

Screening for Lipid-Metabolism-Related Genes and Identifying the Diagnostic Potential of ANGPTL6 for HBV-Related Early-Stage Hepatocellular Carcinoma
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
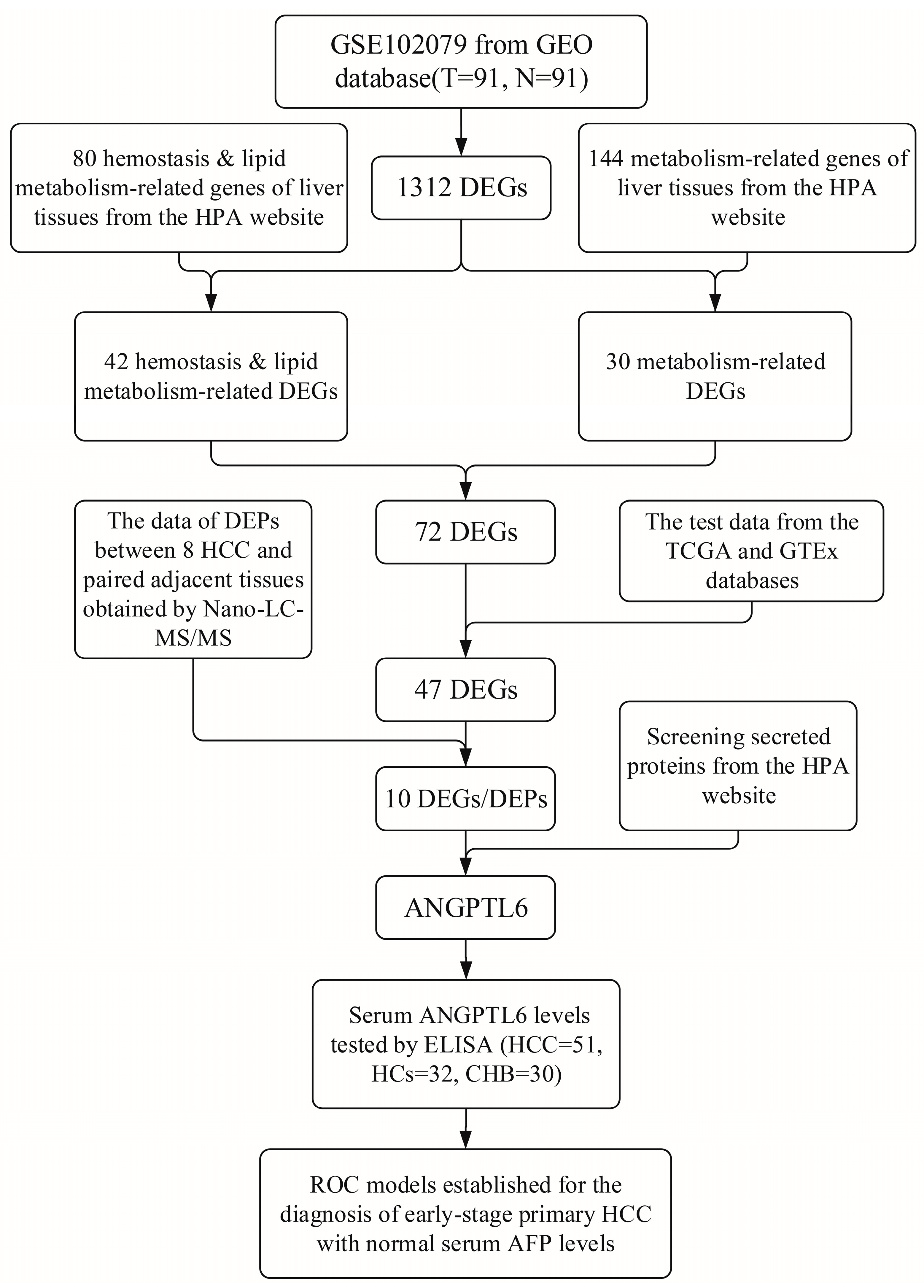
2.1. Identification of DEGs from Microarray Data
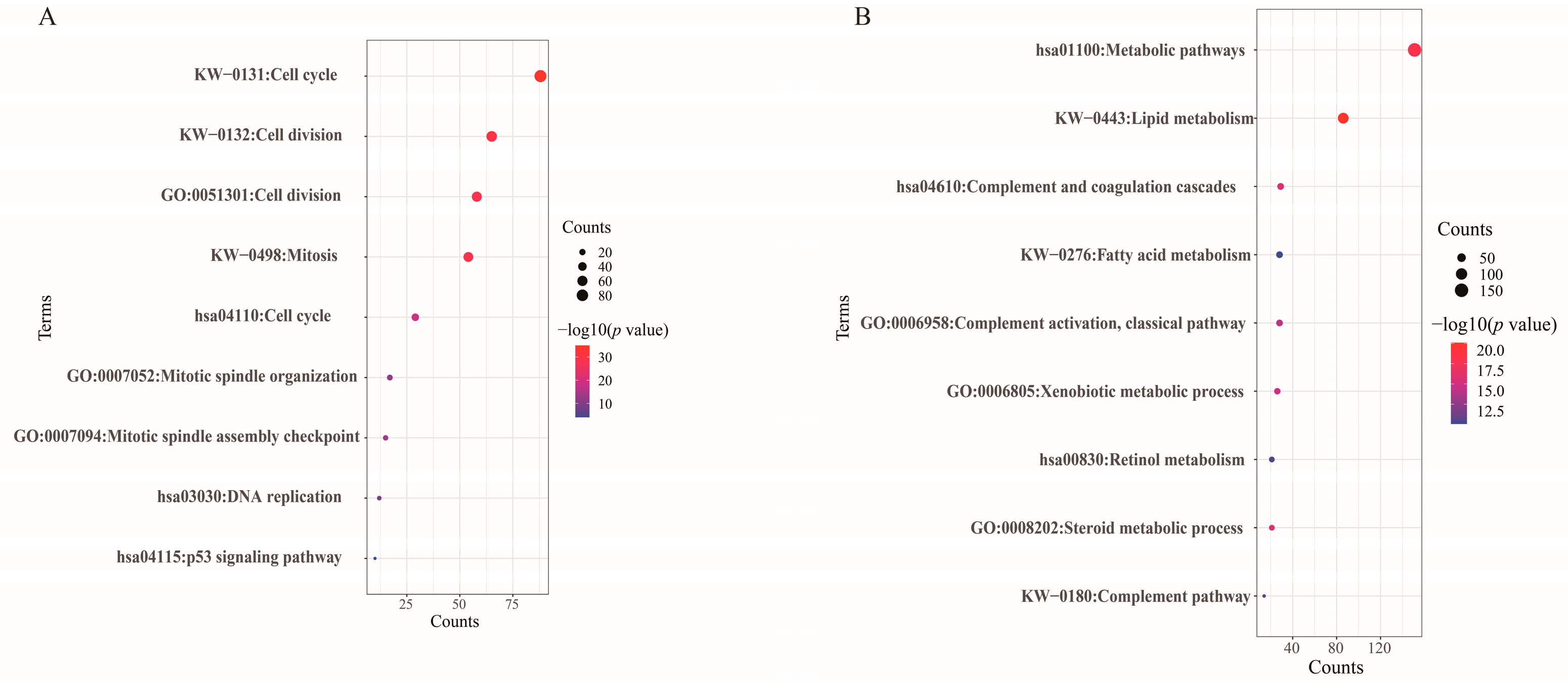
2.2. Functional Enrichment Analyses of DEGs
2.3. Gene Expression Clusters of Transcriptomics Data in Tissues
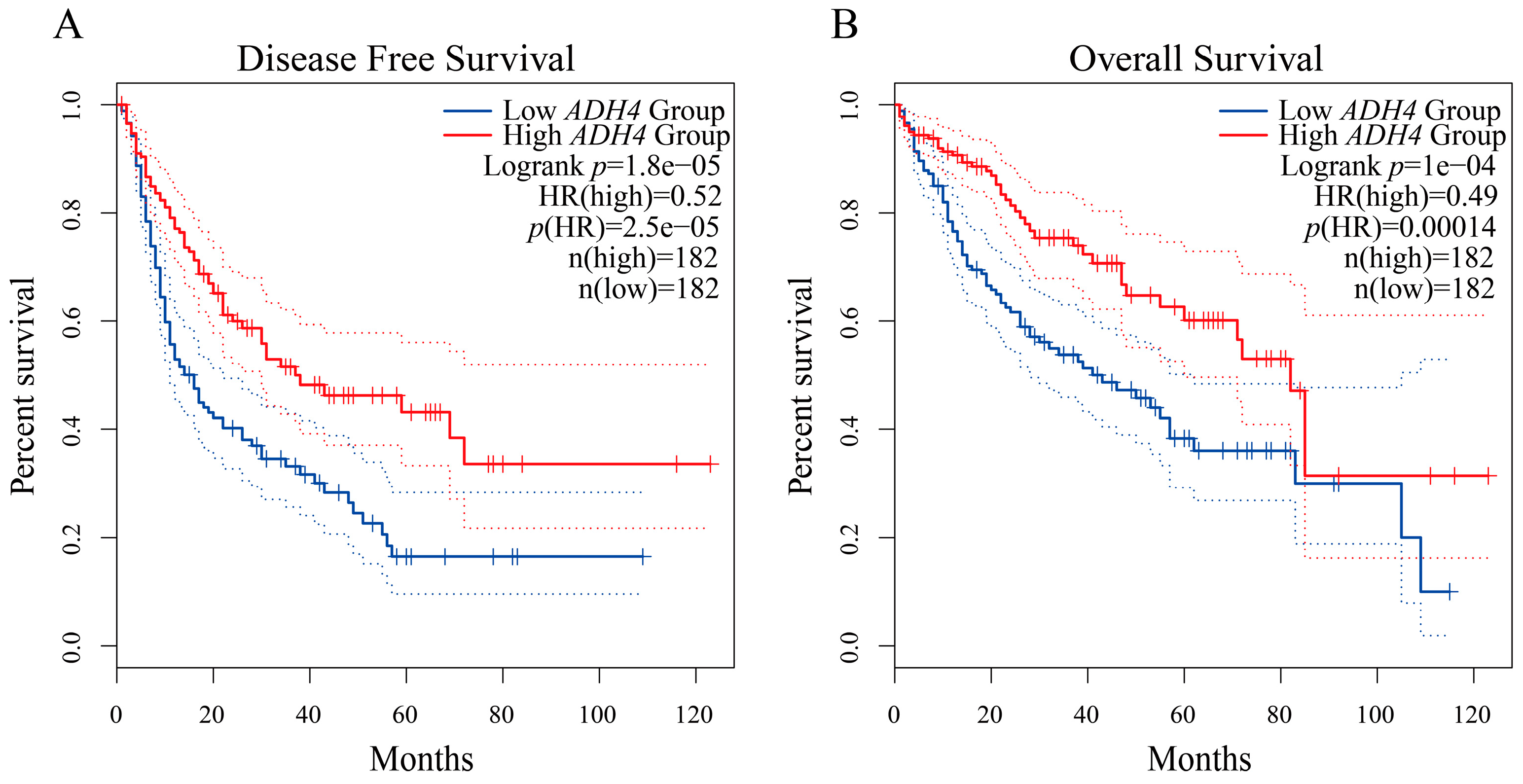
2.4. Gene Expression Profiling Interactive Analysis (GEPIA) for Validating Gene Expression and Survival Analysis
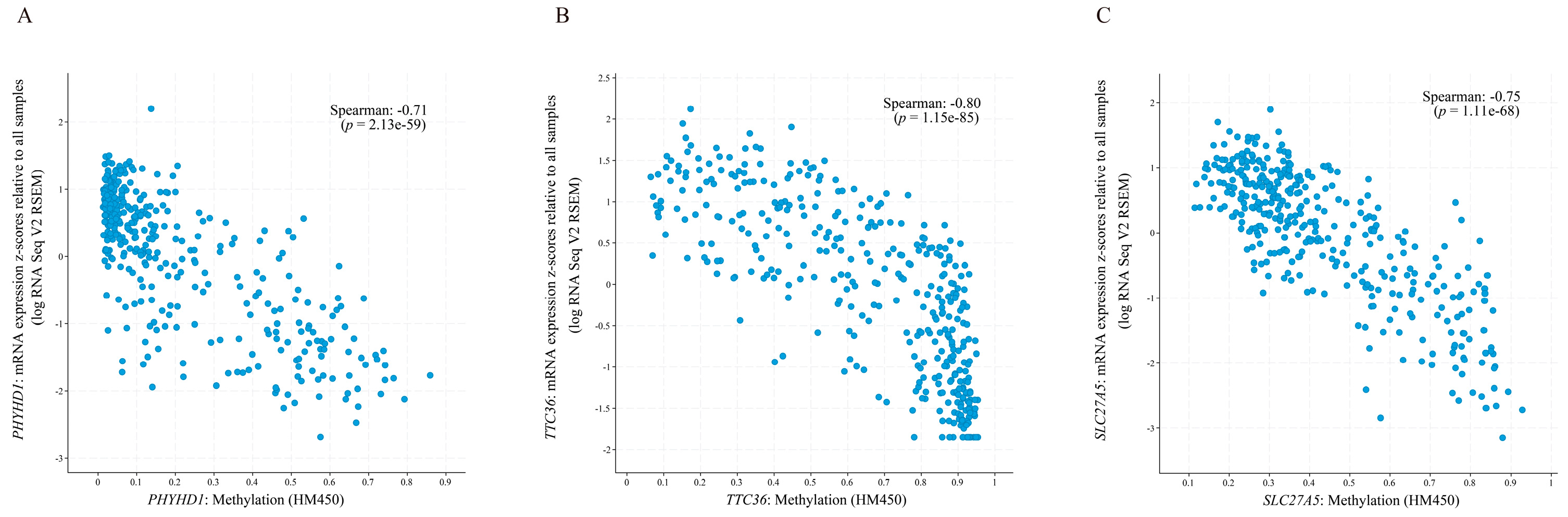
2.5. cBioPortal for Exploring the Correlation between Gene Expression and Methylation
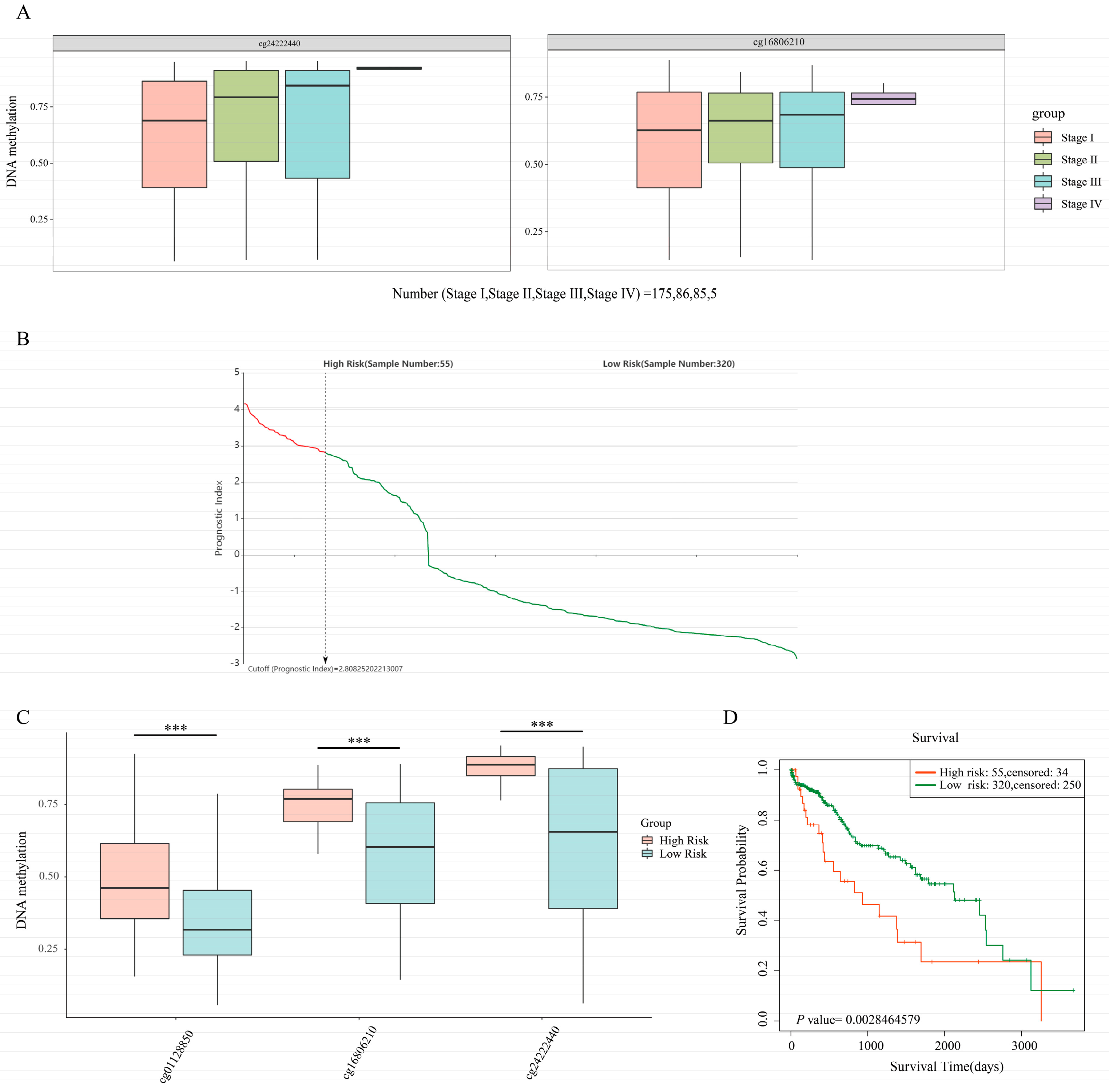
2.6. SurvivalMeth for Exploring the Correlation between DNA Methylation and the Prognosis of HCC Patients
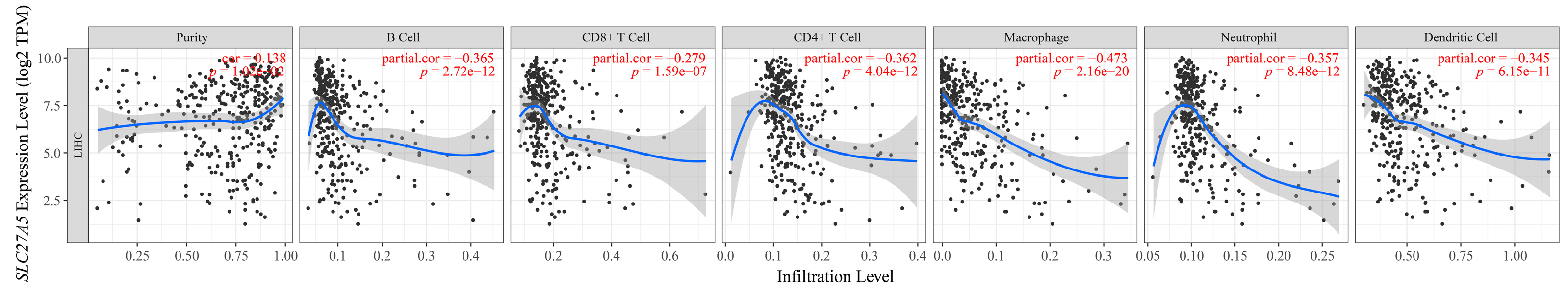
2.7. Tumor Immune Estimation Resource (TIMER) for Exploring the Correlation between Gene Expression and Immune Infiltration
2.8. Ethical Statement
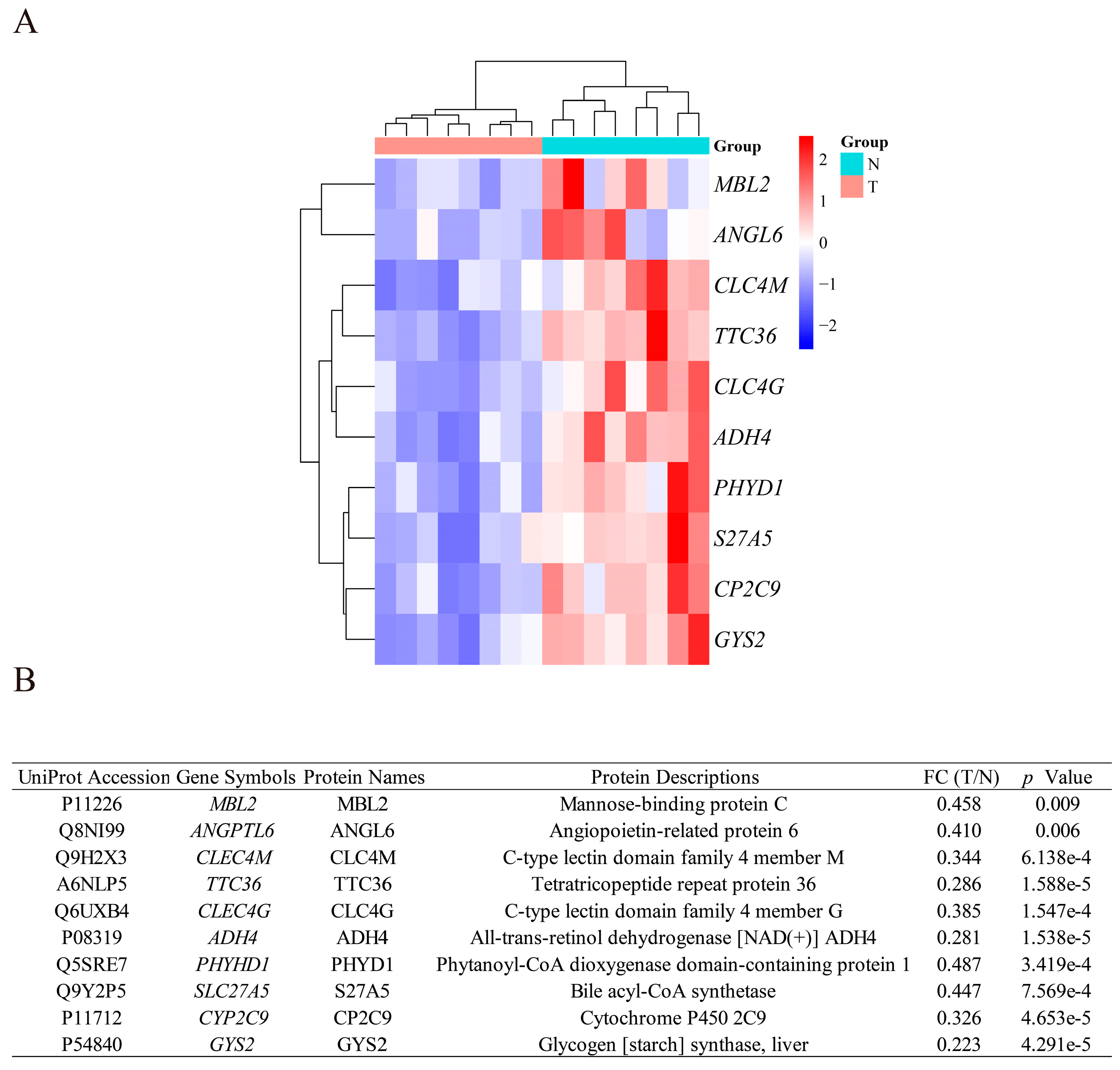
2.9. Tissue Sample Collection and Nano-LC-MS/MS Analysis
2.10. Serum Sample Collection and Storage
2.11. ELISAs of Serum ANGPTL6
2.12. 1H-Nuclear Magnetic Resonance (1H-NMR) Experiments
2.13. Statistical Analysis
3. Results
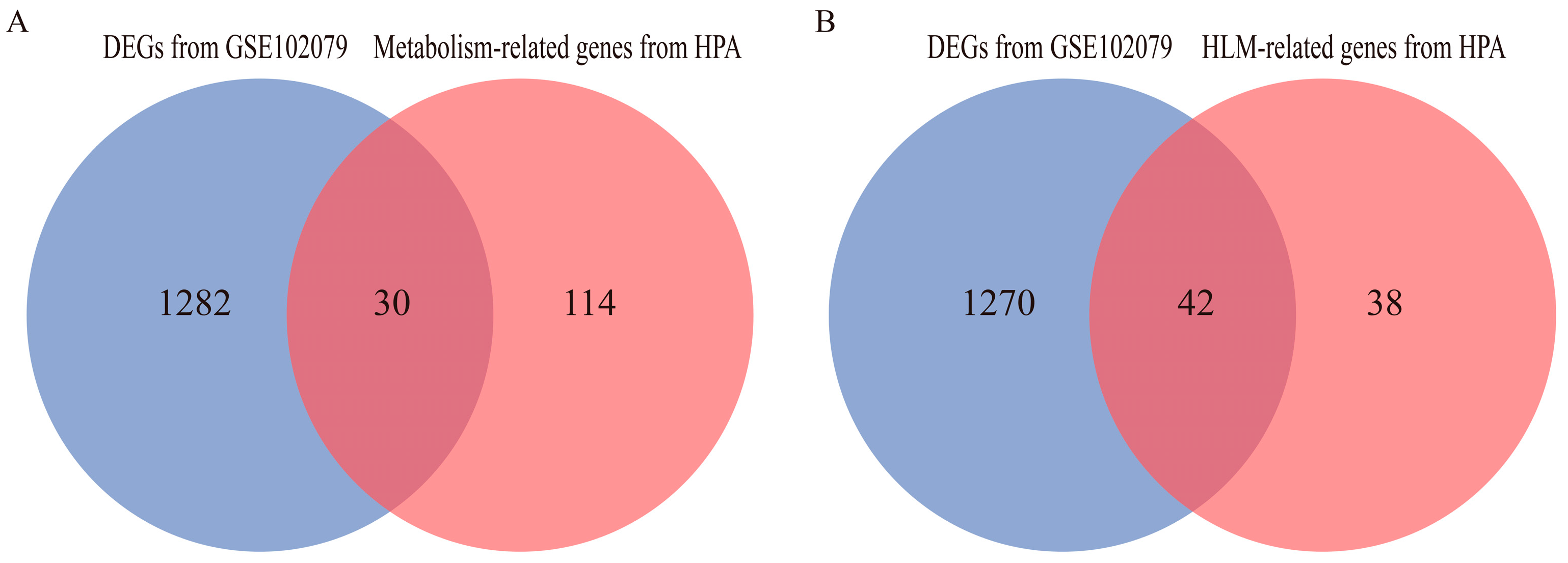
3.1. Identification of DEGs and Functional Enrichment Analyses
3.2. Identification of DEGs Related to Hemostasis and Lipid Metabolism
3.3. The mRNA Expression Levels of DEGs in the TCGA and GTEx Databases
3.4. Identification of DEPs Encoded by the DEGs
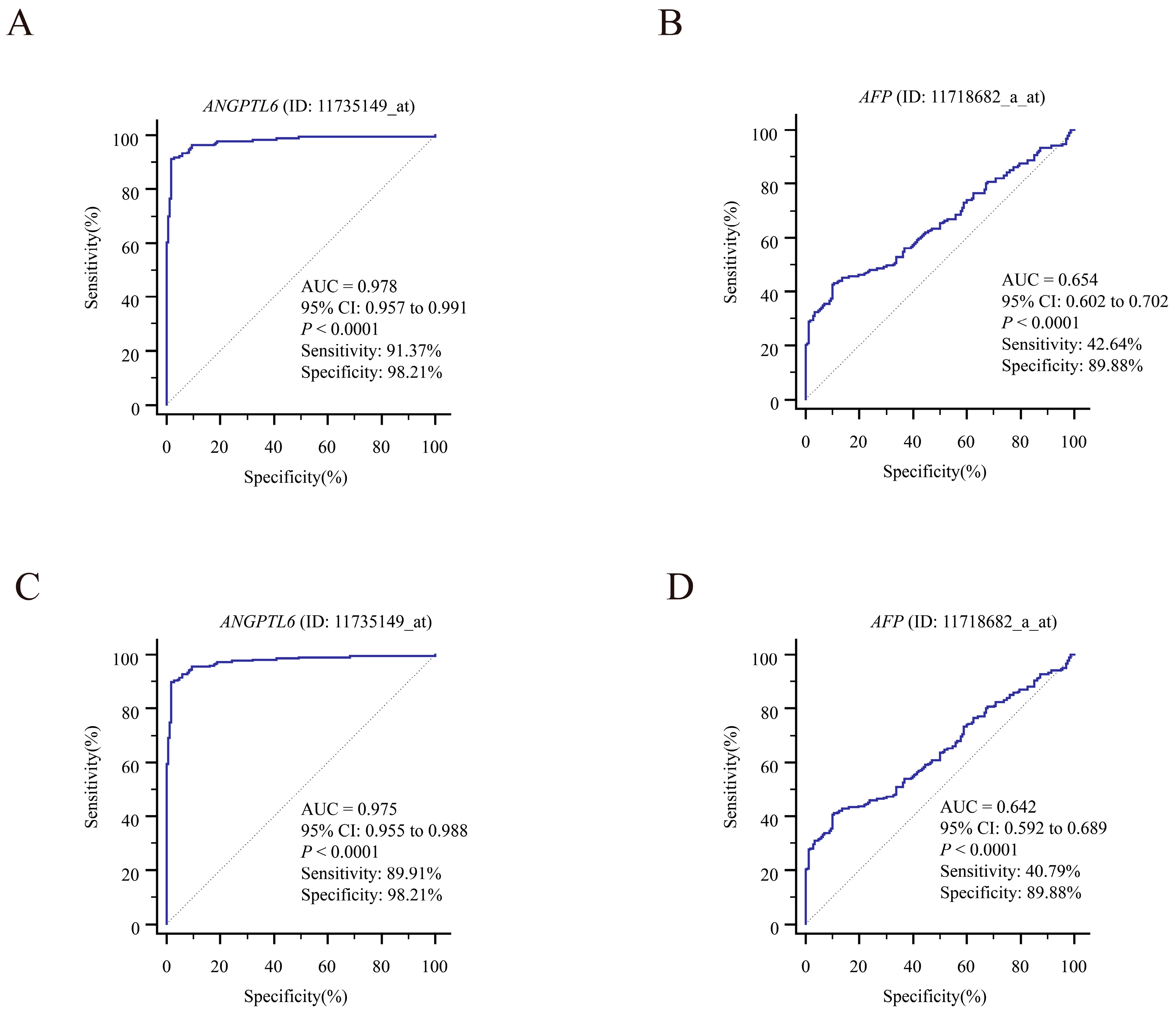
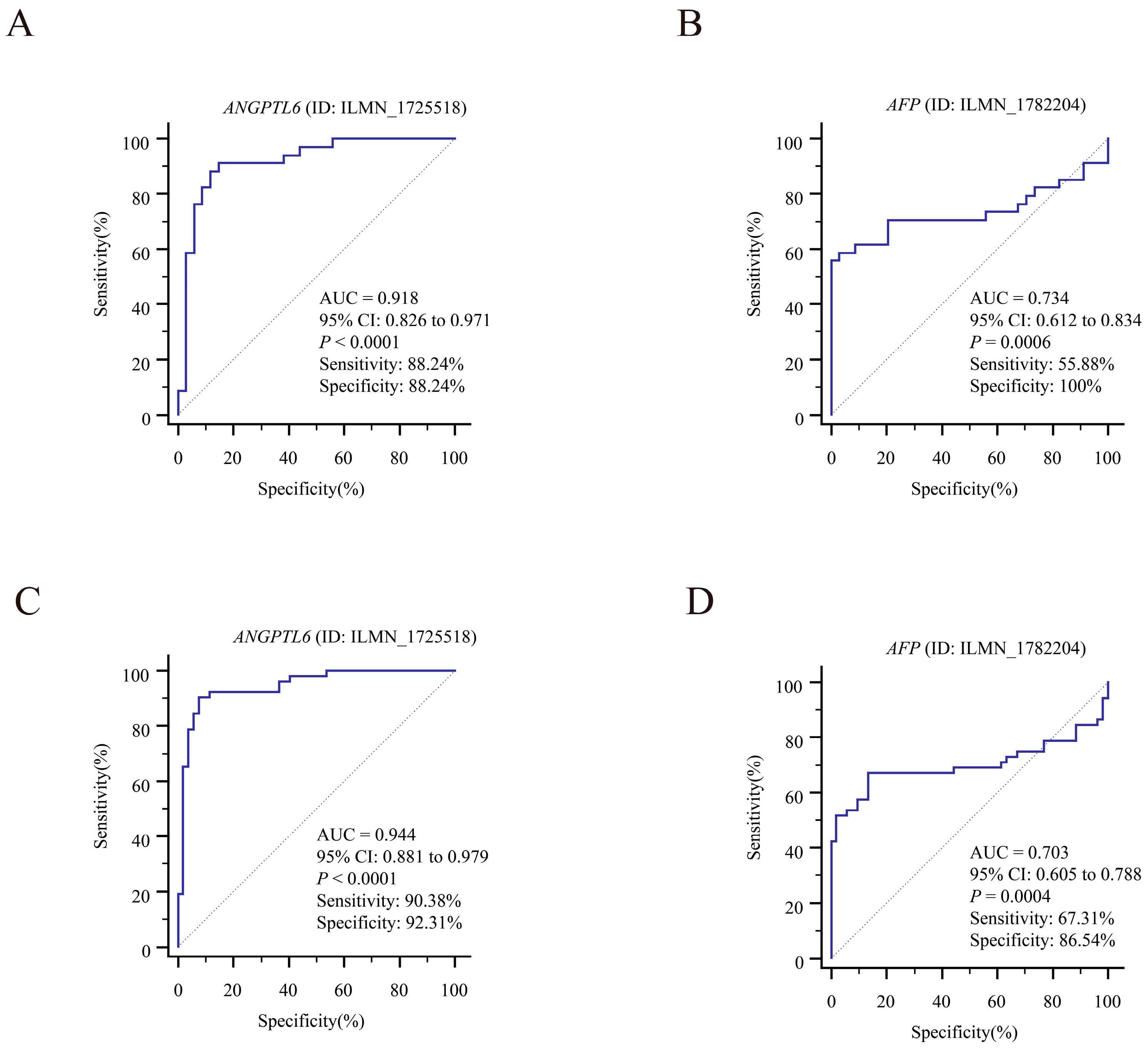
3.5. Exploring the Clinical Value of the 10 DEGs
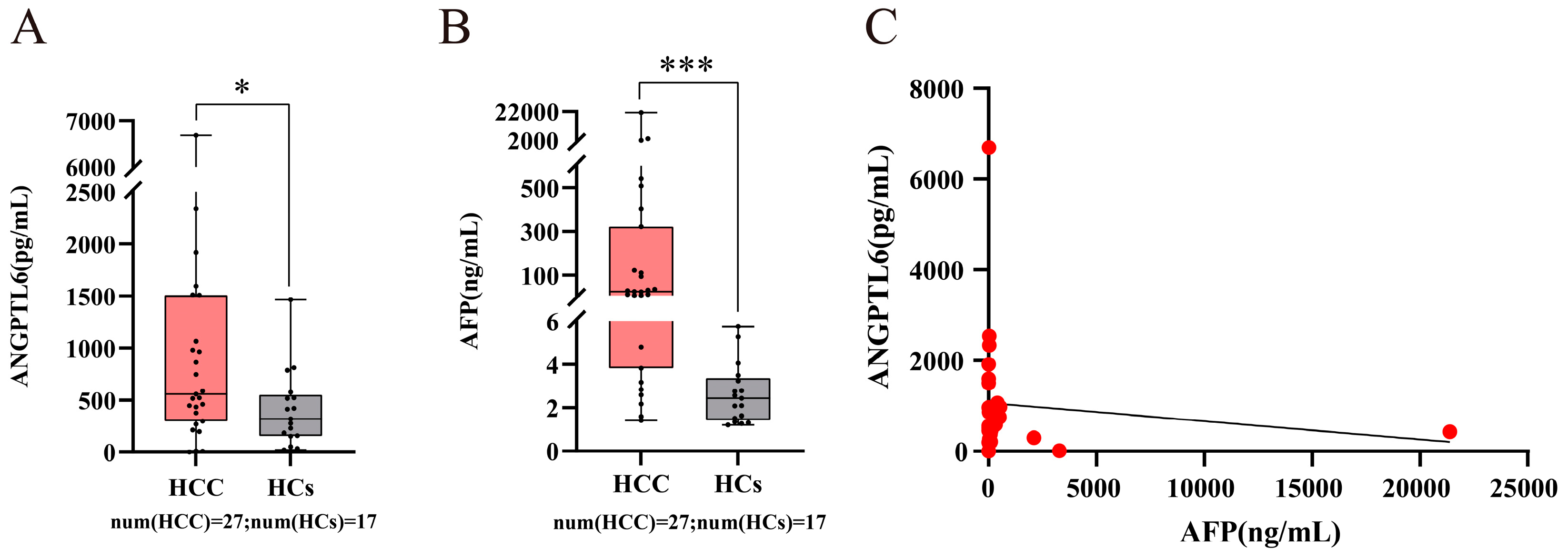
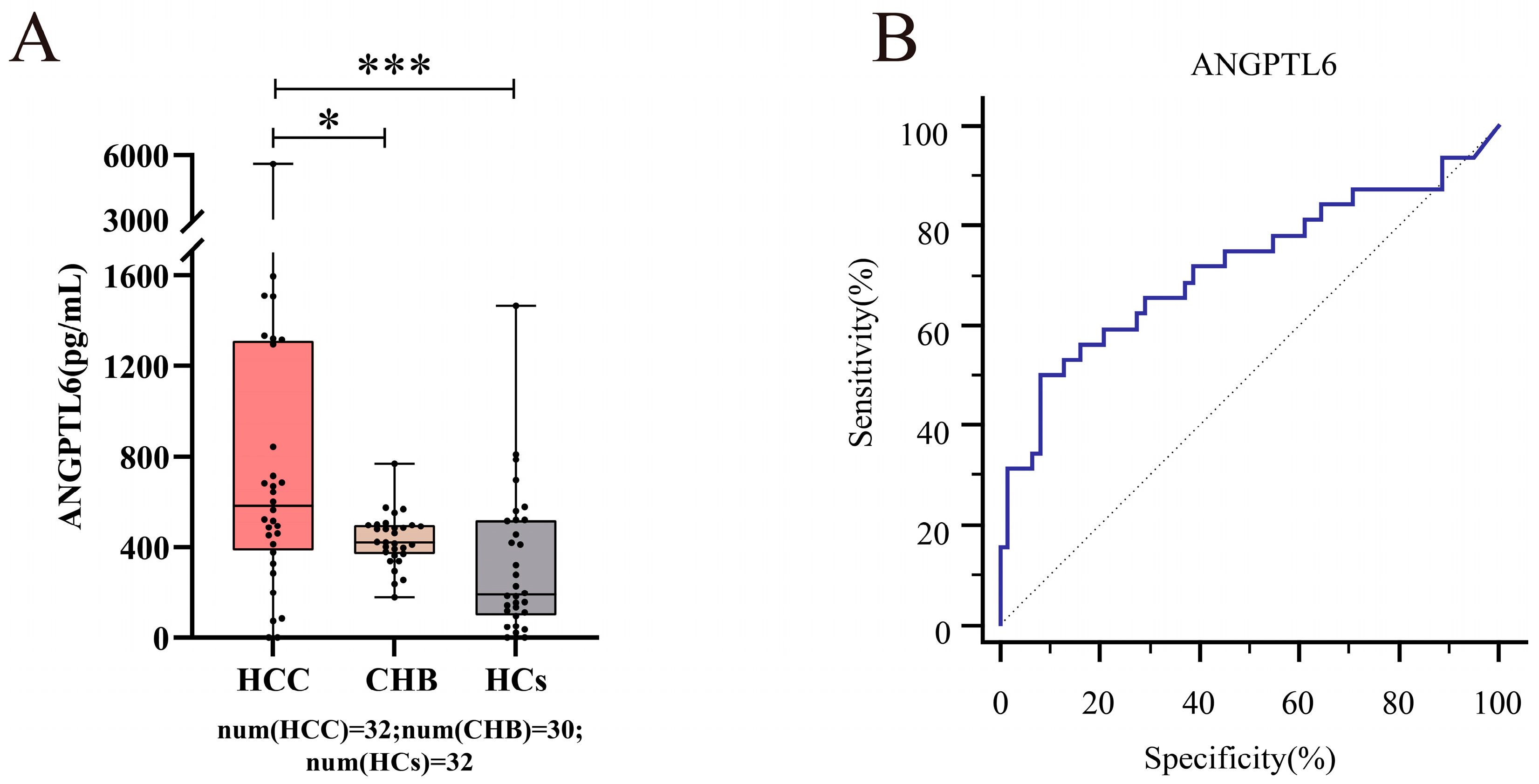
3.6. Serum ANGPTL6 Levels for the Diagnosis of Early Primary HCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1H-NMR | 1H-nuclear Magnetic Resonance |
| ADH4 | alcohol dehydrogenase 4 (class II), pi polypeptide |
| AFP | alpha-fetoprotein |
| ANGPTL6 | angiopoietin-like 6 |
| AUC | area under the ROC curve |
| BCLC | Barcelona Clinic Liver Cancer |
| CCLE | Cancer Cell Line Encyclopedia |
| CLEC4G | C-type lectin domain family 4 member G |
| CLEC4M | C-type lectin domain family 4 member M |
| CYP2C9 | cytochrome P450 family 2 subfamily C member 9 |
| DAVID | Database for Annotation, Visualization, and Integrated Discovery |
| DEGs | differentially expressed genes |
| DEPs | differentially expressed proteins |
| DFS | disease-free survival |
| FC | fold change |
| FDR | false discovery rate |
| GEO | Gene Expression Ominibus |
| GEPIA | Gene Expression Profiling Interactive Analysis |
| GYS2 | glycogen synthase 2 |
| HBP21 | heat shock binding protein 21 |
| HBV | hepatitis B virus |
| HCC | hepatocellular carcinoma |
| HLM | hemostasis and lipid metabolism |
| HM450 | HumanMethylation450 BeadChip |
| HPA | Human Protein Atlas |
| HR | hazard ratio |
| IQR | interquartile range |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LIHC | liver hepatocellular carcinoma |
| MBL2 | mannose binding lectin 2 |
| Nano-LC-MS/MS | Nanoscale Liquid Chromatography–Tandem Mass Spectrometry |
| OS | overall survival |
| PHYHD1 | phytanoyl-CoA dioxygenase domain containing 1 |
| ROC | receiver operating characteristic |
