Plastrum Testudinis Extract Mitigates Thiram Toxicity in Broilers via Regulating PI3K/AKT Signaling


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Compounds
2.2. Approval of Ethics
2.3. Experimental Plan
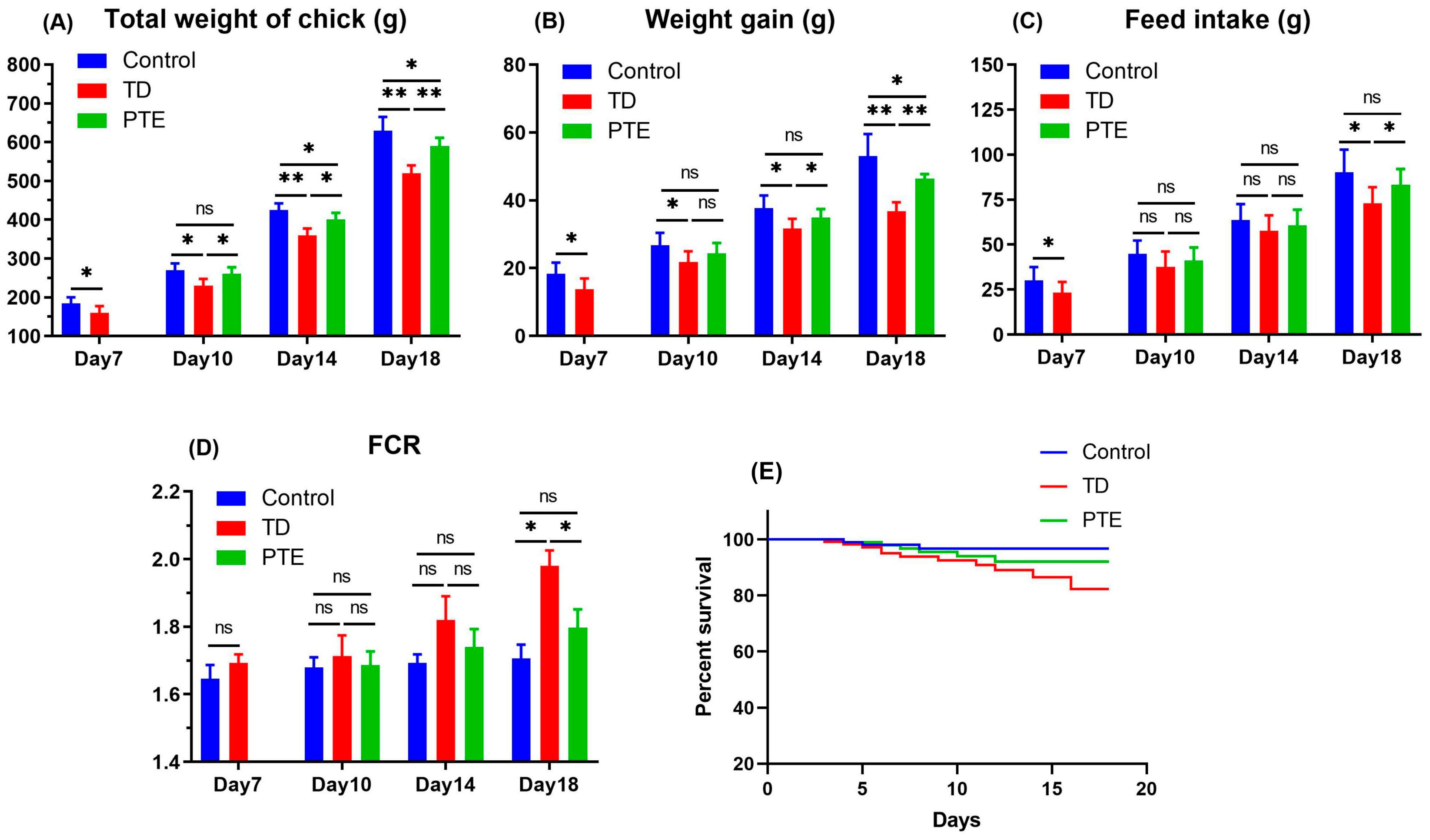
2.4. Analyses of Production Parameters and Tibia Parameters
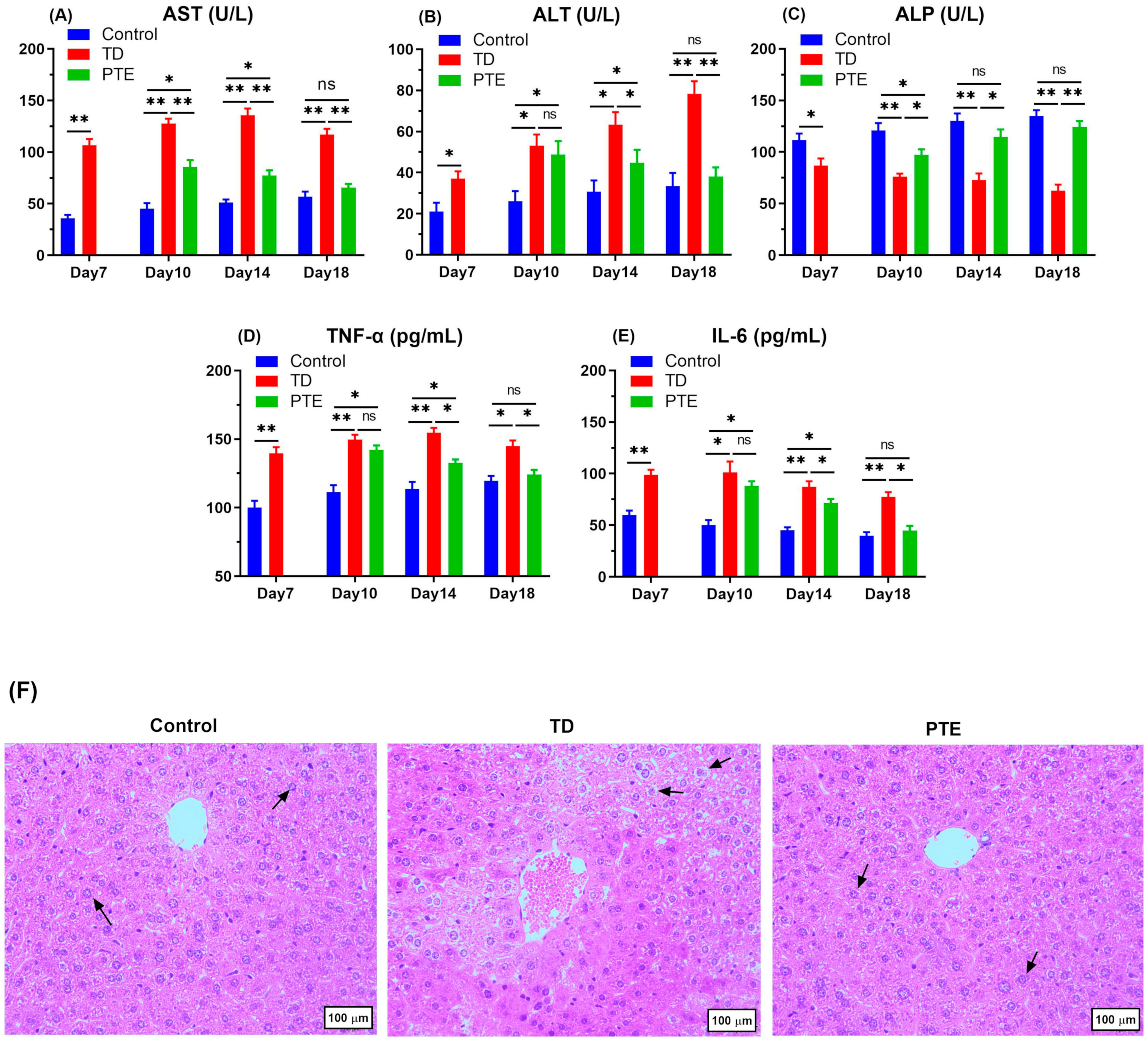
2.5. Serum Biochemicals Analysis
2.6. Liver Weight and Antioxidants Analysis
2.7. Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry
2.8. RNA Extraction and RT-qPCR
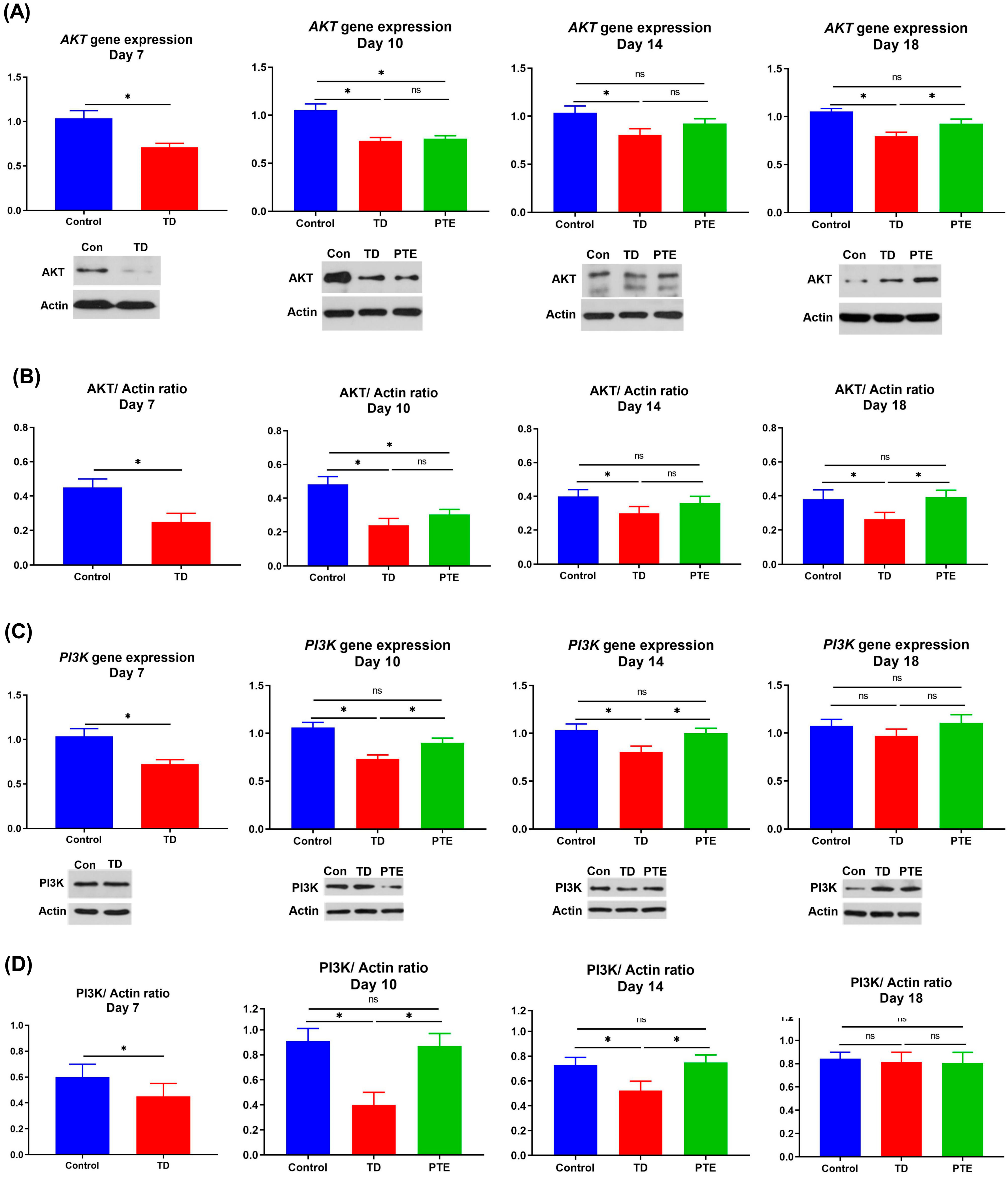
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. Production Parameters
3.2. Effect of PTE on Growth Plate Size, Tibia Parameters, and TD Score
3.3. Biochemical Criterion Analysis of Serum and Liver Histopathology
3.4. Liver Weight and Determination of Antioxidant Status in Liver
3.5. Histopathological Investigation of the Growth Plates of the Tibia
3.6. Immunohistochemistry of Tibial Growth Plates
3.7. AKT and PI3K Genes and Protein Expression in the Growth Plate
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osman, A.G.; Sherif, A.M.; Elhussein, A.A.; Mohamed, A.T. Sensitivity of some nitrogen fixers and the target pest Fusarium oxysporum to fungicide thiram. Interdiscip. Toxicol. 2012, 5, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Bencko, V.; Yan Li Foong, F. The history of arsenical pesticides and health risks related to the use of Agent Blue. Ann. Agric. Environ. Med. AAEM 2017, 24, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Kunkur, V.; Hunje, R.; Biradarpatil, N.K.; Vyakarnahal, B.S. Effect of seed coating with polymer, fungicide and insecticide on seed quality in cotton during storage. Karnataka J. Agric. Sci. 2007, 20, 137–139. [Google Scholar]
- Tian, W.-X.; Li, J.-K.; Qin, P.; Wang, R.; Ning, G.-B.; Qiao, J.-G.; Li, H.-Q.; Bi, D.-R.; Pan, S.-Y.; Guo, D.-Z. Screening of differentially expressed genes in the growth plate of broiler chickens with tibial dyschondroplasia by microarray analysis. BMC Genom. 2013, 14, 276. [Google Scholar] [CrossRef]
- Rath, N.; Richards, M.; Huff, W.; Huff, G.; Balog, J. Changes in the tibial growth plates of chickens with thiram-induced dyschondroplasia. J. Comp. Pathol. 2005, 133, 41–52. [Google Scholar] [CrossRef]
