
Artemisia dracunculus L. Ethanolic Extract and an Isolated Component, DMC2, Ameliorate Inflammatory Signaling in Pancreatic β-Cells via Inhibition of p38 MAPK

 and
and 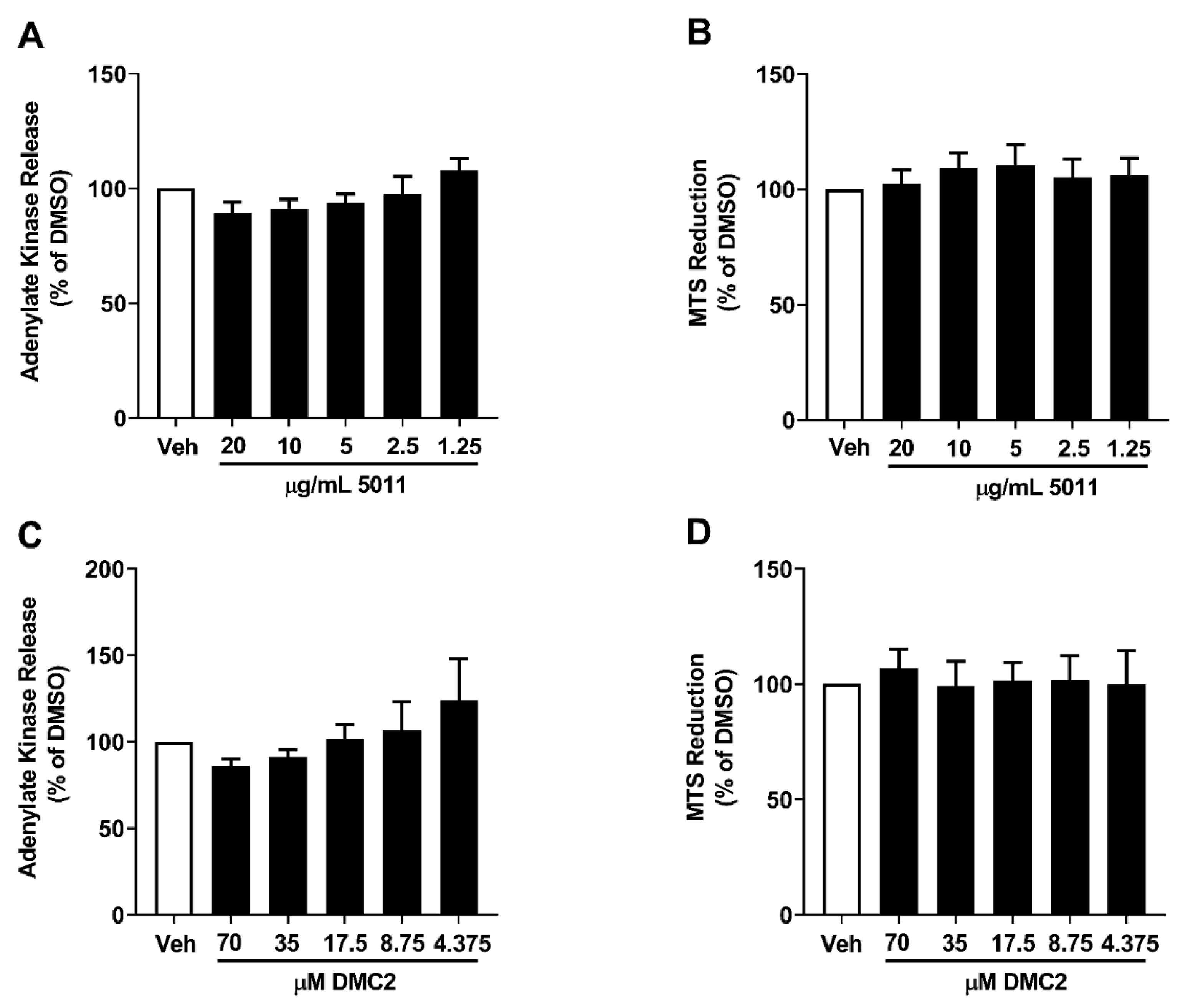{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Islet Isolation and Dispersal, and Reagents
2.2. Plant Material and Isolated Natural Product
2.3. Chemokine ELISAs
2.4. Protein Isolation and Immunoblotting
2.5. Isolation of RNA, cDNA Synthesis, and Real-Time RT-PCR
2.6. Plasmid and siRNA Transfection, and Luciferase Assays
2.7. Adenylate Kinase and MTS Assays
2.8. Glucose-Stimulated Insulin Secretion
2.9. Statistical Analysis
3. Results
3.1. PMI 5011 and DMC2 Do Not Promote β-Cell Toxicity
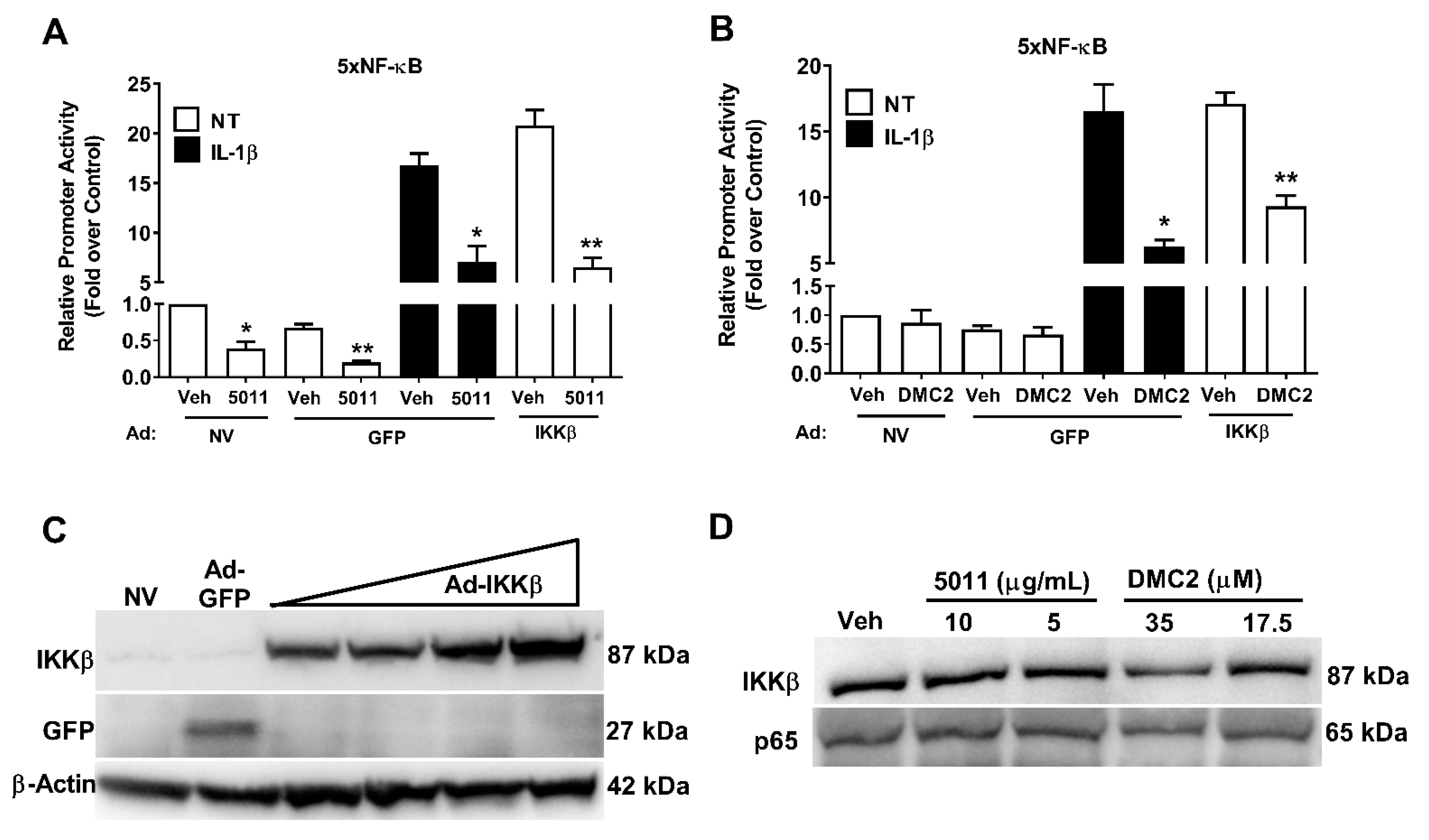
3.2. PMI 5011 and DMC2 Decrease IL-1β-Mediated NF-κB Activity
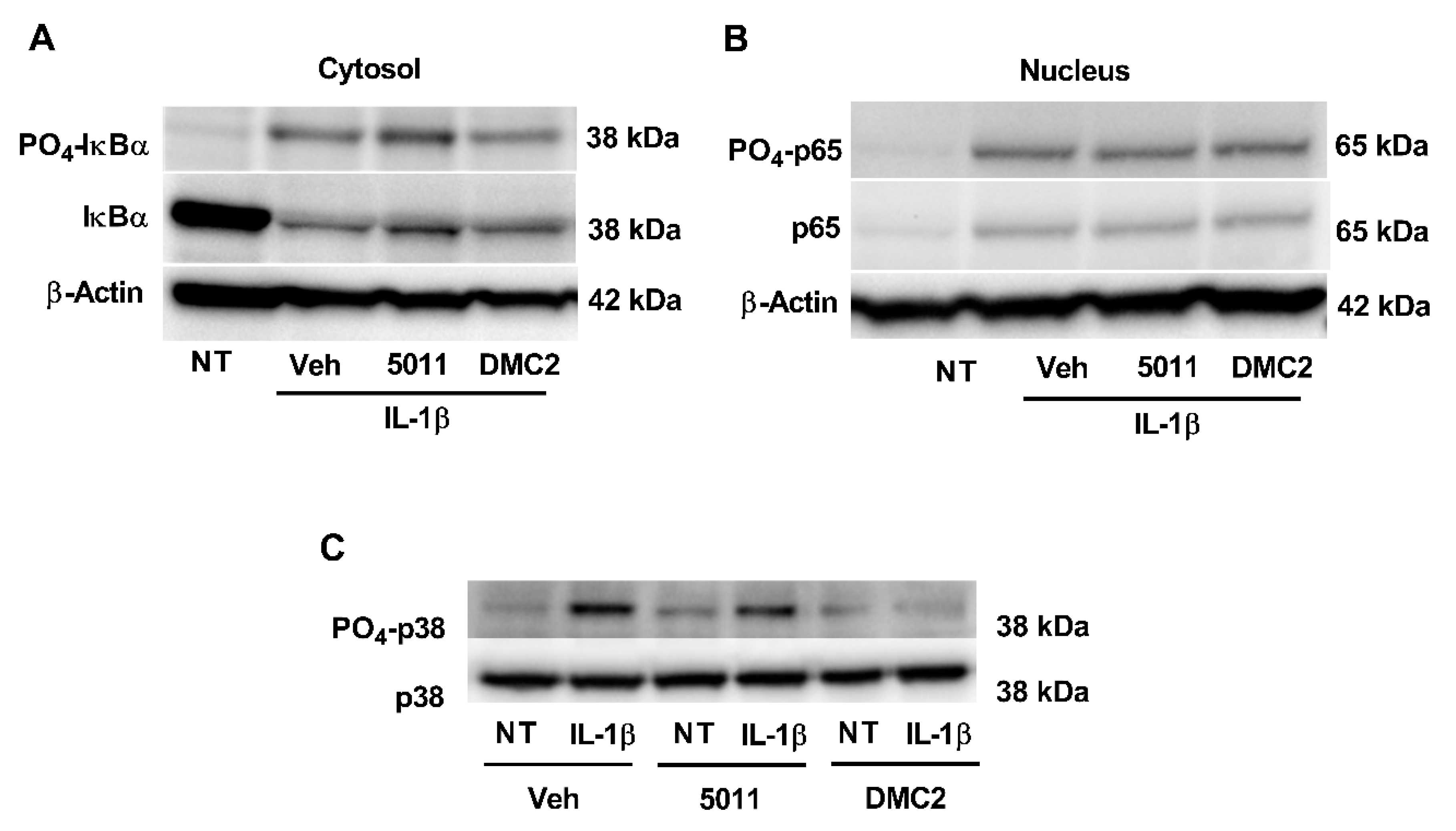
3.3. DMC2 Reduces p38 MAPK Phosphorylation but Not Phosphorylation of Either IκBα or p65
3.4. PMI 5011 and DMC2 Reduce Ccl2 Gene Expression and Protein Secretion
3.5. PMI 5011 and DMC2 Reduce Ccl20 Gene Expression and Protein Secretion
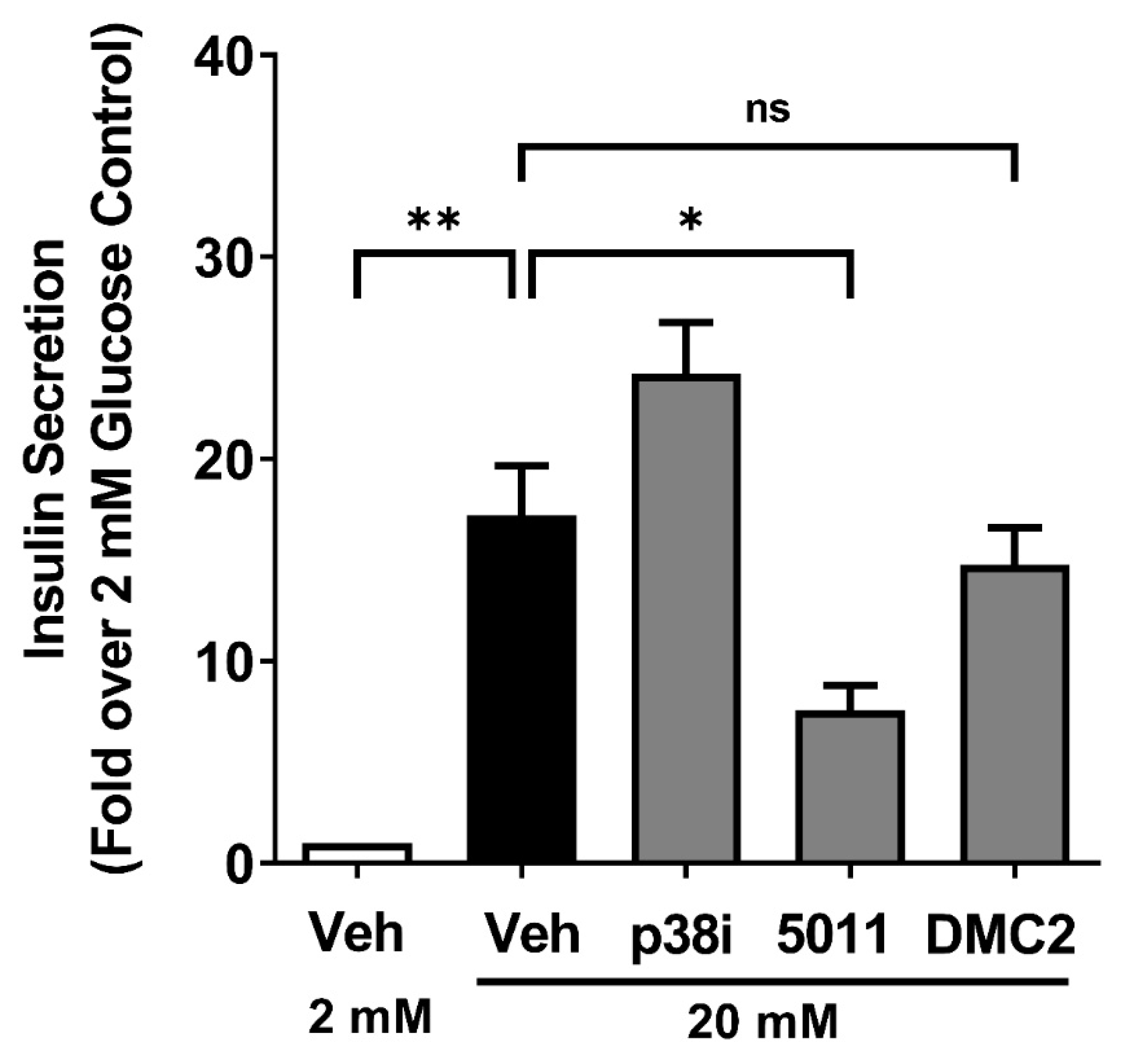
3.6. DMC2 Does Not Impair Glucose-Stimulated Insulin Secretion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kohlgruber, A.; Lynch, L. Adipose tissue inflammation in the pathogenesis of type 2 diabetes. Curr. Diabetes Rep. 2015, 15, 92. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, S.M.; Henschel, A.M.; Hessner, M.J. Innate inflammation in type 1 diabetes. Transl. Res. J. Lab. Clin. Med. 2016, 167, 214–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, S.J.; Collier, J.J. Transcriptional Regulation of Chemokine Genes: A Link to Pancreatic Islet Inflammation? Biomolecules 2015, 5, 1020–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.D.; Dixit, V.D. Immunological complications of obesity. Nat. Immunol. 2012, 13, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-kappaB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. IkappaB kinases: Key regulators of the NF-kappaB pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef]
- Mercurio, F.; Zhu, H.; Murray, B.W.; Shevchenko, A.; Bennett, B.L.; Li, J.; Young, D.B.; Barbosa, M.; Mann, M.; Manning, A.; et al. IKK-1 and IKK-2: Cytokine-activated IkappaB kinases essential for NF-kappaB activation. Science 1997, 278, 860–866. [Google Scholar] [CrossRef]
- Jobin, C.; Panja, A.; Hellerbrand, C.; Iimuro, Y.; Didonato, J.; Brenner, D.A.; Sartor, R.B. Inhibition of proinflammatory molecule production by adenovirus-mediated expression of a nuclear factor kappaB super-repressor in human intestinal epithelial cells. J. Immunol. 1998, 160, 410–418. [Google Scholar] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Shen, B.; Lin, A. Novel strategies for inhibition of the p38 MAPK pathway. Trends Pharmacol. Sci. 2007, 28, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C.; Chang, J.H.; Jin, J. Regulation of nuclear factor-kappaB in autoimmunity. Trends Immunol. 2013, 34, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Karin, M. Missing pieces in the NF-kappaB puzzle. Cell 2002, 109, S81–S96. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Collier, J.J.; Sparer, T.E.; Karlstad, M.D.; Burke, S.J. Pancreatic islet inflammation: An emerging role for chemokines. J. Mol. Endocrinol. 2017, 59, R33–R46. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Collier, J.J. Insulitis and Diabetes: A Perspective on Islet Inflammation. Immunome Res. 2014, 10, e002. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Dobrian, A.D.; Morris, M.A.; Nadler, J.L. Islet inflammation: A unifying target for diabetes treatment? Trends Endocrinol. Metab. 2013, 24, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Eizirik, D.L.; Colli, M.L.; Ortis, F. The role of inflammation in insulitis and beta-cell loss in type 1 diabetes. Nat. Rev. Endocrinol. 2009, 5, 219–226. [Google Scholar] [CrossRef]
- Donath, M.Y.; Boni-Schnetzler, M.; Ellingsgaard, H.; Ehses, J.A. Islet inflammation impairs the pancreatic beta-cell in type 2 diabetes. Physiology 2009, 24, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smoak, P.; Burke, S.J.; Collier, J.J. Botanical Interventions to Improve Glucose Control and Options for Diabetes Therapy. SN Compr. Clin. Med. 2021, 3, 2465–2491. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Karlstad, M.D.; Conley, C.P.; Reel, D.; Whelan, J.; Collier, J.J. Dietary polyherbal supplementation decreases CD3(+) cell infiltration into pancreatic islets and prevents hyperglycemia in nonobese diabetic mice. Nutr. Res. 2015, 35, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, H.; Swiatkowska, J.; Knut, E.; Klin, P.; Rzepiela, A.; Tomczyk, M.; Szopa, A. Artemisia dracunculus (Tarragon): A Review of Its Traditional Uses, Phytochemistry and Pharmacology. Front. Pharmacol. 2021, 12, 653993. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Simmler, C.; Kuhn, P.; Poulev, A.; Raskin, I.; Ribnicky, D.; Floyd, Z.E.; Pauli, G.F. The DESIGNER Approach Helps Decipher the Hypoglycemic Bioactive Principles of Artemisia dracunculus (Russian Tarragon). J. Nat. Prod. 2019, 82, 3321–3329. [Google Scholar] [CrossRef] [PubMed]
- Scherp, P.; Putluri, N.; LeBlanc, G.J.; Wang, Z.Q.; Zhang, X.H.; Yu, Y.; Ribnicky, D.; Cefalu, W.T.; Kheterpal, I. Proteomic analysis reveals cellular pathways regulating carbohydrate metabolism that are modulated in primary human skeletal muscle culture due to treatment with bioactives from Artemisia dracunculus L. J. Proteom. 2012, 75, 3199–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmeier, H.E.; Mulder, H.; Chen, G.; Henkel-Rieger, R.; Prentki, M.; Newgard, C.B. Isolation of INS-1-derived cell lines with robust ATP-sensitive K+ channel-dependent and -independent glucose-stimulated insulin secretion. Diabetes 2000, 49, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Collier, J.J. The gene encoding cyclooxygenase-2 is regulated by IL-1beta and prostaglandins in 832/13 rat insulinoma cells. Cell Immunol. 2011, 271, 379–384. [Google Scholar] [CrossRef]
- Burke, S.J.; Goff, M.R.; Updegraff, B.L.; Lu, D.; Brown, P.L.; Minkin, S.C., Jr.; Biggerstaff, J.P.; Zhao, L.; Karlstad, M.D.; Collier, J.J. Regulation of the CCL2 Gene in Pancreatic beta-Cells by IL-1beta and Glucocorticoids: Role of MKP-1. PLoS ONE 2012, 7, e46986. [Google Scholar] [CrossRef] [Green Version]
- Sekine, O.; Nishio, Y.; Egawa, K.; Nakamura, T.; Maegawa, H.; Kashiwagi, A. Insulin activates CCAAT/enhancer binding proteins and proinflammatory gene expression through the phosphatidylinositol 3-kinase pathway in vascular smooth muscle cells. J. Biol. Chem. 2002, 277, 36631–36639. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Karlstad, M.D.; Regal, K.M.; Sparer, T.E.; Lu, D.; Elks, C.M.; Grant, R.W.; Stephens, J.M.; Burk, D.H.; Collier, J.J. CCL20 is elevated during obesity and differentially regulated by NF-kappaB subunits in pancreatic beta-cells. Biochim. Biophys. Acta 2015, 1849, 637–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, S.J.; Updegraff, B.L.; Bellich, R.M.; Goff, M.R.; Lu, D.; Minkin, S.C., Jr.; Karlstad, M.D.; Collier, J.J. Regulation of iNOS Gene Transcription by IL-1beta and IFN-gamma Requires a Coactivator Exchange Mechanism. Mol. Endocrinol. 2013, 27, 1724–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk-Ballard, H.; Wang, Z.Q.; Acharya, P.; Zhang, X.H.; Yu, Y.; Kilroy, G.; Ribnicky, D.; Cefalu, W.T.; Floyd, Z.E. An extract of Artemisia dracunculus L. inhibits ubiquitin-proteasome activity and preserves skeletal muscle mass in a murine model of diabetes. PLoS ONE 2013, 8, e57112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haycock, J.W. Polyvinylpyrrolidone as a blocking agent in immunochemical studies. Anal. Biochem. 1993, 208, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Collier, J.J.; Fueger, P.T.; Hohmeier, H.E.; Newgard, C.B. Pro- and antiapoptotic proteins regulate apoptosis but do not protect against cytokine-mediated cytotoxicity in rat islets and beta-cell lines. Diabetes 2006, 55, 1398–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, J.J.; Burke, S.J.; Eisenhauer, M.E.; Lu, D.; Sapp, R.C.; Frydman, C.J.; Campagna, S.R. Pancreatic beta-Cell Death in Response to Pro-Inflammatory Cytokines Is Distinct from Genuine Apoptosis. PLoS ONE 2011, 6, e22485. [Google Scholar] [CrossRef]
- Obanda, D.N.; Ribnicky, D.M.; Raskin, I.; Cefalu, W.T. Bioactives of Artemisia dracunculus L. enhance insulin sensitivity by modulation of ceramide metabolism in rat skeletal muscle cells. Nutrition 2014, 30, S59–S66. [Google Scholar] [CrossRef] [Green Version]
- Kheterpal, I.; Scherp, P.; Kelley, L.; Wang, Z.; Johnson, W.; Ribnicky, D.; Cefalu, W.T. Bioactives from Artemisia dracunculus L. enhance insulin sensitivity via modulation of skeletal muscle protein phosphorylation. Nutrition 2014, 30, S43–S51. [Google Scholar] [CrossRef] [Green Version]
- Beg, A.A.; Finco, T.S.; Nantermet, P.V.; Baldwin, A.S., Jr. Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of I kappa B alpha: A mechanism for NF-kappa B activation. Mol. Cell. Biol. 1993, 13, 3301–3310. [Google Scholar]
- Vane, J.R.; Botting, R.M. The mechanism of action of aspirin. Thromb. Res. 2003, 110, 255–258. [Google Scholar] [CrossRef]
- Witters, L.A. The blooming of the French lilac. J. Clin. Investig. 2001, 108, 1105–1107. [Google Scholar] [CrossRef]
- Jack, D.B. One hundred years of aspirin. Lancet 1997, 350, 437–439. [Google Scholar] [CrossRef]
- Kumar, S.; Boehm, J.; Lee, J.C. p38 MAP kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–726. [Google Scholar] [CrossRef]
- Salehi, B.; Quispe, C.; Chamkhi, I.; El Omari, N.; Balahbib, A.; Sharifi-Rad, J.; Bouyahya, A.; Akram, M.; Iqbal, M.; Docea, A.O.; et al. Pharmacological Properties of Chalcones: A Review of Preclinical Including Molecular Mechanisms and Clinical Evidence. Front. Pharmacol. 2020, 11, 592654. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.; Attah, E.I.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Bioprecursors of Flavonoids: Chemistry, Bioactivities, and Pharmacokinetics. Molecules 2021, 26, 7177. [Google Scholar] [CrossRef]
- Meng, C.Q.; Zheng, X.S.; Ni, L.; Ye, Z.; Simpson, J.E.; Worsencroft, K.J.; Hotema, M.R.; Weingarten, M.D.; Skudlarek, J.W.; Gilmore, J.M.; et al. Discovery of novel heteroaryl-substituted chalcones as inhibitors of TNF-alpha-induced VCAM-1 expression. Bioorg. Med. Chem. Lett. 2004, 14, 1513–1517. [Google Scholar] [CrossRef]
- Medicherla, S.; Protter, A.A.; Ma, J.Y.; Mangadu, R.; Almirez, R.; Koppelman, B.; Kerr, I.; Navas, T.A.; Movius, F.; Reddy, M.; et al. Preventive and therapeutic potential of p38 alpha-selective mitogen-activated protein kinase inhibitor in nonobese diabetic mice with type 1 diabetes. J. Pharmacol. Exp. Ther. 2006, 318, 99–107. [Google Scholar] [CrossRef]
- Grimes, J.M.; Grimes, K.V. p38 MAPK inhibition: A promising therapeutic approach for COVID-19. J. Mol. Cell. Cardiol. 2020, 144, 63–65. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smoak, P.; Burke, S.J.; Martin, T.M.; Batdorf, H.M.; Floyd, Z.E.; Collier, J.J. Artemisia dracunculus L. Ethanolic Extract and an Isolated Component, DMC2, Ameliorate Inflammatory Signaling in Pancreatic β-Cells via Inhibition of p38 MAPK. Biomolecules 2022, 12, 708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050708
Smoak P, Burke SJ, Martin TM, Batdorf HM, Floyd ZE, Collier JJ. Artemisia dracunculus L. Ethanolic Extract and an Isolated Component, DMC2, Ameliorate Inflammatory Signaling in Pancreatic β-Cells via Inhibition of p38 MAPK. Biomolecules. 2022; 12(5):708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050708
Chicago/Turabian StyleSmoak, Peter, Susan J. Burke, Thomas M. Martin, Heidi M. Batdorf, Z. Elizabeth Floyd, and J. Jason Collier. 2022. "Artemisia dracunculus L. Ethanolic Extract and an Isolated Component, DMC2, Ameliorate Inflammatory Signaling in Pancreatic β-Cells via Inhibition of p38 MAPK" Biomolecules 12, no. 5: 708. https://0-doi-org.brum.beds.ac.uk/10.3390/biom12050708