| RSEM | RNA-Seq by Expectation-Maximization |
| SLC27A5 | solute carrier family 27 member 5 |
| TAM | tumor-associated macrophages |
| TCGA | The Cancer Genome Atlas |
| TIMER | Tumor Immune Estimation Resource |
| TPM | transcripts per million |
| TTC36 | tetratricopeptide repeat domain 36 |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.I.; Jiao, J.; Kwan, S.Y.; Veillon, L.; Warmoes, M.O.; Tan, L.; Odewole, M.; Rich, N.E.; Wei, P.; Lorenzi, P.L.; et al. Lipidomic Profiles of Plasma Exosomes Identify Candidate Biomarkers for Early Detection of Hepatocellular Carcinoma in Patients with Cirrhosis. Cancer Prev. Res. (Phila) 2021, 14, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, P.; He, Y.; Chen, Z.; Chen, L.; Luo, Y.; Qi, L.; Liu, Y.; Wu, Q.; Cui, Y.; et al. HCC-derived exosomes elicit HCC progression and recurrence by epithelial-mesenchymal transition through MAPK/ERK signalling pathway. Cell. Death Dis. 2018, 9, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Lu, Z.; Zhao, X. Tumorigenesis, diagnosis, and therapeutic potential of exosomes in liver cancer. J. Hematol. Oncol. 2019, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Xu, H.F.; Liu, M.Y.; Xu, Y.J.; He, J.C.; Zhou, Y.; Cang, S.D. Mechanism of exosomal microRNA-224 in development of hepatocellular carcinoma and its diagnostic and prognostic value. World J. Gastroenterol. 2019, 25, 1890–1898. [Google Scholar] [CrossRef]
- Silva, M.A.; Hegab, B.; Hyde, C.; Guo, B.; Buckels, J.A.; Mirza, D.F. Needle track seeding following biopsy of liver lesions in the diagnosis of hepatocellular cancer: A systematic review and meta-analysis. Gut 2008, 57, 1592–1596. [Google Scholar] [CrossRef]
- Ignatiadis, M.; Sledge, G.W.; Jeffrey, S.S. Liquid biopsy enters the clinic—Implementation issues and future challenges. Nat. Rev. Clin. Oncol. 2021, 18, 297–312. [Google Scholar] [CrossRef]
- Ramon, Y.C.S.; Sese, M.; Capdevila, C.; Aasen, T.; De Mattos-Arruda, L.; Diaz-Cano, S.J.; Hernandez-Losa, J.; Castellvi, J. Clinical implications of intratumor heterogeneity: Challenges and opportunities. J. Mol. Med. (Berl.) 2020, 98, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Lu, Q.; Liu, X.; Cong, H.; Zhao, L.; Wang, H.; Lin, D. Application of 1H NMR-based metabonomics in the study of metabolic profiling of human hepatocellular carcinoma and liver cirrhosis. Cancer Sci. 2009, 100, 782–785. [Google Scholar] [CrossRef]
- Luo, P.; Wu, S.; Yu, Y.; Ming, X.; Li, S.; Zuo, X.; Tu, J. Current Status and Perspective Biomarkers in AFP Negative HCC: Towards Screening for and Diagnosing Hepatocellular Carcinoma at an Earlier Stage. Pathol. Oncol. Res. 2020, 26, 599–603. [Google Scholar] [CrossRef]
- Yang, J.D.; Heimbach, J.K. New advances in the diagnosis and management of hepatocellular carcinoma. BMJ (Clin. Res. Ed.) 2020, 371, m3544. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.R.; Sirlin, C.B.; Zaiem, F.; Almasri, J.; Prokop, L.J.; Heimbach, J.K.; Murad, M.H.; Mohammed, K. Imaging for the diagnosis of hepatocellular carcinoma: A systematic review and meta-analysis. Hepatology (Baltim. Md.) 2018, 67, 401–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Tommaso, L.; Spadaccini, M.; Donadon, M.; Personeni, N.; Elamin, A.; Aghemo, A.; Lleo, A. Role of liver biopsy in hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 6041–6052. [Google Scholar] [CrossRef] [PubMed]