- Zhang, H.; Mehmood, K.; Jiang, X.; Yao, W.; Iqbal, M.; Waqas, M.; Rehman, M.U.; Li, A.; Shen, Y.; Li, J. Effect of tetramethyl thiuram disulfide (thiram) in relation to tibial dyschondroplasia in chickens. Environ. Sci. Pollut. Res. Int. 2018, 25, 28264–28274. [Google Scholar] [CrossRef]
- Orth, M.W.; Cook, M.E. Avian tibial dyschondroplasia: A morphological and biochemical review of the growth plate lesion and its causes. Vet. Pathol. 1994, 31, 403–404. [Google Scholar] [CrossRef]
- Rath, N.C.; Huff, W.E.; Balog, J.M.; Huff, G.R. Comparative efficacy of different dithiocarbamates to induce tibial dyschondroplasia in poultry 1. Poult. Sci. 2004, 83, 266–274. [Google Scholar] [CrossRef]
- Nabi, F.; Shahzad, M.; Liu, J.; Li, K.; Han, Z.; Zhang, D.; Iqbal, M.K.; Li, J. Hsp90 inhibitor celastrol reinstates growth plate angiogenesis in thiram-induced tibial dyschondroplasia. Avian Pathol. 2016, 45, 187–193. [Google Scholar] [CrossRef]
- Nabi, F.; Li, K.; Shahzad, M.; Han, Z.; Zhang, D.; Liu, J.; Li, J. Gambogic acid inhibits Hsp90 expressions in thiram-induced tibial dyschondroplasia. Pak. Vet. J. 2016, 36, 224–226. [Google Scholar]
- Jefferies, D.; Houston, B.; Lester, D.; Whitehead, C.C.; Thorp, B.H.; Botman, M.; Farquharson, C. Expression patterns of chondrocyte genes cloned by differential display in tibial dyschondroplasia. Biochim. Biophys. Acta 2000, 1501, 180–188. [Google Scholar] [CrossRef]
- Shahzad, M.; Liu, J.; Gao, J.; Wang, Z.; Zhang, D.; Nabi, F.; Li, J. Hsp-90 inhibitor geldanamycin attenuates liver oxidative stress and toxicity in thiram-induced tibial dyschondroplasia. Pak. Vet. J. 2014, 34, 545–547. [Google Scholar]
- Genin, O.; Hasdai, A.; Shinder, D.; Pines, M. The effect of inhibition of heat-shock proteins on thiram-induced tibial dyschondroplasia. Poult. Sci. 2012, 91, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Rath, N.C.; Huff, W.E.; Huff, G.R. Thiram-induced changes in the expression of genes relating to vascularization and tibial dyschondroplasia. Poult. Sci. 2007, 86, 2390–2395. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Knopov, V.; Genina, O.; Hurwitz, S.; Faerman, A.; Gerstenfeld, L.C.; Leach, R.M. Development of avian tibial dyschondroplasia: Gene expression and protein synthesis. Calcif. Tissue Int. 1998, 63, 521–527. [Google Scholar] [CrossRef]
