.; Saunders, P.T.; Scobie, G.A.; Fraser, H.M. Angiogenesis during follicular development in the primate and its inhibition by treatment with truncated Flt-1-Fc (vascular endothelial growth factor Trap(A40)). Endocrinology 2001, 142, 3244–3254. [Google Scholar] [CrossRef] [PubMed]
- Hazzard, T.M.; Molskness, T.A.; Chaffin, C.L.; Stouffer, R.L. Vascular endothelial growth factor (VEGF) and angiopoietin regulation by gonadotrophin and steroids in macaque granulosa cells during the peri-ovulatory interval. Mol. Hum. Reprod. 1999, 5, 1115–1121. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.W.; Scaramuzzi, R.J.; Wheeler-Jones, C.P.; Khalid, M. The expression of angiogenic growth factors and their receptors in ovarian follicles throughout the estrous cycle in the ewe. Theriogenology 2010, 73, 856–872. [Google Scholar] [CrossRef] [PubMed]
- Friedman, C.I.; Seifer, D.B.; Kennard, E.A.; Arbogast, L.; Alak, B.; Danforth, D.R. Elevated level of follicular fluid vascular endothelial growth factor is a marker of diminished pregnancy potential. Fertil. Steril. 1998, 70, 836–839. [Google Scholar] [CrossRef]
- Ocal, P.; Aydin, S.; Cepni, I.; Idil, S.; Idil, M.; Uzun, H.; Benian, A. Follicular fluid concentrations of vascular endothelial growth factor, inhibin A and inhibin B in IVF cycles: Are they markers for ovarian response and pregnancy outcome? Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Artini, P.G.; Ruggiero, M.; Parisen Toldin, M.R.; Monteleone, P.; Monti, M.; Cela, V.; Genazzani, A.R. Vascular endothelial growth factor and its soluble receptor in patients with polycystic ovary syndrome undergoing IVF. Hum. Fertil. 2009, 12, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Young, F.M.; Rodger, F.E.; Illingworth, P.J.; Fraser, H.M. Cell proliferation and vascular morphology in the marmoset corpus luteum. Hum. Reprod. 2000, 15, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Wada, Y.; Ozaki, H.; Abe, N.; Mori, A.; Sakamoto, K.; Nagamitsu, T.; Nakahara, T.; Ishii, K. Role of vascular endothelial growth factor in maintenance of pregnancy in mice. Endocrinology 2013, 154, 900–910. [Google Scholar] [CrossRef]
- Woad, K.J.; Hunter, M.G.; Mann, G.E.; Laird, M.; Hammond, A.J.; Robinson, R.S. Fibroblast growth factor 2 is a key determinant of vascular sprouting during bovine luteal angiogenesis. Reproduction 2012, 143, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwu, Y.M.; Chen, C.P.; Li, S.H.; Chen, P.W.; Lee, R.K.; Fleming, S. Expression of vascular endothelial growth factor messenger ribonucleic acid and protein in human preimplantation embryos. Fertil. Steril. 2006, 85, 1830–1832. [Google Scholar] [CrossRef]
- Thouas, G.A.; Dominguez, F.; Green, M.P.; Vilella, F.; Simon, C.; Gardner, D.K. Soluble ligands and their receptors in human embryo development and implantation. Endocr. Rev. 2015, 36, 92–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baston-Buest, D.M.; Kruessel, J.S.; Ingmann, S.C.; Hirchenhain, J.; Janni, W.; Hess, A.P. In vitro culture does not alter the expression of vascular endothelial growth factor and its receptors in single murine preimplantation embryos. Gynecol. Obstet. Investig. 2011, 71, 24–31. [Google Scholar] [CrossRef]
- Licht, P.; Russu, V.; Wildt, L. On the role of human chorionic gonadotropin (hCG) in the embryo-endometrial microenvironment: Implications for differentiation and implantation. Semin. Reprod. Med. 2001, 19, 37–47. [Google Scholar] [CrossRef]
- Lessey, B.A. Adhesion molecules and implantation. J. Reprod. Immunol. 2002, 55, 101–112. [Google Scholar] [CrossRef]
- Massimiani, M.; Lacconi, V.; La Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular Signaling Regulating Endometrium-Blastocyst Crosstalk. Int. J. Mol. Sci. 2019, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, C.; Ledger, W.; Wang, Q.; Liu, F.; Demirol, A.; Gurgan, T.; Cutting, R.; Ong, K.; Sallam, H.; Li, T.C. Recurrent implantation failure: Definition and management. Reprod. Biomed. Online 2014, 28, 14–38. [Google Scholar] [CrossRef] [Green Version]
- Bashiri, A.; Halper, K.I.; Orvieto, R. Recurrent Implantation Failure-update overview on etiology, diagnosis, treatment and future directions. Reprod. Biol. Endocrinol. RbE 2018, 16, 121. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Man, G.C.W.; Liu, Y.; Wu, F.; Huang, J.; Li, T.C.; Wang, C.C. Physiological and pathological angiogenesis in endometrium at the time of embryo implantation. Am. J. Reprod. Immunol. 2017, 78, e12693. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Ford, B.; Bhaskaran, S.; Thum, M.; Bansal, A. Elevated Levels of Serum Vascular Endothelial Growth Factor-A Are Not Related to NK Cell Parameters in Recurrent IVF Failure. J. Reprod. Infertil. 2017, 18, 280–287. [Google Scholar]
- Boudjenah, R.; Molina-Gomes, D.; Torre, A.; Boitrelle, F.; Taieb, S.; Dos Santos, E.; Wainer, R.; de Mazancourt, P.; Selva, J.; Vialard, F. Associations between Individual and Combined Polymorphisms of the TNF and VEGF Genes and the Embryo Implantation Rate in Patients Undergoing In Vitro Fertilization (IVF) Programs. PLoS ONE 2014, 9, e108287. [Google Scholar] [CrossRef]
- Turienzo, A.; Lledó, B.; Ortiz, J.A.; Morales, R.; Sanz, J.; Llácer, J.; Bernabeu, R. Prevalence of candidate single nucleotide polymorphisms on p53, IL-11, IL-10, VEGF and APOE in patients with repeated implantation failure (RIF) and pregnancy loss (RPL). Hum. Fertil. 2020, 23, 117–122. [Google Scholar] [CrossRef]
- Vagnini, L.D.; Nascimento, A.M.; Canas Mdo, C.; Renzi, A.; Oliveira-Pelegrin, G.R.; Petersen, C.G.; Mauri, A.L.; Oliveira, J.B.; Baruffi, R.L.; Cavagna, M.; et al. The Relationship between Vascular Endothelial Growth Factor 1154G/A Polymorphism and Recurrent Implantation Failure. Med. Princ. Pr. 2015, 24, 533–537. [Google Scholar] [CrossRef]
- Jung, Y.W.; Kim, J.O.; Rah, H.; Kim, J.H.; Kim, Y.R.; Lee, Y.; Lee, W.S.; Kim, N.K. Genetic variants of vascular endothelial growth factor are associated with recurrent implantation failure in Korean women. Reprod. Biomed. Online 2016, 32, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Goodman, C.; Jeyendran, R.S.; Coulam, C.B. P53 tumor suppressor factor, plasminogen activator inhibitor, and vascular endothelial growth factor gene polymorphisms and recurrent implantation failure. Fertil. Steril. 2009, 92, 494–498. [Google Scholar] [CrossRef]
- Royal College of Obstetricians and Gynaecologists. (Third ed.) The Investigation and Treatment of Couples with Recurrent First-Trimester and Second-Trimester Miscarriage. Green-Top Guideline, 2011, No. 17. Available online: https://www.rcog.org.uk/en/guidelines-research-services/guidelines/gtg17 (accessed on 30 December 2020).
- Li, T.C.; Makris, M.; Tomsu, M.; Tuckerman, E.; Laird, S. Recurrent miscarriage: Aetiology, management and prognosis. Hum. Reprod. Update 2002, 8, 463–481. [Google Scholar] [CrossRef]
- Amirchaghmaghi, E.; Rezaei, A.; Moini, A.; Roghaei, M.A.; Hafezi, M.; Aflatoonian, R. Gene expression analysis of VEGF and its receptors and assessment of its serum level in unexplained recurrent spontaneous abortion. Cell J. 2015, 16, 538–545. [Google Scholar]
- Banerjee, P.; Jana, S.K.; Pasricha, P.; Ghosh, S.; Chakravarty, B.; Chaudhury, K. Proinflammatory cytokines induced altered expression of cyclooxygenase-2 gene results in unreceptive endometrium in women with idiopathic recurrent spontaneous miscarriage. Fertil. Steril. 2013, 99, 179–187.e172. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, L.; Wang, C.C.; Huang, J.; Li, T.C. Hypoxia inducible factor and microvessels in peri-implantation endometrium of women with recurrent miscarriage. Fertil. Steril. 2016, 105, 1496–1502.e1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, L.H.; Li, M.J.; Li, M.Q.; Yang, D.M.; Shi, L. Vascular endothelial growth factor (VEGF) and the VEGF soluble receptor-1 (sFlt-1) in chorionic villus tissue from Chinese women with early recurrent spontaneous abortion. J. Int. Med. Res. 2011, 39, 830–837. [Google Scholar] [CrossRef]
- He, X.; Chen, Q. Reduced expressions of connexin 43 and VEGF in the first-trimester tissues from women with recurrent pregnancy loss. Reprod. Biol. Endocrinol. 2016, 14, 46. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.K.; Choi, B.C.; Lee, S.H.; Kim, J.W.; Cha, K.Y.; Baek, K.H. Expression of angiogenesis- and apoptosis-related genes in chorionic villi derived from recurrent pregnancy loss patients. Mol. Reprod. Dev. 2003, 66, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Al-Khateeb, G.M.; Mustafa, F.E.; Sater, M.S.; Almawi, W.Y. Effect of the functional VEGFA-583C/T variant on vascular endothelial growth factor levels and the risk of recurrent spontaneous miscarriage. Fertil. Steril. 2011, 95, 2471–2473. [Google Scholar] [CrossRef]
- Bagheri, A.; Kumar, P.; Kamath, A.; Rao, P. Association of angiogenic cytokines (VEGF-A and VEGF-C) and clinical characteristic in women with unexplained recurrent miscarriage. Bratisl. Lek. Listy 2017, 118, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Almawi, W.Y.; Saldanha, F.L.; Mahmood, N.A.; Al-Zaman, I.; Sater, M.S.; Mustafa, F.E. Relationship between VEGFA polymorphisms and serum VEGF protein levels and recurrent spontaneous miscarriage. Hum. Reprod. 2013, 28, 2628–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, C.J.; Webb, N.J.; Bottomley, M.J.; Brenchley, P.E. Identification of polymorphisms within the vascular endothelial growth factor (VEGF) gene: Correlation with variation in VEGF protein production. Cytokine 2000, 12, 1232–1235. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Du, C.; Li, H.; Du, J.; Yan, X.; Peng, L.; Li, G.; Chen, Z.J. Association of VEGF genetic polymorphisms with recurrent spontaneous abortion risk: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0123696. [Google Scholar] [CrossRef]
- Su, M.T.; Lin, S.H.; Lee, I.W.; Chen, Y.C.; Kuo, P.L. Association of polymorphisms/haplotypes of the genes encoding vascular endothelial growth factor and its KDR receptor with recurrent pregnancy loss. Hum. Reprod. 2011, 26, 758–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xiong, W.; Xiong, Y.; Liu, H.; Liu, Y. 17 β-Estradiol promotes vascular endothelial growth factor expression via the Wnt/β-catenin pathway during the pathogenesis of endometriosis. Mol. Hum. Reprod. 2016, 22, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Lian, F.; Liu, H.P.; Wang, Y.X.; Zhang, J.W.; Sun, Z.G.; Ma, F.M.; Zhang, N.; Liu, Y.H.; Meng, Q. Expressions of VEGF and Ki-67 in eutopic endometrium of patients with endometriosis and effect of Quyu Jiedu Recipe on VEGF expression. Chin. J. Integr. Med. 2007, 13, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Lupicka, M.; Zadroga, A.; Szczepańska, A.; Korzekwa, A.J. Effect of ovarian steroids on vascular endothelial growth factor a expression in bovine uterine endothelial cells during adenomyosis. BMC Vet. Res. 2019, 15, 473. [Google Scholar] [CrossRef]
- Kim, C.H.; Ahn, J.W.; Kim, S.H.; Chae, H.D.; Kang, B.M. Effects on in vitro fertilization-embryo transfer outcomes of vascular endothelial growth factor receptor-1, -2 and -3 in eutopic endometrial tissue of women with endometriosis. J. Obs. Gynaecol. Res. 2011, 37, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, S.; Yang, F.; Sun, Y.; Niu, L.; Ren, Y.; Wang, H.; He, Y.; Du, J.; Yang, J.; et al. Biological characteristics of endometriotic mesenchymal stem cells isolated from ectopic lesions of patients with endometriosis. Stem Cell Res. Ther. 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xin, X.; Hua, T.; Shi, R.; Chi, S.; Jin, Z.; Wang, H. Efficacy of Anti-VEGF/VEGFR Agents on Animal Models of Endometriosis: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0166658. [Google Scholar]
- Ferrara, N.; Frantz, G.; LeCouter, J.; Dillard-Telm, L.; Pham, T.; Draksharapu, A.; Giordano, T.; Peale, F. Differential expression of the angiogenic factor genes vascular endothelial growth factor (VEGF) and endocrine gland-derived VEGF in normal and polycystic human ovaries. Am. J. Pathol. 2003, 162, 1881–1893. [Google Scholar] [CrossRef] [Green Version]
- Tal, R.; Seifer, D.B.; Arici, A. The emerging role of angiogenic factor dysregulation in the pathogenesis of polycystic ovarian syndrome. Semin. Reprod. Med. 2015, 33, 195–207. [Google Scholar]
- Di Pietro, M.; Pascuali, N.; Parborell, F.; Abramovich, D. Ovarian angiogenesis in polycystic ovary syndrome. Reproduction 2018, 155, R199–R209. [Google Scholar] [CrossRef] [Green Version]
- Kamat, B.R.; Brown, L.F.; Manseau, E.J.; Senger, D.R.; Dvorak, H.F. Expression of vascular permeability factor/vascular endothelial growth factor by human granulosa and theca lutein cells. Role in corpus luteum development. Am. J. Pathol. 1995, 146, 157–165. [Google Scholar] [PubMed]
- Agrawal, R.; Sladkevicius, P.; Engmann, L.; Conway, G.S.; Payne, N.N.; Bekis, J.; Tan, S.L.; Campbell, S.; Jacobs, H.S. Serum vascular endothelial growth factor concentrations and ovarian stromal blood flow are increased in women with polycystic ovaries. Hum. Reprod. 1998, 13, 651–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, C.; Artini, P.G.; D’Ambrogio, G.; Genazzani, A.D.; Genazzani, A.R. The role of color Doppler imaging in the diagnosis of polycystic ovary syndrome. Am. J. Obs. Gynecol 1995, 172, 108–113. [Google Scholar] [CrossRef]
- Zaidi, J.; Campbell, S.; Pittrof, R.; Kyei-Mensah, A.; Shaker, A.; Jacobs, H.S.; Tan, S.L. Ovarian stromal blood flow in women with polycystic ovaries--a possible new marker for diagnosis? Hum. Reprod. 1995, 10, 1992–1996. [Google Scholar] [CrossRef]
- Abramovich, D.; Irusta, G.; Bas, D.; Cataldi, N.I.; Parborell, F.; Tesone, M. Angiopoietins/TIE2 system and VEGF are involved in ovarian function in a DHEA rat model of polycystic ovary syndrome. Endocrinology 2012, 153, 3446–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tal, R.; Seifer, D.B.; Grazi, R.V.; Malter, H.E. Follicular fluid placental growth factor is increased in polycystic ovarian syndrome: Correlation with ovarian stimulation. Reprod. Biol. Endocrinol. 2014, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Qu, Q.; Dai, H.; Liu, Y.; Jiang, L.; Huang, X.; Hao, C. Effects of hypoxia-inducible factor-1α on endometrial receptivity of women with polycystic ovary syndrome. Mol. Med. Rep. 2018, 17, 414–421. [Google Scholar] [CrossRef]
- Lee, E.J.; Oh, B.; Lee, J.Y.; Kimm, K.; Park, J.M.; Baek, K.H. Association study between single nucleotide polymorphisms in the VEGF gene and polycystic ovary syndrome. Fertil. Steril. 2008, 89, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Guruvaiah, P.; Govatati, S.; Reddy, T.V.; Lomada, D.; Deenadayal, M.; Shivaji, S.; Bhanoori, M. The VEGF +405 G>C 5′ untranslated region polymorphism and risk of PCOS: A study in the South Indian Women. J. Assist. Reprod. Genet. 2014, 31, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istrate, M.; Mihu, C.; Şuşman, S.; Melincovici, C.S.; Măluţan, A.M.; Buiga, R.; Bolboacă, S.D.; Mihu, C.M. Highlighting the R1 and R2 VEGF receptors in placentas resulting from normal development pregnancies and from pregnancies complicated by preeclampsia. Rom. J. Morphol. Embryol. 2018, 59, 139–146. [Google Scholar]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Jakovljevic, A.; Bogavac, M.; Lozanov-Crvenkovic, Z.; Milosević-Tosic, M.; Nikolic, A.; Mitic, G. Early pregnancy angiogenic proteins levels and pregnancy related hypertensive disorders. J. Matern.-Fetal Neonatal Med. 2017, 30, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; McMaster, M.; Woo, K.; Janatpour, M.; Perry, J.; Karpanen, T.; Alitalo, K.; Damsky, C.; Fisher, S.J. Vascular endothelial growth factor ligands and receptors that regulate human cytotrophoblast survival are dysregulated in severe preeclampsia and hemolysis, elevated liver enzymes, and low platelets syndrome. Am. J. Pathol. 2002, 160, 1405–1423. [Google Scholar] [CrossRef] [Green Version]
- Mochan, S.; Dhingra, M.K.; Gupta, S.K.; Saxena, S.; Arora, P.; Yadav, V.; Rani, N.; Luthra, K.; Dwivedi, S.; Bhatla, N.; et al. Status of VEGF in preeclampsia and its effect on endoplasmic reticulum stress in placental trophoblast cells. Eur. J. Obs. Gynecol. Reprod. Biol. X 2019, 4, 100070. [Google Scholar] [CrossRef]
- Weed, S.; Bastek, J.A.; Anton, L.; Elovitz, M.A.; Parry, S.; Srinivas, S.K. Examining the correlation between placental and serum placenta growth factor in preeclampsia. Am. J. Obs. Gynecol. 2012, 207, e141–e146. [Google Scholar] [CrossRef]
- Ferris, T.F. Pregnancy, preeclampsia, and the endothelial cell. N. Engl. J. Med. 1991, 325, 1439–1440. [Google Scholar] [CrossRef]
- Roberts, J.M.; Taylor, R.N.; Musci, T.J.; Rodgers, G.M.; Hubel, C.A.; McLaughlin, M.K. Preeclampsia: An endothelial cell disorder. Am. J. Obs. Gynecol. 1989, 161, 1200–1204. [Google Scholar] [CrossRef]
- Runyan, C.L.; McIntosh, S.Z.; Maestas, M.M.; Quinn, K.E.; Boren, B.P.; Ashley, R.L. CXCR4 signaling at the ovine fetal-maternal interface regulates vascularization, CD34+ cell presence, and autophagy in the endometrium. Biol. Reprod. 2019, 101, 102–111. [Google Scholar] [CrossRef]
- Lala, P.K.; Nandi, P. Mechanisms of trophoblast migration, endometrial angiogenesis in preeclampsia: The role of decorin. Cell Adhes. Migr. 2016, 10, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhu, H.; Klausen, C.; Peng, B.; Leung, P.C. Vascular Endothelial Growth Factor-A (VEGF-A) Mediates Activin A-Induced Human Trophoblast Endothelial-Like Tube Formation. Endocrinology 2015, 156, 4257–4268. [Google Scholar] [CrossRef] [Green Version]
- Hornig, C.; Barleon, B.; Ahmad, S.; Vuorela, P.; Ahmed, A.; Weich, H.A. Release and complex formation of soluble VEGFR-1 from endothelial cells and biological fluids. Lab. Investig. 2000, 80, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jardim, L.L.; Rios, D.R.; Perucci, L.O.; de Sousa, L.P.; Gomes, K.B.; Dusse, L.M. Is the imbalance between pro-angiogenic and anti-angiogenic factors associated with preeclampsia? Clin. Chim. Acta Int. J. Clin. Chem. 2015, 447, 34–38. [Google Scholar] [CrossRef]
- Nikuei, P.; Malekzadeh, K.; Rajaei, M.; Nejatizadeh, A.; Ghasemi, N. The imbalance in expression of angiogenic and anti-angiogenic factors as candidate predictive biomarker in preeclampsia. Iran. J. Reprod. Med. 2015, 13, 251–262. [Google Scholar] [PubMed]
- Kar, M. Role of biomarkers in early detection of preeclampsia. J. Clin. Diagn. Res. 2014, 8, Be01–Be04. [Google Scholar] [CrossRef]
- Ruckman, J.; Green, L.S.; Beeson, J.; Waugh, S.; Gillette, W.L.; Henninger, D.D.; Claesson-Welsh, L.; Janjić, N. 2′-Fluoropyrimidine RNA-based aptamers to the 165-amino acid form of vascular endothelial growth factor (VEGF165). Inhibition of receptor binding and VEGF-induced vascular permeability through interactions requiring the exon 7-encoded domain. J. Biol. Chem. 1998, 273, 20556–20567. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Li, J.J.; Bay, B.H.; Yung, L.Y. Investigating the antiproliferative activity of high affinity DNA aptamer on cancer cells. PLoS ONE 2013, 8, e50964. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltz, L.B.; Clarke, S.; Díaz-Rubio, E.; Scheithauer, W.; Figer, A.; Wong, R.; Koski, S.; Lichinitser, M.; Yang, T.S.; Rivera, F.; et al. Bevacizumab in combination with oxaliplatin-based chemotherapy as first-line therapy in metastatic colorectal cancer: A randomized phase III study. J. Clin. Oncol. 2008, 26, 2013–2019. [Google Scholar] [CrossRef] [Green Version]
- Tebbutt, N.C.; Wilson, K.; Gebski, V.J.; Cummins, M.M.; Zannino, D.; van Hazel, G.A.; Robinson, B.; Broad, A.; Ganju, V.; Ackland, S.P.; et al. Capecitabine, bevacizumab, and mitomycin in first-line treatment of metastatic colorectal cancer: Results of the Australasian Gastrointestinal Trials Group Randomized Phase III MAX Study. J. Clin. Oncol. 2010, 28, 3191–3198. [Google Scholar] [CrossRef]
- Reck, M.; von Pawel, J.; Zatloukal, P.; Ramlau, R.; Gorbounova, V.; Hirsh, V.; Leighl, N.; Mezger, J.; Archer, V.; Moore, N.; et al. Phase III trial of cisplatin plus gemcitabine with either placebo or bevacizumab as first-line therapy for nonsquamous non-small-cell lung cancer: AVAil. J. Clin. Oncol. 2009, 27, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Sandler, A.; Gray, R.; Perry, M.C.; Brahmer, J.; Schiller, J.H.; Dowlati, A.; Lilenbaum, R.; Johnson, D.H. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. N. Engl. J. Med. 2006, 355, 2542–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, N.A.; Bracha, P.; Hussain, R.M.; Morral, N.; Ciulla, T.A. Gene therapy for age-related macular degeneration. Expert Opin. Biol. 2017, 17, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of vascular endothelial growth factor (VEGF) in the human endometrium and interaction between endometrium and embryo.
Figure 1.
Role of vascular endothelial growth factor (VEGF) in the human endometrium and interaction between endometrium and embryo.

Figure 2.
Role of VEGF in the development of human oocytes and embryo.


{kind=link}
{kind=link}
Table 1.
Role of VEGF in embryo implantation. PlGF, placental growth factor, VEGFR-2, VEGF receptor-2.
Table 1.
Role of VEGF in embryo implantation. PlGF, placental growth factor, VEGFR-2, VEGF receptor-2.
| VEGF Family Members | Function |
|---|---|
| Endometrial receptivity | |
| VEGF | Promotes endometrial angiogenesis and vascular permeability [59,60] |
| VEGF | Monocyte activation [54] |
| VEGF | Recruitment and activation of macrophages [32,56] |
| PlGF | Monocyte activation [53] |
| PlGF | Recruitment and activation of macrophages [55] |
| PlGF | Proliferation and differentiation of uNK cells [57] |
| PlGF | Differentiation and maturation of dendritic cells; skews type 1 T helper immune response to the Th2 phenotype [58] |
| Embryo development | |
| VEGF | Development of the perifollicular capillary network [61]. |
| VEGF | Formation of and hormone production by corpus luteum [62,63] |
| VEGF | Regulates oviduct mobility [64] |
| VEGF | Regulates oviductal fluid secretion [65,66] |
| VEGF | Increases blastocyst yield and blastocyst cell numbers, enhances blastocyst outgrowth, and reduces cavitation time [39,67,68] |
| PlGF | Increases blastocyst cell numbers and enhances blastocyst outgrowth [47] |
| Embryo implantation | |
| VEGF | Improves endometrial epithelial cells’ adhesion ability and increases implantation rates [39,67] |
| VEGF | Embryo-derived VEGF stimulates angiogenesis at the implantation site [69] |
| VEGFR-2 | Interacts with integrin αvβ3 [70] |
| PlGF | Improves endometrial epithelial cells’ adhesion ability [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, X.; Yi, H.; Li, T.C.; Wang, Y.; Wang, H.; Chen, X. Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications. Biomolecules 2021, 11, 253. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020253
AMA Style
Guo X, Yi H, Li TC, Wang Y, Wang H, Chen X. Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications. Biomolecules. 2021; 11(2):253. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020253
Chicago/Turabian StyleGuo, Xi, Hong Yi, Tin Chiu Li, Yu Wang, Huilin Wang, and Xiaoyan Chen. 2021. "Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications" Biomolecules 11, no. 2: 253. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020253
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.