- Pinero, F.; Dirchwolf, M.; Pessoa, M.G. Biomarkers in Hepatocellular Carcinoma: Diagnosis, Prognosis and Treatment Response Assessment. Cells 2020, 9, 1370. [Google Scholar] [CrossRef]
- Cai, F.F.; Song, Y.N.; Lu, Y.Y.; Zhang, Y.; Hu, Y.Y.; Su, S.B. Analysis of plasma metabolic profile, characteristics and enzymes in the progression from chronic hepatitis B to hepatocellular carcinoma. Aging (Albany NY) 2020, 12, 14949–14965. [Google Scholar] [CrossRef]
- Rumgay, H.; Arnold, M.; Ferlay, J.; Lesi, O.; Cabasag, C.J.; Vignat, J.; Laversanne, M.; McGlynn, K.A.; Soerjomataram, I. Global burden of primary liver cancer in 2020 and predictions to 2040. J. Hepatol. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Shi, J.F.; Cao, M.; Wang, Y.; Bai, F.Z.; Lei, L.; Peng, J.; Feletto, E.; Canfell, K.; Qu, C.; Chen, W. Is it possible to halve the incidence of liver cancer in China by 2050? Int. J. Cancer 2021, 148, 1051–1065. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Broadfield, L.A.; Pane, A.A.; Talebi, A.; Swinnen, J.V.; Fendt, S.M. Lipid metabolism in cancer: New perspectives and emerging mechanisms. Dev. Cell 2021, 56, 1363–1393. [Google Scholar] [CrossRef]
- Merino Salvador, M.; Gomez de Cedron, M.; Moreno Rubio, J.; Falagan Martinez, S.; Sanchez Martinez, R.; Casado, E.; Ramirez de Molina, A.; Sereno, M. Lipid metabolism and lung cancer. Crit. Rev. Oncol. Hematol. 2017, 112, 31–40. [Google Scholar] [CrossRef]
- Nazih, H.; Bard, J.M. Cholesterol, Oxysterols and LXRs in Breast Cancer Pathophysiology. Int. J. Mol. Sci. 2020, 21, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in lipids composition and metabolism in colorectal cancer: A review. Lipids. Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Zheng, Q.; Zhu, W.; Chen, X.; Lu, H.; Chen, D.; Zhang, H.; Shao, M.; Zhou, L.; Zheng, S. Alterations in glycolytic/cholesterogenic gene expression in hepatocellular carcinoma. Aging 2020, 12, 10300–10316. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhai, L.; Yi, T.; Gao, H.; Fan, F.; Li, Y.; Wang, Y.; Li, N.; Xing, X.; Su, N.; et al. Hepatitis B virus X induces inflammation and cancer in mice liver through dysregulation of cytoskeletal remodeling and lipid metabolism. Oncotarget 2016, 7, 70559–70574. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.X.; Huang, C.J.; Yang, Z.G. Impact of hepatitis B virus infection on hepatic metabolic signaling pathway. World J. Gastroenterol. 2016, 22, 8161–8167. [Google Scholar] [CrossRef]
- Qiao, L.; Luo, G.G. Human apolipoprotein E promotes hepatitis B virus infection and production. PLoS Pathog. 2019, 15, e1007874. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, N.M.; Wing, P.A.C.; Diniz, M.O.; Pallett, L.J.; Swadling, L.; Harris, J.M.; Burton, A.R.; Jeffery-Smith, A.; Zakeri, N.; Amin, O.E.; et al. Targeting human Acyl-CoA:cholesterol acyltransferase as a dual viral and T cell metabolic checkpoint. Nat. Commun. 2021, 12, 2814. [Google Scholar] [CrossRef]
- Buechler, C.; Aslanidis, C. Role of lipids in pathophysiology, diagnosis and therapy of hepatocellular carcinoma. Biochim. Et Biophys. Acta Mol. Cell. Biol. Lipids. 2020, 1865, 158658. [Google Scholar] [CrossRef]
- Wang, X.; Gong, Y.; Deng, T.; Zhang, L.; Liao, X.; Han, C.; Yang, C.; Huang, J.; Wang, Q.; Song, X.; et al. Diagnostic and prognostic significance of mRNA expressions of apolipoprotein A and C family genes in hepatitis B virus-related hepatocellular carcinoma. J. Cell. Biochem. 2019, 120, 18246–18265. [Google Scholar] [CrossRef]
- Zuo, D.; An, H.; Li, J.; Xiao, J.; Ren, L. The Application Value of Lipoprotein Particle Numbers in the Diagnosis of HBV-Related Hepatocellular Carcinoma with BCLC Stage 0-A. J. Pers Med. 2021, 11, 1143. [Google Scholar] [CrossRef]
- Deprince, A.; Haas, J.T.; Staels, B. Dysregulated lipid metabolism links NAFLD to cardiovascular disease. Mol. Metab. 2020, 42, 101092. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, L.; Xu, S.; Shen, A.Z. ATP-citrate lyase (ACLY) in lipid metabolism and atherosclerosis: An updated review. Prog. Lipid Res. 2020, 77, 101006. [Google Scholar] [CrossRef] [PubMed]
- Soppert, J.; Lehrke, M.; Marx, N.; Jankowski, J.; Noels, H. Lipoproteins and lipids in cardiovascular disease: From mechanistic insights to therapeutic targeting. Adv. Drug. Deliv. Rev. 2020, 159, 4–33. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.J.; Sorci-Thomas, M.G. Linking Lipid-Related Transcription and Cardiovascular Disease Through GWAS. Circ. Res. 2020, 127, 1362–1364. [Google Scholar] [CrossRef] [PubMed]
- Abumrad, N.A.; Cabodevilla, A.G.; Samovski, D.; Pietka, T.; Basu, D.; Goldberg, I.J. Endothelial Cell Receptors in Tissue Lipid Uptake and Metabolism. Circ. Res. 2021, 128, 433–450. [Google Scholar] [CrossRef]
- Kim, A.S.; Conte, M.S. Specialized pro-resolving lipid mediators in cardiovascular disease, diagnosis, and therapy. Adv. Drug. Deliv. Rev. 2020, 159, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, L.; Faucher, G.; Tchernof, A.; Deshaies, Y.; Lebel, S.; Hould, F.S.; Marceau, P.; Vohl, M.C. Comprehensive genetic analysis of the dipeptidyl peptidase-4 gene and cardiovascular disease risk factors in obese individuals. Acta Diabetol. 2009, 46, 13–21. [Google Scholar] [CrossRef]
- Hernandez-Gea, V.; Camprecios, G.; Betancourt, F.; Perez-Campuzano, V.; Seijo, S.; Diaz, A.; Gallego-Duran, R.; Olivas, P.; Orts, L.; Magaz, M.; et al. Co-expression gene network analysis reveals novel regulatory pathways involved in porto-sinusoidal vascular disease. J. Hepatol. 2021, 75, 924–934. [Google Scholar] [CrossRef]
- Harshfield, E.L.; Fauman, E.B.; Stacey, D.; Paul, D.S.; Ziemek, D.; Ong, R.M.Y.; Danesh, J.; Butterworth, A.S.; Rasheed, A.; Sattar, T.; et al. Genome-wide analysis of blood lipid metabolites in over 5000 South Asians reveals biological insights at cardiometabolic disease loci. BMC Med. 2021, 19, 232. [Google Scholar] [CrossRef]
- De Jesus, D.F.; Orime, K.; Kaminska, D.; Kimura, T.; Basile, G.; Wang, C.H.; Haertle, L.; Riemens, R.; Brown, N.K.; Hu, J.; et al. Parental metabolic syndrome epigenetically reprograms offspring hepatic lipid metabolism in mice. J. Clin. Investig. 2020, 130, 2391–2407. [Google Scholar] [CrossRef]
- Chiyonobu, N.; Shimada, S.; Akiyama, Y.; Mogushi, K.; Itoh, M.; Akahoshi, K.; Matsumura, S.; Ogawa, K.; Ono, H.; Mitsunori, Y.; et al. Fatty Acid Binding Protein 4 (FABP4) Overexpression in Intratumoral Hepatic Stellate Cells within Hepatocellular Carcinoma with Metabolic Risk Factors. Am. J. Pathol. 2018, 188, 1213–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva, A.; Portela, A.; Sayols, S.; Battiston, C.; Hoshida, Y.; Mendez-Gonzalez, J.; Imbeaud, S.; Letouze, E.; Hernandez-Gea, V.; Cornella, H.; et al. DNA methylation-based prognosis and epidrivers in hepatocellular carcinoma. Hepatology 2015, 61, 1945–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinchuk, O.V.; Yenamandra, S.P.; Iyer, R.; Singh, M.; Lee, H.K.; Lim, K.H.; Chow, P.K.; Kuznetsov, V.A. Tumor-adjacent tissue co-expression profile analysis reveals pro-oncogenic ribosomal gene signature for prognosis of resectable hepatocellular carcinoma. Mol. Oncol. 2018, 12, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, G.; Engle, R.E.; Tice, A.; Melis, M.; Montenegro, S.; Rodriguez-Canales, J.; Hanson, J.; Emmert-Buck, M.R.; Bock, K.W.; Moore, I.N.; et al. Molecular Signature and Mechanisms of Hepatitis D Virus-Associated Hepatocellular Carcinoma. Mol. Cancer Res. 2018, 16, 1406–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids. Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Zuo, D.; Chen, Y.; Zhang, X.; Wang, Z.; Jiang, W.; Tang, F.; Cheng, R.; Sun, Y.; Sun, L.; Ren, L.; et al. Identification of hub genes and their novel diagnostic and prognostic significance in pancreatic adenocarcinoma. Cancer Biol. Med. 2021, 19, 1029–1046. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids. Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Dastsooz, H.; Cereda, M.; Donna, D.; Oliviero, S. A Comprehensive Bioinformatics Analysis of UBE2C in Cancers. Int. J. Mol. Sci. 2019, 20, 2228. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, N.; Zhang, X.; Xiao, J.; Li, J.; Lv, D.; Zhou, W.; Li, Y.; Xu, J.; Li, X. SurvivalMeth: A web server to investigate the effect of DNA methylation-related functional elements on prognosis. Brief. Bioinform. 2021, 22, bbaa162. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Severson, E.; Pignon, J.C.; Zhao, H.; Li, T.; Novak, J.; Jiang, P.; Shen, H.; Aster, J.C.; Rodig, S.; et al. Comprehensive analyses of tumor immunity: Implications for cancer immunotherapy. Genome Biol. 2016, 17, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Association for the Study of the Liver. Electronic address, e.e.e.; European Association for the Study of the, L. EASL Clinical Practice Guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forner, A.; Reig, M.E.; de Lope, C.R.; Bruix, J. Current strategy for staging and treatment: The BCLC update and future prospects. Semin.Liver. Dis. 2010, 30, 61–74. [Google Scholar] [CrossRef]
- Ahn, J.C.; Teng, P.C.; Chen, P.J.; Posadas, E.; Tseng, H.R.; Lu, S.C.; Yang, J.D. Detection of Circulating Tumor Cells and Their Implications as a Biomarker for Diagnosis, Prognostication, and Therapeutic Monitoring in Hepatocellular Carcinoma. Hepatology 2021, 73, 422–436. [Google Scholar] [CrossRef]
- De Carvalho, D.D.; Sharma, S.; You, J.S.; Su, S.F.; Taberlay, P.C.; Kelly, T.K.; Yang, X.; Liang, G.; Jones, P.A. DNA methylation screening identifies driver epigenetic events of cancer cell survival. Cancer Cell. 2012, 21, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Hu, B.; Lin, J.Z.; Yang, X.B.; Sang, X.T. Aberrant lipid metabolism in hepatocellular carcinoma cells as well as immune microenvironment: A review. Cell Prolif. 2020, 53, e12772. [Google Scholar] [CrossRef] [Green Version]
- Loglio, A.; Iavarone, M.; Vigano, M.; Orenti, A.; Facchetti, F.; Cortinovis, I.; Lunghi, G.; Ceriotti, F.; Occhipinti, V.; Rumi, M.; et al. Minimal increases of serum alpha-foetoprotein herald HCC detection in Caucasian HBV cirrhotic patients under long-term oral therapy. Liver. Int. 2019, 39, 1964–1974. [Google Scholar] [CrossRef]