- Williams, A.; Wang, E.C.; Thurner, L.; Liu, C.J. Review: Novel Insights Into Tumor Necrosis Factor Receptor, Death Receptor 3, and Progranulin Pathways in Arthritis and Bone Remodeling. Arthritis Rheumatol. 2016, 68, 2845–2856. [Google Scholar] [CrossRef]
- Tartaglia, L.A.; Goeddel, D.V. Two TNF receptors. Immunol. Today 1992, 13, 151–153. [Google Scholar] [CrossRef]
- Naude, P.J.; den Boer, J.A.; Luiten, P.G.; Eisel, U.L. Tumor necrosis factor receptor cross-talk. FEBS J. 2011, 278, 888–898. [Google Scholar] [CrossRef]
- Guntur, A.R.; Rosen, C.J. The skeleton: A multi-functional complex organ: New insights into osteoblasts and their role in bone formation: The central role of PI3Kinase. J. Endocrinol. 2011, 211, 123–130. [Google Scholar] [CrossRef]
- Xi, J.C.; Zang, H.Y.; Guo, L.X.; Xue, H.B.; Liu, X.D.; Bai, Y.B.; Ma, Y.Z. The PI3K/AKT cell signaling pathway is involved in regulation of osteoporosis. J. Recept. Signal Transduct. Res. 2015, 35, 640–645. [Google Scholar] [CrossRef]
- Kawamura, N.; Kugimiya, F.; Oshima, Y.; Ohba, S.; Ikeda, T.; Saito, T.; Shinoda, Y.; Kawasaki, Y.; Ogata, N.; Hoshi, K.; et al. Akt1 in osteoblasts and osteoclasts controls bone remodeling. PLoS ONE 2007, 2, e1058. [Google Scholar] [CrossRef]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Mukherjee, A.; Wilson, E.M.; Rotwein, P. Selective Signaling by Akt2 Promotes Bone Morphogenetic Protein 2-Mediated Osteoblast Differentiation. Mol. Cell. Biol. 2010, 30, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Neural Beginnings for the Turtle’s Shell. Science 2004, 303, 951. [Google Scholar] [CrossRef]
- Chen, D.F.; Zeng, H.P.; Du, S.H.; Li, H.; Zhou, J.H.; Li, Y.W.; Wang, T.T.; Hua, Z.C. Extracts from Plastrum testudinis promote proliferation of rat bone-marrow-derived mesenchymal stem cells. Cell Prolif. 2007, 40, 196–212. [Google Scholar] [CrossRef]
- Yang, L.; Tao, T.; Wang, X.; Du, N.; Chen, W.; Tao, S.; Wang, Z.; Wu, L. Effects of dexamethasone on proliferation, differentiation and apoptosis of adult human osteoblasts in vitro. Chin. Med. J. 2003, 116, 1357–1360. [Google Scholar]
- Rocchi, P.; Tonelli, R.; Camerin, C.; Purgato, S.; Fronza, R.; Bianucci, F.; Guerra, F.; Pession, A.; Ferreri, A.M. p21Waf1/Cip1 is a common target induced by short-chain fatty acid HDAC inhibitors (valproic acid, tributyrin and sodium butyrate) in neuroblastoma cells. Oncol. Rep. 2005, 13, 1139–1144. [Google Scholar] [CrossRef]
- Nugent, A.; Roche, H.; Noone, E.; Long, A.; Kelleher, D.; Gibney, M. The effects of conjugated linoleic acid supplementation on immune function in healthy volunteers. Eur. J. Clin. Nutr. 2005, 59, 742. [Google Scholar] [CrossRef]
- Lallès, J.-P. Intestinal alkaline phosphatase: Multiple biological roles in maintenance of intestinal homeostasis and modulation by diet. Nutr. Rev. 2010, 68, 323–332. [Google Scholar] [CrossRef]
- Kunisawa, J.; Hashimoto, E.; Inoue, A.; Nagasawa, R.; Suzuki, Y.; Ishikawa, I.; Shikata, S.; Arita, M.; Aoki, J.; Kiyono, H. Regulation of intestinal IgA responses by dietary palmitic acid and its metabolism. J. Immunol. 2014, 193, 1666–1671. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.Z.; Hu, J.X.; Wu, H.; Li, Y.L.; Chen, H.L.; Bai, H.; Hai, C.X. ROS-activated p38 MAPK/ERK-Akt cascade plays a central role in palmitic acid-stimulated hepatocyte proliferation. Free Radic. Biol. Med. 2011, 51, 539–551. [Google Scholar] [CrossRef]
- Goradel, N.H.; Eghbal, M.A.; Darabi, M.; Roshangar, L.; Asadi, M.; Zarghami, N.; Nouri, M. Improvement of liver cell therapy in rats by dietary stearic acid. Iran. Biomed. J. 2016, 20, 217. [Google Scholar]
- Pan, P.-H.; Lin, S.-Y.; Ou, Y.-C.; Chen, W.-Y.; Chuang, Y.-H.; Yen, Y.-J.; Liao, S.-L.; Raung, S.-L.; Chen, C.-J. Stearic acid attenuates cholestasis-induced liver injury. Biochem. Biophys. Res. Commun. 2010, 391, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Jokelainen, K.; Rahemtulla, A.; Miao, L.; Fogt, F.; Matsumoto, H.; Tahan, S.R.; Su, G.L. Activation of nuclear factor kappa B and cytokine imbalance in experimental alcoholic liver disease in the rat. Hepatology 1999, 30, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Liang, C.L.; Li, G.M.; Yu, C.Y.; Yin, M. Stearic acid protects primary cultured cortical neurons against oxidative stress 4. Acta Pharm. Sin. 2007, 28, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xiu, C.; Zhou, Q.; Ni, L.; Du, J.; Gong, T.; Li, M.; Yang, H.; Chen, J. A dual role of cholesterol in osteogenic differentiation of bone marrow stromal cells. J. Cell. Physiol. 2019, 234, 2058–2066. [Google Scholar] [CrossRef]
- Pinto, M.E.; Araujo, S.G.; Morais, M.I.; Sá, N.P.; Lima, C.M.; Rosa, C.A.; Siqueira, E.P.; Johann, S.; Lima, L.A. Antifungal and antioxidant activity of fatty acid methyl esters from vegetable oils. An. Acad. Bras. Ciências 2017, 89, 1671–1681. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Shen, G.; Tang, J.; Qiu, T.; Zhang, Z.; Zhao, W.; Yu, X.; Huang, J.; Yao, Z. Promotion effect of extracts from plastrum testudinis on alendronate against glucocorticoid-induced osteoporosis in rat spine. Sci. Rep. 2017, 7, 10617. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.Y.; Ren, H.; Huang, J.J.; Zhang, Z.D.; Zhao, W.H.; Yu, X.; Shang, Q.; Qiu, T.; Zhang, Y.Z.; Tang, J.J.; et al. Plastrum Testudinis Extracts Promote BMSC Proliferation and Osteogenic Differentiation by Regulating Let-7f-5p and the TNFR2/PI3K/AKT Signaling Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharm. 2018, 47, 2307–2318. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Poultry; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Iqbal, M.K.; Nabi, F.; Mehmood, K.; Rehman, M.U.; Huang, S.; Zhang, H.; Zhang, L.; Ahmad, H.I.; Iqbal, M.; Li, J. Healing of Growth Plate Cartilage by Hypoxia Inducible Factor-1a Inhibitor Apigenin on Thiram Induced Tibial Dyschondroplasia. Pak. Vet. J. 2018, 38, 143–148. [Google Scholar]
- Liang, D.; Ren, H.; Qiu, T.; Shen, G.; Xie, B.; Wei, Q.; Yao, Z.; Tang, J.; Zhang, Z.; Jiang, X. Extracts from plastrum testudinis reverse glucocorticoid-induced spinal osteoporosis of rats via targeting osteoblastic and osteoclastic markers. Biomed. Pharm. 2016, 82, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, S.; Tong, X.; Zhang, L.; Jiang, X.; Zhang, H.; Mehmood, K.; Li, J. Chlorogenic Acid Alleviates Thiram-Induced Tibial Dyschondroplasia by Modulating Caspases, BECN1 Expression and ECM Degradation. Int. J. Mol. Sci. 2019, 20, 3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, K.; Zhang, H.; Li, K.; Wang, L.; Rehman, M.U.; Nabi, F.; Iqbal, M.K.; Luo, H.; Shahzad, M.; Li, J. Effect of tetramethylpyrazine on tibial dyschondroplasia incidence, tibial angiogenesis, performance and characteristics via HIF-1alpha/VEGF signaling pathway in chickens. Sci. Rep. 2018, 8, 2495. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Wang, Y.; Li, A.; Qamar, H.; Yao, W.; Tong, X.; Zhang, J.; Iqbal, M.; Mehmood, K.; Li, J. Osthole: A Coumarin Derivative Assuage Thiram-Induced Tibial Dyschondroplasia by Regulating BMP-2 and RUNX-2 Expressions in Chickens. Antioxidants 2019, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, S.A.; Sun, L.; Zhang, N.-Y.; Khalil, M.M.; Ling, Z.; Chong, L.; Wang, S.; Rajput, I.R.; Bloch, D.M.; Khan, F.A.; et al. Grape Seed Proanthocyanidin Extract Alleviates AflatoxinB1-Induced Immunotoxicity and Oxidative Stress via Modulation of NF-κB and Nrf2 Signaling Pathways in Broilers. Toxins 2019, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, K.; Zhang, H.; Jiang, X.; Yao, W.; Tong, X.; Iqbal, M.K.; Rehman, M.U.; Iqbal, M.; Waqas, M.; Qamar, H.; et al. Ligustrazine recovers thiram-induced tibial dyschondroplasia in chickens: Involvement of new molecules modulating integrin beta 3. Ecotoxicol. Environ. Saf. 2019, 168, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.; Sun, L.; Zhang, N.; Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.; Zhang, J.; Qi, D.; et al. Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1. Toxins 2018, 10, 366. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Zhang, H.; Mehmood, K.; Li, A.; Jiang, X.; Wang, Y.; Zhang, J.; Iqbal, M.K.; Rehman, M.U.; Yao, W. Icariin: A Potential Compound for the Recovery of Tibial Dyschondroplasia Affected Chicken Via Up-Regulating BMP-2 Expression. Biol. Proced. Online 2018, 20, 15. [Google Scholar] [CrossRef]
- Mehmood, K.; Zhang, H.; Yao, W.; Jiang, X.; Waqas, M.; Li, A.; Wang, Y.; Lei, L.; Zhang, L.; Qamar, H.; et al. Protective effect of Astragaloside IV to inhibit thiram-induced tibial dyschondroplasia. Environ. Sci. Pollut. Res. Int. 2019, 26, 16210–16219. [Google Scholar] [CrossRef]
- Levene, H. Contributions to Probability and Statistics: Essays in Honor of Ingram Olkin; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1960; pp. 278–292. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Van Boxtel, A.L.; Pieterse, B.; Cenijn, P.; Kamstra, J.H.; Brouwer, A.; van Wieringen, W.; de Boer, J.; Legler, J. Dithiocarbamates induce craniofacial abnormalities and downregulate sox9a during zebrafish development. Toxicol. Sci. 2010, 117, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasaputra, K.S.; Liyanage, R.; Lay, J.O., Jr.; Slavik, M.F.; Rath, N.C. Effect of thiram on avian growth plate chondrocytes in culture. J. Toxicol. Sci. 2013, 38, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, K.; Zhang, H.; Iqbal, M.K.; Rehman, M.U.; Shahzad, M.; Li, K.; Huang, S.; Nabi, F.; Zhang, L.; Li, J. In Vitro effect of apigenin and danshen in tibial dyschondroplasia through inhibition of heat-shock protein 90 and vascular endothelial growth factor expressions in avian growth plate cells. Avian Dis. 2017, 61, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.; Gao, J.; Qin, P.; Liu, J.; Wang, Z.; Zhang, D.; Li, J. Expression of genes encoding matrilin-3 and cyclin-I during the impairment and recovery of chicken growth plate in tibial dyschondroplasia. Avian Dis. 2014, 58, 468–473. [Google Scholar] [CrossRef]
- Shahzad, M.; Liu, J.; Gao, J.; Wang, Z.; Zhang, D.; Nabi, F.; Li, K.; Li, J. Differential expression of extracellular matrix metalloproteinase inducer (EMMPRIN/CD147) in avian tibial dyschondroplasia. Avian Pathol. 2015, 44, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Nabi, F.; Iqbal, M.K.; Zhang, H.; Rehman, M.U.; Shahzad, M.; Huang, S.; Han, Z.; Mehmood, K.; Ahmed, N.; Chachar, B.; et al. Clinical efficiency and safety of Hsp90 inhibitor Novobiocin in avian tibial dyschondroplasia. J. Vet. Pharm. 2018, 41, 902–911. [Google Scholar] [CrossRef]
- Velada, I.; Capela-Silva, F.; Reis, F.; Pires, E.; Egas, C.; Rodrigues-Santos, P.; Barros, M.T. Expression of genes encoding extracellular matrix macromolecules and metalloproteinases in avian tibial dyschondroplasia. J. Comp. Pathol. 2011, 145, 174–186. [Google Scholar] [CrossRef]
- Shim, M.Y.; Karnuah, A.B.; Anthony, N.B.; Pesti, G.M.; Aggrey, S.E. The effects of broiler chicken growth rate on valgus, varus, and tibial dyschondroplasia. Poult. Sci. 2012, 91, 62–65. [Google Scholar] [CrossRef]
- Zhang, H.; Mehmood, K.; Jiang, X.; Yao, W.; Iqbal, M.; Li, K.; Tong, X.; Wang, L.; Wang, M.; Zhang, L.; et al. Effect of Icariin on Tibial Dyschondroplasia Incidence and Tibial Characteristics by Regulating P2RX7 in Chickens. Biomed Res. Int. 2018, 2018, 6796271. [Google Scholar] [CrossRef]
- Iqbal, M.K.; Nabi, F.; Rehman, M.U.; Mehmood, K.; Huang, S.; Zhang, H.; Zhang, L.; Iqbal, M.; Li, J. FK228 recovers thiram-induced tibial dyschondroplasia in chicken via hypoxia inducible factor-1alpha. J. Biol. Regul. Homeost. Agents 2018, 32, 89–95. [Google Scholar]
- Genin, O.; Hasdai, A.; Shinder, D.; Pines, M. Hypoxia, hypoxia-inducible factor-1alpha (HIF-1alpha), and heat-shock proteins in tibial dyschondroplasia. Poult. Sci. 2008, 87, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Chen, W.; Zeng, H.P.; Chen, D.F. Chemical components in extracts from Plastrum testudinis with proliferation-promoting effects on rat mesenchymal stem cells. Chem. Biol. Drug Des. 2012, 79, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.P.; Westwood, F.R.; Wadsworth, P.F. Review of the effects of anti-angiogenic compounds on the epiphyseal growth plate. Toxicol. Pathol. 2006, 34, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Algandaby, M.M.; Breikaa, R.M.; Eid, B.G.; Neamatallah, T.A.; Abdel-Naim, A.B.; Ashour, O.M. Icariin protects against thioacetamide-induced liver fibrosis in rats: Implication of anti-angiogenic and anti-autophagic properties. Pharm. Rep. 2017, 69, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Marikovsky, M. Thiram inhibits angiogenesis and slows the development of experimental tumours in mice. Br. J. Cancer 2002, 86, 779–787. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.J.; Shin, D.S.; Getzoff, E.D.; Tainer, J.A. The structural biochemistry of the superoxide dismutases. Biochim. Biophys. Acta 2010, 1804, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, K.; Zhang, H.; Iqbal, M.K.; Rehman, M.U.; Li, K.; Huang, S.; Shahzad, M.; Nabi, F.; Iqbal, M.; Li, J. Tetramethylpyrazine mitigates toxicity and liver oxidative stress in tibial dyschondroplasia chickens. Pak. Vet. J. 2018, 38, 76–80. [Google Scholar] [CrossRef]
- Qian, X.; Chen, J.; Zhao, D.; Guo, L.; Qian, X. Plastrum testudinis induces gamma-globin gene expression through epigenetic histone modifications within the gamma-globin gene promoter via activation of the p38 MAPK signaling pathway. Int. J. Mol. Med. 2013, 31, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Marie, P.J. