The Steroidogenesis Inhibitor Finasteride Reduces the Response to Both Stressful and Rewarding Stimuli


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Orchiectomy
2.4. Locomotor Activity
2.5. Defensive Withdrawal
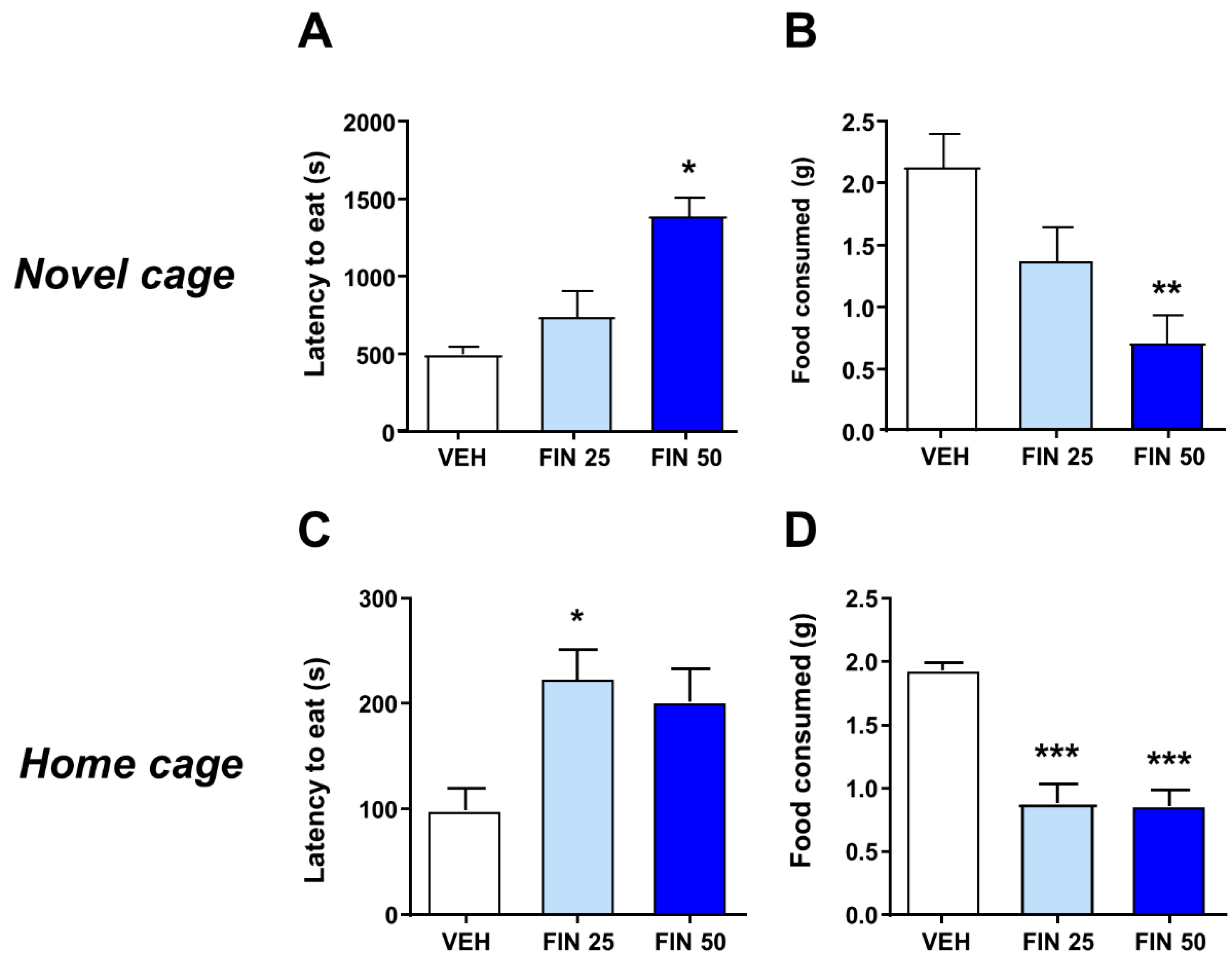
2.6. Novelty-Induced Hypophagia
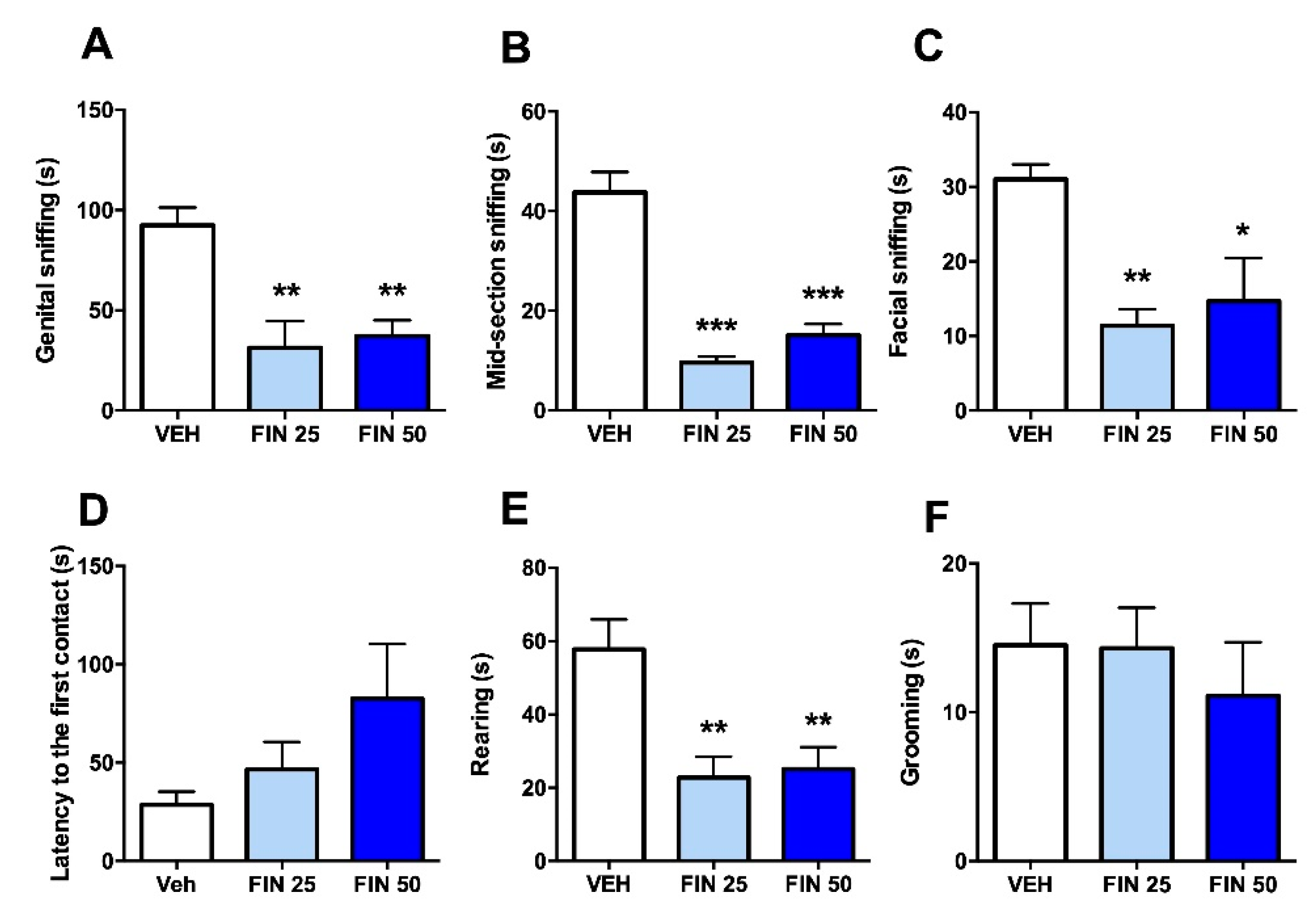
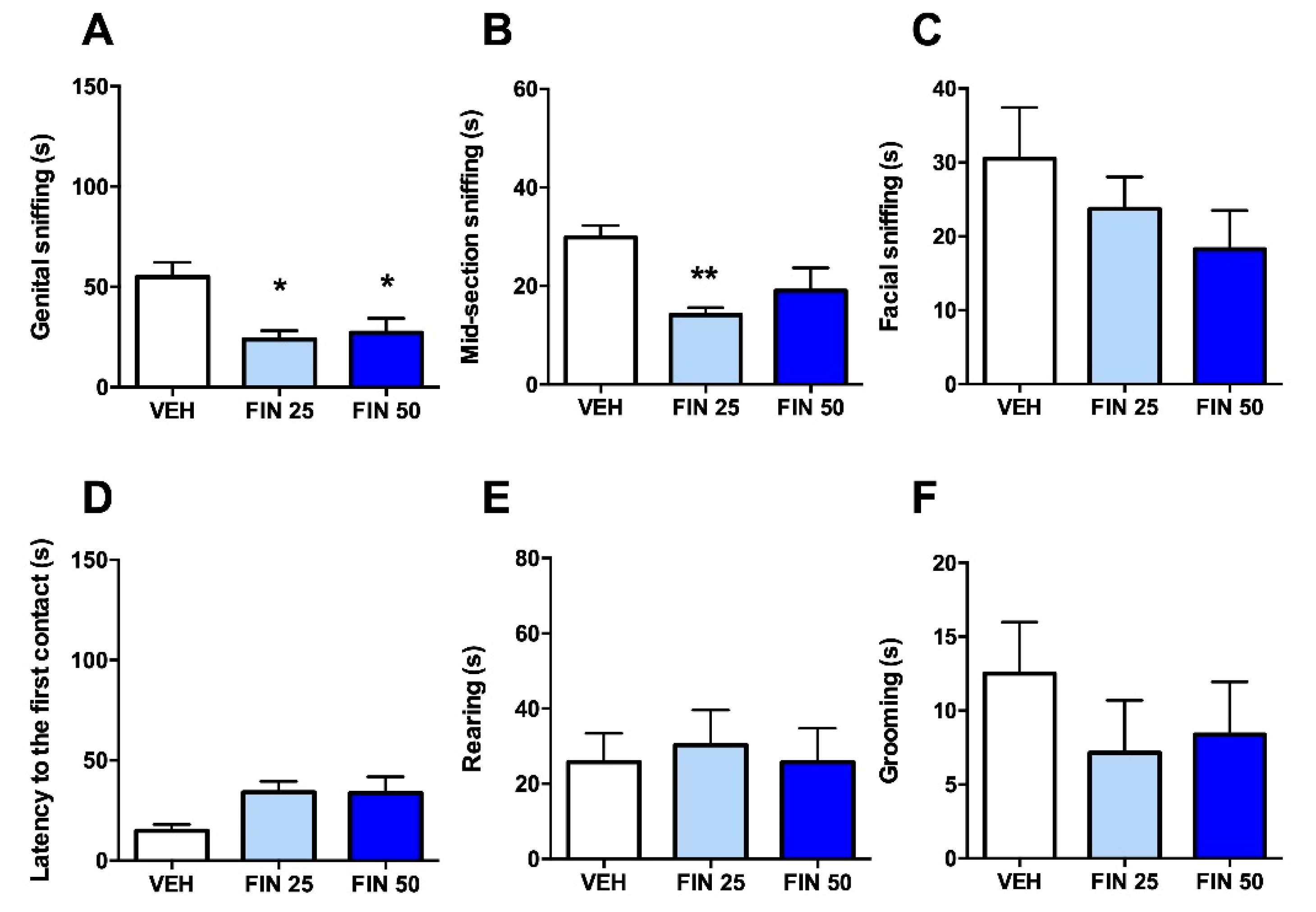
2.7. Social Interaction
2.8. Delay Discounting
2.9. Wire-Beam Bridge Test
2.10. Forced-Swim Test
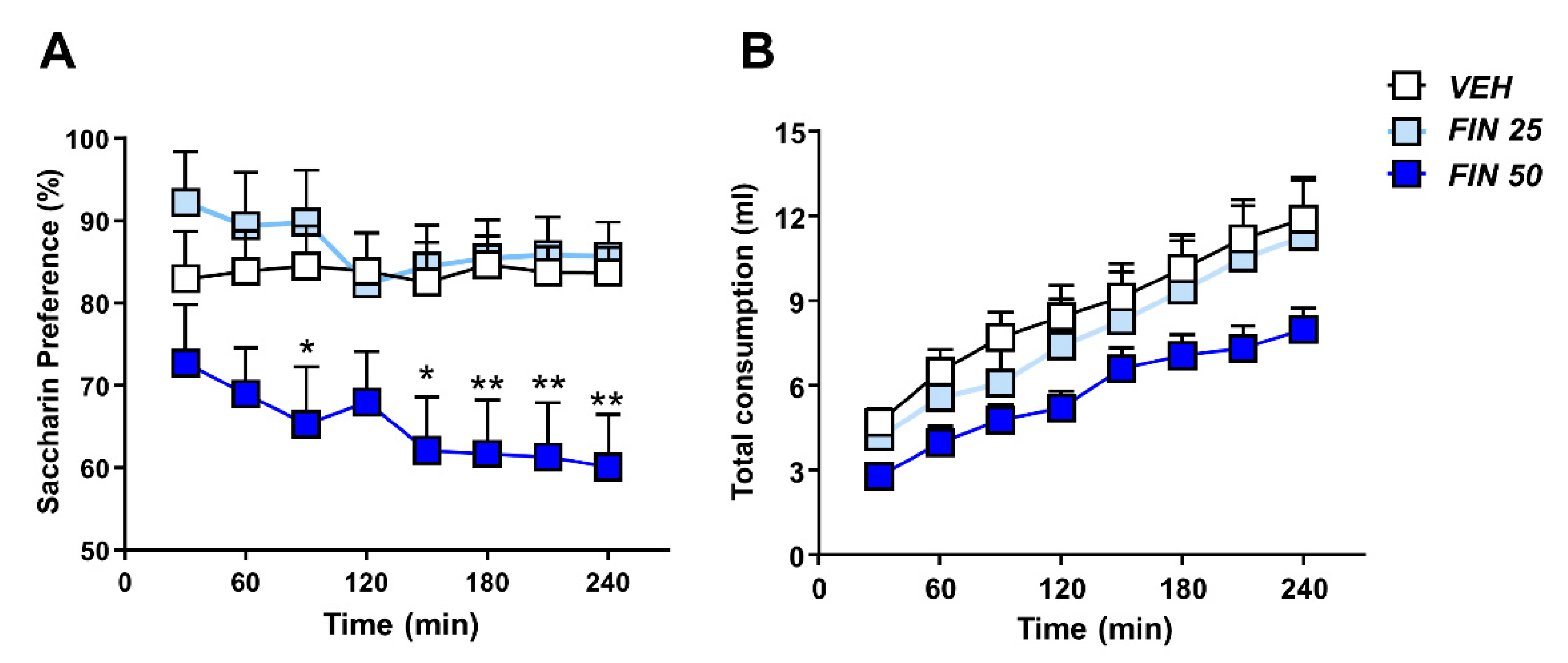
2.11. Saccharin Preference
2.12. Quantification of CRH and ACTH
2.13. Statistical Analyses
3. Results
3.1. FIN Reduced Exploratory and Appetitive Behavior at Doses That Did Not Affect Locomotor Activity
3.2. FIN Reduced Sociability in Rats
3.3. FIN Reduced Impulsivity and Risk-Taking Responses
3.4. FIN Reduced Saccharin Preference
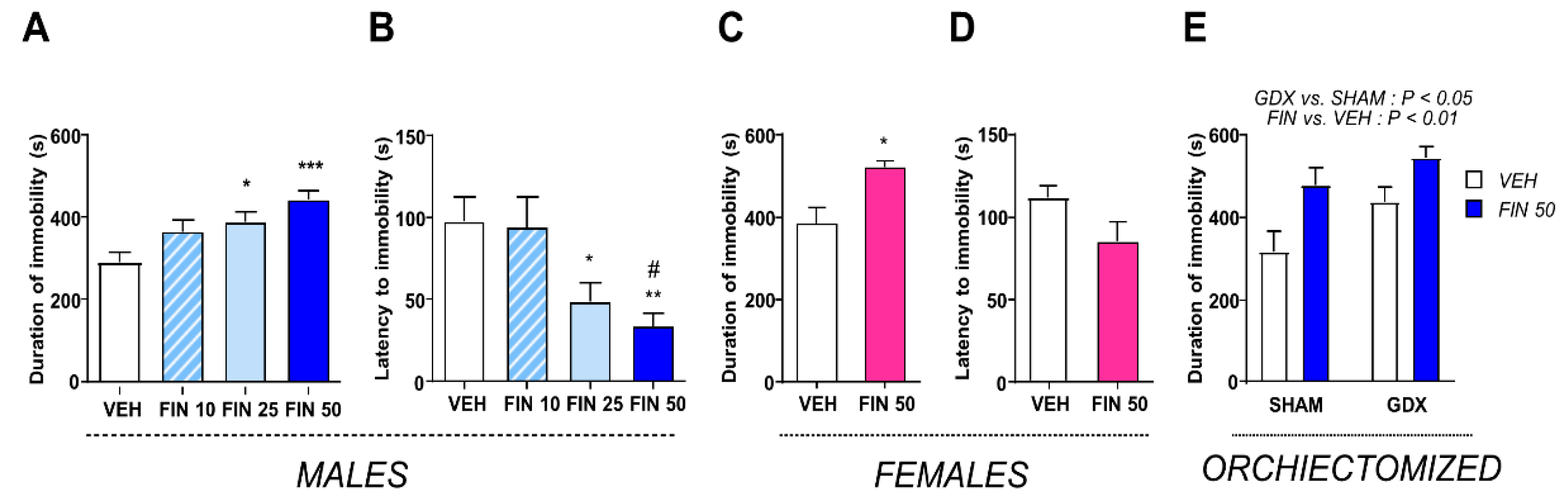
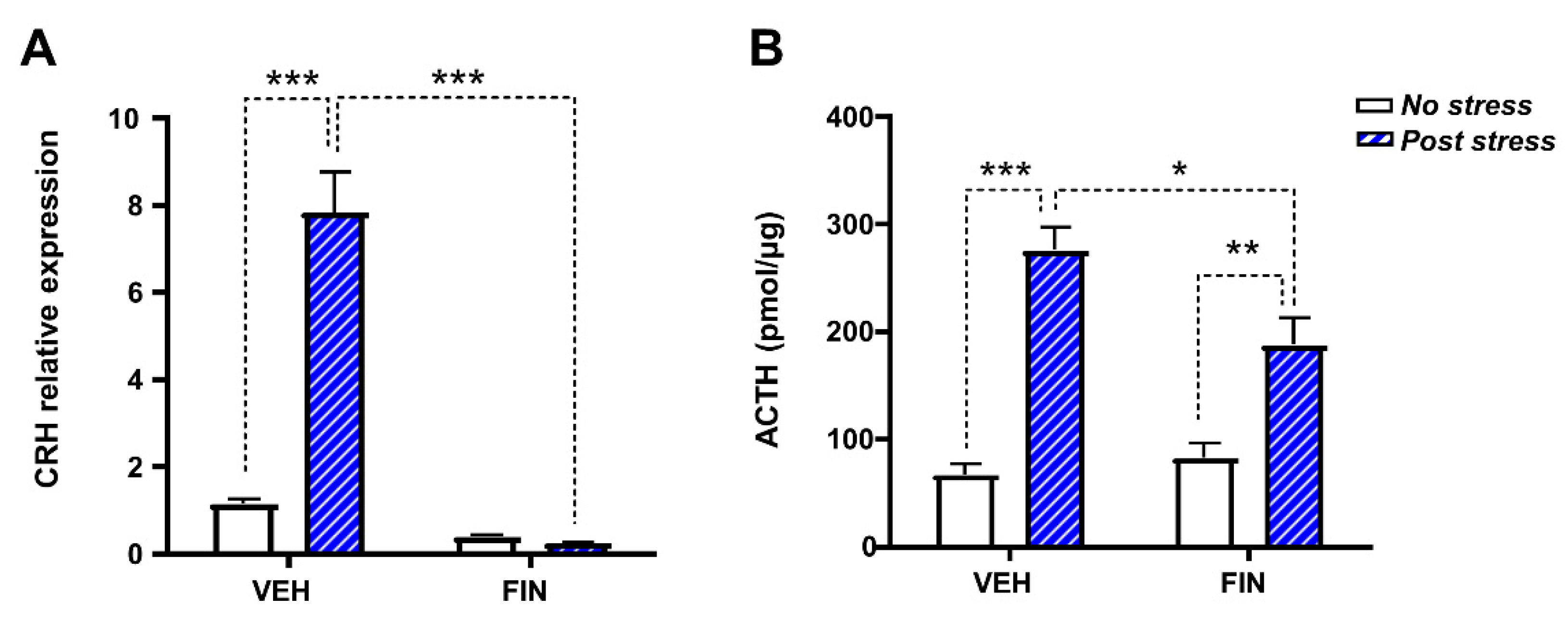
3.5. FIN Reduced Stress Coping Behavior and Suppressed HPA Axis Responses
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Russell, D. Steroid 5-alpha Reductase: Two Genes/Two Enzymes. Annu. Rev. Biochem. 1994, 63, 25–61. [Google Scholar] [CrossRef] [PubMed]
- Paba, S.; Frau, R.C.; Godar, S.; Devoto, P.; Marrosu, F.; Bortolato, M. Steroid 5α-Reductase as a Novel Therapeutic Target for Schizophrenia and Other Neuropsychiatric Disorders. Curr. Pharm. Des. 2011, 17, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.D. Neurosteroids: Endogenous bimodal modulators of the GABAA receptor mechanism of action and physiological significance. Prog. Neurobiol. 1992, 38, 379–395. [Google Scholar] [CrossRef]
- Reddy, D.S.; Jian, K. The testosterone-derived neurosteroid androstanediol is a positive allosteric modulator of GABAA receptors. J. Pharmacol. Exp. Ther. 2010, 334, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Eser, D.; Schüle, C.; Baghai, T.C.; Romeo, E.; Rupprecht, R. Neuroactive steroids in depression and anxiety disorders: Clinical studies. Neuroendocrinology 2007, 84, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Schüle, C.; Baghai, T.; Laakmann, G. Mirtazapine Decreases Stimulatory Effects of Reboxetine on Cortisol, Adrenocorticotropin and Prolactin Secretion in Healthy Male Subjects. Neuroendocrinology 2004, 79, 54–62. [Google Scholar] [CrossRef]
- Reddy, D.S. Is there a physiological role for the neurosteroid THDOC in stress-sensitive conditions? Trends Pharmacol. Sci. 2003, 24, 103–106. [Google Scholar] [CrossRef]
- Frye, C.A.; Duncan, J.E.; Basham, M.; Erskine, M.S. Behavioral effects of 3a-androstanediol II: Hypothalamic and preoptic area actions via a GABAergic mechanism. Behav. Brain Res. 1996, 79, 119–130. [Google Scholar] [CrossRef]
- Frye, C.A.; Van Keuren, K.R.; Erskine, M.S. Behavioral effects of 3α-androstanediol I: Modulation of sexual receptivity and promotion of GABA-stimulated chloride flux. Behav. Brain Res. 1996, 79, 109–118. [Google Scholar] [CrossRef]
- McInnes, K.J.; Kenyon, C.J.; Chapman, K.E.; Livingstone, D.E.W.; Macdonald, L.J.; Walker, B.R.; Andrew, R. 5α-reduced glucocorticoids, novel endogenous activators of the glucocorticoid receptor. J. Biol. Chem. 2004, 279, 22908–22912. [Google Scholar] [CrossRef]
- Uygur, M.C.; Arik, A.I.; Altuğ, U.; Erol, D. Effects of the 5 alpha-reductase inhibitor finasteride on serum levels of gonadal, adrenal, and hypophyseal hormones and its clinical significance: A prospective clinical study. Steroids 1998, 63, 208–213. [Google Scholar] [CrossRef]
- Drake, L.; Hordinsky, M.; Fiedler, V.; Swinehart, J.; Unger, W.P.; Cotterill, P.C.; Thiboutot, D.M.; Lowe, N.; Jacobson, C.; Whiting, D.; et al. The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. J. Am. Acad. Dermatol. 1999, 41, 550–554. [Google Scholar] [PubMed]
- Mukai, Y.; Higashi, T.; Nagura, Y.; Shimada, K. Studies on neurosteroids XXV. Influence of a 5α-reductase inhibitor, finasteride, on rat brain neurosteroid levels and metabolism. Biol. Pharm. Bull. 2008, 31, 1646–1650. [Google Scholar] [CrossRef] [PubMed]
- Concas, A.; Mostallino, M.C.; Porcu, P.; Follesa, P.; Barbaccia, M.L.; Trabucchi, M.; Purdy, R.H.; Grisenti, P.; Biggio, G. Role of brain allopregnanolone in the plasticity of γ-aminobutyric acid type A receptor in rat brain during pregnancy and after delivery. Proc. Natl. Acad. Sci. USA 1998, 95, 13284–13289. [Google Scholar] [CrossRef]
- Dusková, M.; Hill, M.; Hanus, M.; Matousková, M.; Stárka, L. Finasteride treatment and neuroactive steroid formation. Prague Med. Rep. 2009, 110, 222–230. [Google Scholar]
- Altomare, G.; Capella, G.L. Depression Circumstantially Related to the Administration of Finasteride for Androgenetic Alopecia. J. Dermatol. 2002, 29, 665–669. [Google Scholar] [CrossRef]
- Rahimi-Ardabili, B.; Pourandarjani, R.; Habibollahi, P.; Mualeki, A. Finasteride induced depression: A prospective study. BMC Clin. Pharmacol. 2006, 6, 7. [Google Scholar] [CrossRef]
- Traish, A.M.; Melcangi, R.C.; Bortolato, M.; Garcia-Segura, L.M.; Zitzmann, M. Adverse effects of 5α-reductase inhibitors: What do we know, don’t know, and need to know? Rev. Endocr. Metab. Disord. 2015, 16, 177–198. [Google Scholar] [CrossRef]
- Irwig, M.S. Depressive Symptoms and Suicidal Thoughts Among Former Users of Finasteride with Persistent Sexual Side Effects. J. Clin. Psychiatry 2012, 73, 1220–1223. [Google Scholar] [CrossRef]
- Melcangi, R.C.; Caruso, D.; Abbiati, F.; Giatti, S.; Calabrese, D.; Piazza, F.; Cavaletti, G. Neuroactive Steroid Levels are Modified in Cerebrospinal Fluid and Plasma of Post-Finasteride Patients Showing Persistent Sexual Side Effects and Anxious/Depressive Symptomatology. J. Sex. Med. 2013, 10, 2598–2603. [Google Scholar] [CrossRef]
- Ganzer, C.A.; Jacobs, A.R.; Iqbal, F. Persistent Sexual, Emotional, and Cognitive Impairment Post-Finasteride. Am. J. Mens. Health 2015, 9, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Ganzer, C.A.; Jacobs, A.R. Emotional Consequences of Finasteride: Fool’s Gold. Am. J. Mens. Health 2018, 12, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.L.; Semla, T.P. Post-finasteride syndrome. BMJ 2019, 366, l5047. [Google Scholar] [CrossRef] [PubMed]
- Muroni, A.; Paba, S.; Puligheddu, M.; Marrosu, F.; Bortolato, M. A preliminary study of finasteride in Tourette syndrome. Mov. Disord. 2011, 26, 2146–2147. [Google Scholar] [CrossRef]
- Bortolato, M.; Cannas, A.; Solla, P.; Bini, V.; Puligheddu, M.; Marrosu, F. Finasteride Attenuates Pathological Gambling in Patients With Parkinson Disease. J. Clin. Psychopharmacol. 2012, 32, 424–425. [Google Scholar] [CrossRef]
- Devoto, P.; Frau, R.; Bini, V.; Pillolla, G.; Saba, P.; Flore, G.; Corona, M.; Marrosu, F.; Bortolato, M. Inhibition of 5α-reductase in the nucleus accumbens counters sensorimotor gating deficits induced by dopaminergic activation. Psychoneuroendocrinology 2012, 37, 1630–1645. [Google Scholar] [CrossRef]
- Pes, R.; Godar, S.C.; Fox, A.T.; Burgeno, L.M.; Strathman, H.J.; Jarmolowicz, D.P.; Devoto, P.; Levant, B.; Phillips, P.E.; Fowler, S.C.; et al. Pramipexole enhances disadvantageous decision-making: Lack of relation to changes in phasic dopamine release. Neuropharmacology 2017, 114, 77–87. [Google Scholar] [CrossRef]
- Bortolato, M.; Campolongo, P.; Mangieri, R.A.; Scattoni, M.L.; Frau, R.; Trezza, V.; La Rana, G.; Russo, R.; Calignano, A.; Gessa, G.L.; et al. Anxiolytic-Like Properties of the Anandamide Transport Inhibitor AM404. Neuropsychopharmacology 2006, 31, 2652–2659. [Google Scholar] [CrossRef]
- Tanda, G.; Di Chiara, G. A dopamine-μ 1 opioid link in the rat ventral tegmentum shared by palatable food (Fonzies) and non-psychostimulant drugs of abuse. Eur. J. Neurosci. 1998, 10, 1179–1187. [Google Scholar] [CrossRef]
- Bortolato, M.; Bini, V.; Frau, R.; Devoto, P.; Pardu, A.; Fan, Y.; Solbrig, M.V. Juvenile cannabinoid treatment induces frontostriatal gliogenesis in Lewis rats. Eur. Neuropsychopharmacol. 2014, 24, 974–985. [Google Scholar] [CrossRef]
- Evenden, J.L.; Ryan, C.N. The pharmacology of impulsive behaviour in rats: The effects of drugs on response choice with varying delays of reinforcement. Psychopharmacology. 1996, 128, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Frau, R.; Bini, V.; Soggiu, A.; Scheggi, S.; Pardu, A.; Fanni, S.; Roncada, P.; Puligheddu, M.; Marrosu, F.; Caruso, D.; et al. The Neurosteroidogenic Enzyme 5α-Reductase Mediates Psychotic-Like Complications of Sleep Deprivation. Neuropsychopharmacology 2017, 42, 2196–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, G.; Bambico, F.R.; Mangieri, R.; Bortolato, M.; Campolongo, P.; Solinas, M.; Cassano, T.; Morgese, M.G.; Debonnel, G.; Duranti, A.; et al. Antidepressant-like activity and modulation of brain monoaminergic transmission by blockade of anandamide hydrolysis. Proc. Natl. Acad. Sci. USA 2005, 102, 18620–18625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Muma, N.A.; Battaglia, G.; Van de Kar, L.D. A desensitization of hypothalamic 5-HT1A receptors by repeated injections of paroxetine: Reduction in the levels of G(i) and G(o) proteins and neuroendocrine responses, but not in the density of 5-HT1A receptors. J. Pharmacol. Exp. Ther. 1997, 282, 1581–1590. [Google Scholar]
- McAllister, C.E.; Mi, Z.; Mure, M.; Li, Q.; Muma, N.A. GPER1 Stimulation Alters Posttranslational Modification of RGSz1 and Induces Desensitization of 5-HT 1A Receptor Signaling in the Rat Hypothalamus. Neuroendocrinology 2014, 100, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.-Q.; Kawashima, H.; Iwasaki, Y.; Uchida, K.; Sugimoto, K.; Itoi, K. Differential effects of forced swim-stress on the corticotropin-releasing hormone and vasopressin gene transcription in the parvocellular division of the paraventricular nucleus of rat hypothalamus. Neurosci. Lett. 2004, 358, 201–204. [Google Scholar] [CrossRef]
- Jutkiewicz, E.M.; Wood, S.K.; Houshyar, H.; Hsin, L.W.; Rice, K.C.; Woods, J.H. The effects of CRF antagonists, antalarmin, CP154,526, LWH234, and R121919, in the forced swim test and on swim-induced increases in adrenocorticotropin in rats. Psychopharmacology 2005, 180, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheggi, S.; De Montis, M.G.; Gambarana, C. Making Sense of Rodent Models of Anhedonia. Int. J. Neuropsychopharmacol. 2018, 21, 1049–1065. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Aldridge, J.W.; LaPorte, J.L.; Murphy, D.L.; Tuohimaa, P. Analyzing grooming microstructure in neurobehavioral experiments. Nat. Protoc. 2007, 2, 2538–2544. [Google Scholar] [CrossRef]
- Bortolato, M.; Muroni, A.; Marrosu, F. Treatment of Tourette’s Syndrome with Finasteride. Am. J. Psychiatry 2007, 164, 1914–1915. [Google Scholar] [CrossRef]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The Rodent Forced Swim Test Measures Stress-Coping Strategy, Not Depression-like Behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overstreet, D.H. Modeling depression in animal models. Methods Mol. Biol. 2012, 829, 125–144. [Google Scholar] [PubMed]
- Walf, A.A.; Kaurejo, S.; Frye, C.A. Research Brief: Self-Reports of a Constellation of Persistent Antiandrogenic, Estrogenic, Physical, and Psychological Effects of Finasteride Usage Among Men. Am. J. Mens Health. 2018, 12, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Ciotta, L.; Cianci, A.; Calogero, A.E.; Palumbo, M.A.; Marletta, E.; Sciuto, A.; Palumbo, G. Clinical and endocrine effects of finasteride, a 5α-reductase inhibitor, in women with idiopathic hirsutism. Fertil. Steril. 1995, 64, 299–306. [Google Scholar] [CrossRef]
- Lakryc, E.M.; Motta, E.L.A.; Soares, J.M.; Haidar, M.A.; Rodrigues de Lima, G.; Baracat, E.C. The benefits of finasteride for hirsute women with polycystic ovary syndrome or idiopathic hirsutism. Gynecol. Endocrinol. 2003, 17, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. Finasteride Treatment of Patterned Hair Loss in Normoandrogenic Postmenopausal Women. Dermatology 2004, 209, 202–207. [Google Scholar] [CrossRef]
- Iorizzo, M.; Vincenzi, C.; Voudouris, S.; Piraccini, B.M.; Tosti, A. Finasteride Treatment of Female Pattern Hair Loss. Arch. Dermatol. 2006, 142, 298–302. [Google Scholar] [CrossRef]
- Makhlouf, A.A.; Mohamed, M.A.; Seftel, A.D.; Niederberger, C. Hypogonadism is associated with overt depression symptoms in men with erectile dysfunction. Int. J. Impot. Res. 2008, 20, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, J.; Wakefield, S.; MacKenzie, G.; Moss, S.J.; Maguire, J. Neurosteroidogenesis is required for the physiological response to stress: Role of neurosteroid-sensitive GABAA receptors. J. Neurosci. 2011, 31, 18198–18210. [Google Scholar] [CrossRef] [Green Version]
- Melón, L.C.; Maguire, J. GABAergic regulation of the HPA and HPG axes and the impact of stress on reproductive function. J. Steroid Biochem. Mol. Biol. 2016, 160, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.N.; Reid, R.L.; Van Vugt, D.A. The effects of estrogen and progesterone on corticotropin-releasing hormone and arginine vasopressin messenger ribonucleic acid levels in the paraventricular nucleus and supraoptic nucleus of the rhesus monkey. Endocrinology. 1999, 140, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Dallman, M.F.; Yates, F.E. Dynamic asymmetries in the corticosteroid feedback path and distribution-metabolism-binding elements of the adrenocortical system. Ann. Acad. Sci. 1969, 156, 696–721. [Google Scholar] [CrossRef] [PubMed]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; Figueiredo, H.; Mueller, N.K.; Ulrich-Lai, Y.; Ostrander, M.M.; Choi, D.C.; Cullinan, W.E. Central mechanisms of stress integration: Hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front. Neuroendocrinol. 2003, 24, 151–180. [Google Scholar] [CrossRef]
- Bale, T.L.; Vale, W.W. CRF and CRF receptors: Role in Stress Responsivity and Other Behaviors. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 525–557. [Google Scholar] [CrossRef]
- García-Lecumberri, C.; Ambrosio, E. Role of corticotropin-releasing factor in forced swimming test. Eur. J. Pharmacol. 1998, 343, 17–26. [Google Scholar] [CrossRef]
- Zebrowska-Łupina, I.; Pietrasiewicz, T.; Ossowska, G.; Łupina, T.; Klenk-Majewska, B. ACTH 4-9 analogue facilitates the antiimmobility effect of antidepressants and dopamine agonists in swimming rats. J. Physiol. Pharmacol. 1997, 48, 263–275. [Google Scholar]
- Korte, S.M.; Bouws, G.A.; Bohus, B. Central actions of corticotropin-releasing hormone (CRH) on behavioral, neuroendocrine, and cardiovascular regulation: Brain corticoid receptor involvement. Horm Behav. 1993, 27, 167–183. [Google Scholar] [CrossRef]
- Shibasaki, T.; Yamauchi, N.; Hotta, M.; Imaki, T.; Oda, T.; Ling, N.; Demura, H. Brain corticotropin-releasing hormone increases arousal in stress. Brain Res. 1991, 554, 352–354. [Google Scholar] [CrossRef]
- Dunn, A.J.; Berridge, C.W. Physiological and behavioral responses to corticotropin-releasing factor administration: Is CRF a mediator of anxiety or stress responses? Brain Res. Rev. 1990, 15, 71–100. [Google Scholar] [CrossRef]
- Bortolato, M.; Frau, R.; Orru, M.; Bourov, Y.; Marrosu, F.; Mereu, G.; Devoto, P.; Gessa, G.L. Antipsychotic-like properties of 5-alpha-reductase inhibitors. Neuropsychopharmacology 2008, 33, 3146–3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frau, R.; Mosher, L.J.; Bini, V.; Pillolla, G.; Pes, R.; Saba, P.; Fanni, S.; Devoto, P.; Bortolato, M. The neurosteroidogenic enzyme 5α-reductase modulates the role of D1 dopamine receptors in rat sensorimotor gating. Psychoneuroendocrinology 2016, 63, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soggiu, A.; Piras, C.; Greco, V.; Devoto, P.; Urbani, A.; Calzetta, L.; Bortolato, M.; Roncada, P. Exploring the neural mechanisms of finasteride: A proteomic analysis in the nucleus accumbens. Psychoneuroendocrinology 2016, 74, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feenstra, M.G.; Botterblom, M.H.; van Uum, J.F. Behavioral arousal and increased dopamine efflux after blockade of NMDA-receptors in the prefrontal cortex are dependent on activation of glutamatergic neurotransmission. Neuropharmacology 2002, 42, 752–763. [Google Scholar] [CrossRef]
- Horvitz, J.C. Mesolimbocortical and nigrostriatal dopamine responses to salient non-reward events. Neuroscience. 2000, 96, 651–656. [Google Scholar] [CrossRef]
- Sasibhushana, R.B.; Shankaranarayana Rao, B.S.; Srikumar, B.N. Repeated finasteride administration induces depression-like behavior in adult male rats. Behav. Brain Res. 2019, 365, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Mook, R.; Moss, M.L.; Frye, S.V. Mechanism of Time-Dependent Inhibition of 5.alpha.-Reductases by Δ1-4-Azasteroids: Toward Perfection of Rates of Time-Dependent Inhibition by Using Ligand-Binding Energies. Biochemistry 1995, 34, 13453–13459. [Google Scholar] [CrossRef]
- Thigpen, A.E.; Russell, D.W. Four-amino acid segment in steroid 5 alpha-reductase 1 confers sensitivity to finasteride, a competitive inhibitor. J. Biol. Chem. 1992, 267, 8577–8583. [Google Scholar]
- Eicheler, W.; Tuohimaa, P.; Vilja, P.; Adermann, K.; Forssmann, W.G.; Aumüller, G. Immunocytochemical localization of human 5 alpha-reductase 2 with polyclonal antibodies in androgen target and non-target human tissues. J. Histochem. Cytochem. 1994, 42, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Aumuller, G.; Eicheler, W.; Renneberg, H.; Adermann, K.; Vilja, P.; Forssmann, W.G. Immunocytochemical evidence for differential subcellular localization of 5 alpha-reductase isoenzymes in human tissues. Cells Tissues Organs 1996, 156, 241–252. [Google Scholar] [CrossRef]
- Azzouni, F.; Godoy, A.; Li, Y.; Mohler, J. The 5 Alpha-Reductase Isozyme Family: A Review of Basic Biology and Their Role in Human Diseases. Adv. Urol. 2012, 2012, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, M.P.; Casti, A.; Casu, A.; Frau, R.; Bortolato, M.; Spiga, S.; Ennas, M.G. Regional distribution of 5α-reductase type 2 in the adult rat brain: An immunohistochemical analysis. Psychoneuroendocrinology 2013, 38, 281–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolato, M.; Devoto, P.; Roncada, P.; Frau, R.; Flore, G.; Saba, P.; Pistritto, G.; Soggiu, A.; Pisanu, S.; Zappala, A.; et al. Isolation rearing-induced reduction of brain 5α-reductase expression: Relevance to dopaminergic impairments. Neuropharmacology. 2011, 60, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Diviccaro, S.; Giatti, S.; Borgo, F.; Barcella, M.; Borghi, E.; Trejo, J.L.; Garcia-Segura, L.M.; Melcangi, R.C. Treatment of male rats with finasteride, an inhibitor of 5alpha-reductase enzyme, induces long-lasting effects on depressive-like behavior, hippocampal neurogenesis, neuroinflammation and gut microbiota composition. Psychoneuroendocrinology 2019, 99, 206–215. [Google Scholar] [CrossRef]
- Uzunova, V.; Sheline, Y.; Davis, J.M.; Rasmusson, A.; Uzunov, D.P.; Costa, E.; Guidotti, A. Increase in the cerebrospinal fluid content of neurosteroids in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine. Proc. Natl. Acad. Sci. USA 1998, 95, 3239–3244. [Google Scholar] [CrossRef] [Green Version]
- Dong, E.; Matsumoto, K.; Uzunova, V.; Sugaya, I.; Takahata, H.; Nomura, H.; Watanabe, H.; Costa, E.; Guidotti, A. Brain 5 -dihydroprogesterone and allopregnanolone synthesis in a mouse model of protracted social isolation. Proc. Natl. Acad. Sci. USA 2001, 98, 2849–2854. [Google Scholar] [CrossRef] [Green Version]
- Schüle, C.; Nothdurfter, C.; Rupprecht, R. The role of allopregnanolone in depression and anxiety. Prog. Neurobiol. 2014, 113, 79–87. [Google Scholar] [CrossRef]
- Meltzer-Brody, S.; Colquhoun, H.; Riesenberg, R.; Epperson, C.N.; Deligiannidis, K.M.; Rubinow, D.R.; Li, H.; Sankoh, A.J.; Clemson, C.; Schacterle, A.; et al. Brexanolone injection in post-partum depression: Two multicentre, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet 2018, 392, 1058–1070. [Google Scholar] [CrossRef]
- Frye, C.A.; Walf, A.A. Depression-like behavior of aged male and female mice is ameliorated with administration of testosterone or its metabolites. Physiol. Behav. 2009, 97, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Frye, C.A.; Edinger, K.L.; Lephart, E.D.; Walf, A.A. 3alpha-androstanediol, but not testosterone, attenuates age-related decrements in cognitive, anxiety, and depressive behavior of male rats. Front. Aging Neurosci. 2010, 2, 15. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godar, S.C.; Cadeddu, R.; Floris, G.; Mosher, L.J.; Mi, Z.; Jarmolowicz, D.P.; Scheggi, S.; Walf, A.A.; Koonce, C.J.; Frye, C.A.; et al. The Steroidogenesis Inhibitor Finasteride Reduces the Response to Both Stressful and Rewarding Stimuli. Biomolecules 2019, 9, 749. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110749
Godar SC, Cadeddu R, Floris G, Mosher LJ, Mi Z, Jarmolowicz DP, Scheggi S, Walf AA, Koonce CJ, Frye CA, et al. The Steroidogenesis Inhibitor Finasteride Reduces the Response to Both Stressful and Rewarding Stimuli. Biomolecules. 2019; 9(11):749. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110749
Chicago/Turabian StyleGodar, Sean C., Roberto Cadeddu, Gabriele Floris, Laura J. Mosher, Zhen Mi, David P. Jarmolowicz, Simona Scheggi, Alicia A. Walf, Carolyn J. Koonce, Cheryl A. Frye, and et al. 2019. "The Steroidogenesis Inhibitor Finasteride Reduces the Response to Both Stressful and Rewarding Stimuli" Biomolecules 9, no. 11: 749. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9110749