- Li, H.; Chen, Z.; Zhang, Y.; Yuan, P.; Liu, J.; Ding, L.; Ye, Q. MiR-4310 regulates hepatocellular carcinoma growth and metastasis through lipid synthesis. Cancer Lett. 2021, 519, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Soukupova, J.; Malfettone, A.; Bertran, E.; Hernandez-Alvarez, M.I.; Penuelas-Haro, I.; Dituri, F.; Giannelli, G.; Zorzano, A.; Fabregat, I. Epithelial-Mesenchymal Transition (EMT) Induced by TGF-beta in Hepatocellular Carcinoma Cells Reprograms Lipid Metabolism. Int. J. Mol. Sci. 2021, 22, 5543. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tan, Y.; Jiang, T.; Wang, X.; Li, Q.; Li, Y.; Dong, L.; Liu, X.; Xu, G. Metabolic Reprogramming and Its Relationship to Survival in Hepatocellular Carcinoma. Cells 2022, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, C.; Yu, Q.; Zheng, X.; Yin, C.; Yan, X.; Liu, G.; Song, Z. Development of a novel lipid metabolism-based risk score model in hepatocellular carcinoma patients. BMC Gastroenterol. 2021, 21, 68. [Google Scholar] [CrossRef]
- Berndt, N.; Eckstein, J.; Heucke, N.; Gajowski, R.; Stockmann, M.; Meierhofer, D.; Holzhutter, H.G. Characterization of Lipid and Lipid Droplet Metabolism in Human HCC. Cells 2019, 8, 512. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Cai, L.; Huang, W.; Weng, Q.; Lin, X.; You, M.; Liao, S. Prognostic value of fatty acid metabolism-related genes in patients with hepatocellular carcinoma. Aging (Albany NY) 2021, 13, 17847–17863. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Z.; Xia, S.; Yao, L.; Li, L.; Gan, Z.; Tang, H.; Guo, Q.; Yan, X.; Sun, Z. GDI2 is a novel diagnostic and prognostic biomarker in hepatocellular carcinoma. Aging (Albany NY) 2021, 13, 25304–25324. [Google Scholar] [CrossRef]
- Chai, H.; Xia, L.; Zhang, L.; Yang, J.; Zhang, Z.; Qian, X.; Yang, Y.; Pan, W. An Adaptive Transfer-Learning-Based Deep Cox Neural Network for Hepatocellular Carcinoma Prognosis Prediction. Front. Oncol. 2021, 11, 692774. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, Q.; Xue, B.; Wang, X.; Tian, R.; Deng, R.; Chen, S.; Wang, X.; Wang, L.; Wang, C.; et al. Heat Shock-Binding Protein 21 Regulates the Innate Immune Response to Viral Infection. J. Virol. 2022, 96, e0000122. [Google Scholar] [CrossRef]
- Jiang, L.; Kwong, D.L.; Li, Y.; Liu, M.; Yuan, Y.F.; Li, Y.; Fu, L.; Guan, X.Y. HBP21, a chaperone of heat shock protein 70, functions as a tumor suppressor in hepatocellular carcinoma. Carcinogenesis 2015, 36, 1111–1120. [Google Scholar] [CrossRef]
- Miyashita, A.; Hatsuta, H.; Kikuchi, M.; Nakaya, A.; Saito, Y.; Tsukie, T.; Hara, N.; Ogishima, S.; Kitamura, N.; Akazawa, K.; et al. Genes associated with the progression of neurofibrillary tangles in Alzheimer’s disease. Transl. Psychiatry 2014, 4, e396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Huang, Q.; Ding, Z.; Liu, T.; Xue, C.; Sang, X.; Gu, J. Genome-wide DNA methylation analysis identifies candidate epigenetic markers and drivers of hepatocellular carcinoma. Brief. Bioinform. 2018, 19, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef]
- Zhang, F.; Xue, M.; Jiang, X.; Yu, H.; Qiu, Y.; Yu, J.; Yang, F.; Bao, Z. Identifying SLC27A5 as a potential prognostic marker of hepatocellular carcinoma by weighted gene co-expression network analysis and in vitro assays. Cancer Cell Int. 2021, 21, 174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, Z.; Sun, S.; Wang, K.; Qian, J.; Cui, Z.; Tao, T.; Zhou, J. Integrated Proteomics and Bioinformatics to Identify Potential Prognostic Biomarkers in Hepatocellular Carcinoma. Cancer Manag. Res. 2021, 13, 2307–2317. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Chen, Z.; Liu, G.; Dai, Y.; Xu, X.; Ma, D.; Liu, L. Identification of a Potential PPAR-Related Multigene Signature Predicting Prognosis of Patients with Hepatocellular Carcinoma. PPAR Res. 2021, 2021, 6642939. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Fang, X.; Feng, Y.; Cui, X.; Li, F.; Luo, W.; Ma, X.; Liang, J.; Feng, J. Identification of key genes associated with the progression of liver fibrosis to hepatocellular carcinoma based on iTRAQ proteomics and GEO database. Ann. Hepatol. 2022, 27, 100681. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Yang, X.B.; Sang, X.T. Construction of a lipid metabolism-related and immune-associated prognostic signature for hepatocellular carcinoma. Cancer Med. 2020, 9, 7646–7662. [Google Scholar] [CrossRef]
- Su, P.; Wang, Q.; Bi, E.; Ma, X.; Liu, L.; Yang, M.; Qian, J.; Yi, Q. Enhanced Lipid Accumulation and Metabolism Are Required for the Differentiation and Activation of Tumor-Associated Macrophages. Cancer Res. 2020, 80, 1438–1450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, H.; Mao, C.; Sun, M.; Dominah, G.; Chen, L.; Zhuang, Z. Fatty acid oxidation contributes to IL-1beta secretion in M2 macrophages and promotes macrophage-mediated tumor cell migration. Mol. Immunol. 2018, 94, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Oniciu, D.C.; Hashiguchi, T.; Shibazaki, Y.; Bisgaier, C.L. Gemcabene downregulates inflammatory, lipid-altering and cell-signaling genes in the STAM model of NASH. PLoS ONE 2018, 13, e0194568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.R.; Zhang, M.Y.; Rao, H.L.; Pu, H.Y.; Zhang, H.Z.; Wang, H.Y. Identification of ADH4 as a novel and potential prognostic marker in hepatocellular carcinoma. Med. Oncol. 2012, 29, 2737–2743. [Google Scholar] [CrossRef]
- Zhou, C.; Wei, W.; Ma, J.; Yang, Y.; Liang, L.; Zhang, Y.; Wang, Z.; Chen, X.; Huang, L.; Wang, W.; et al. Cancer-secreted exosomal miR-1468-5p promotes tumor immune escape via the immunosuppressive reprogramming of lymphatic vessels. Mol. Ther. J. Am. Soc. Gene Ther. 2021, 29, 1512–1528. [Google Scholar] [CrossRef]
- Liu, X.; Li, T.; Kong, D.; You, H.; Kong, F.; Tang, R. Prognostic implications of alcohol dehydrogenases in hepatocellular carcinoma. BMC Cancer 2020, 20, 1204. [Google Scholar] [CrossRef]
- Li, Z.; Lei, G.; Meng, X.; Yang, Z. Identification of a new five-gene risk score for risk stratification and prognosis prediction in HCC. Nucleosides Nucleotides Nucleic Acids 2022, 41, 736–754. [Google Scholar] [CrossRef]
- Xu, C.Y.; Dong, J.F.; Chen, Z.Q.; Ding, G.S.; Fu, Z.R. MiR-942-3p Promotes the Proliferation and Invasion of Hepatocellular Carcinoma Cells by Targeting MBL2. Cancer Control. 2019, 26, 1073274819846593. [Google Scholar] [CrossRef]
- Su, C.; Lin, Y.; Cai, L.; Mao, Q.; Niu, J. Association between mannose-binding lectin variants, haplotypes and risk of hepatocellular carcinoma: A case-control study. Sci. Rep. 2016, 6, 32147. [Google Scholar] [CrossRef] [Green Version]
- Awan, F.M.; Naz, A.; Obaid, A.; Ali, A.; Ahmad, J.; Anjum, S.; Janjua, H.A. Identification of Circulating Biomarker Candidates for Hepatocellular Carcinoma (HCC): An Integrated Prioritization Approach. PLoS ONE 2015, 10, e0138913. [Google Scholar] [CrossRef]
- Li, J.; Cheng, Z.J.; Liu, Y.; Yan, Z.L.; Wang, K.; Wu, D.; Wan, X.Y.; Xia, Y.; Lau, W.Y.; Wu, M.C.; et al. Serum thioredoxin is a diagnostic marker for hepatocellular carcinoma. Oncotarget 2015, 6, 9551–9563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, M. The Roles of ANGPTL Families in Cancer Progression. J. UOEH 2019, 41, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Tang, C.; Peng, K.; Cheng, X.; Wei, Y.; Liu, T. ANGPTL6-mediated angiogenesis promotes alpha fetoprotein-producing gastric cancer progression. Pathol. Res. Pract. 2019, 215, 152454. [Google Scholar] [CrossRef] [PubMed]
- Marchio, S.; Soster, M.; Cardaci, S.; Muratore, A.; Bartolini, A.; Barone, V.; Ribero, D.; Monti, M.; Bovino, P.; Sun, J.; et al. A complex of alpha6 integrin and E-cadherin drives liver metastasis of colorectal cancer cells through hepatic angiopoietin-like 6. EMBO Mol. Med. 2012, 4, 1156–1175. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Gao, F.; Wang, G.; Fang, Y.; Guo, Y.; Zhou, J.; Gu, Y.; Zhang, C.; Gao, N.; Wen, Q.; et al. Use of proteomics to identify mechanisms of hepatocellular carcinoma with the CYP2D6*10 polymorphism and identification of ANGPTL6 as a new diagnostic and prognostic biomarker. J. Transl. Med. 2021, 19, 359. [Google Scholar] [CrossRef]
- Kang, S.G.; Yi, H.S.; Choi, M.J.; Ryu, M.J.; Jung, S.; Chung, H.K.; Chang, J.Y.; Kim, Y.K.; Lee, S.E.; Kim, H.W.; et al. ANGPTL6 expression is coupled with mitochondrial OXPHOS function to regulate adipose FGF21. J. Endocrinol. 2017, 233, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Namkung, J.; Chang, J.S.; Kim, S.J.; Park, K.S.; Kong, I.D. Leptin regulates the expression of angiopoietin-like 6. Biochem. Biophys. Res. Commun. 2018, 502, 397–402. [Google Scholar] [CrossRef]
- Oike, Y.; Akao, M.; Yasunaga, K.; Yamauchi, T.; Morisada, T.; Ito, Y.; Urano, T.; Kimura, Y.; Kubota, Y.; Maekawa, H.; et al. Angiopoietin-related growth factor antagonizes obesity and insulin resistance. Nat. Med. 2005, 11, 400–408. [Google Scholar] [CrossRef] [PubMed]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, D.; Xiao, J.; An, H.; Chen, Y.; Li, J.; Yang, X.; Wang, X.; Ren, L. Screening for Lipid-Metabolism-Related Genes and Identifying the Diagnostic Potential of ANGPTL6 for HBV-Related Early-Stage Hepatocellular Carcinoma. Biomolecules 2022, 12, 1700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12111700
Zuo D, Xiao J, An H, Chen Y, Li J, Yang X, Wang X, Ren L. Screening for Lipid-Metabolism-Related Genes and Identifying the Diagnostic Potential of ANGPTL6 for HBV-Related Early-Stage Hepatocellular Carcinoma. Biomolecules. 2022; 12(11):1700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12111700
Chicago/Turabian StyleZuo, Duo, Jiawei Xiao, Haohua An, Yongzi Chen, Jianhua Li, Xiaohui Yang, Xia Wang, and Li Ren. 2022. "Screening for Lipid-Metabolism-Related Genes and Identifying the Diagnostic Potential of ANGPTL6 for HBV-Related Early-Stage Hepatocellular Carcinoma" Biomolecules 12, no. 11: 1700. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12111700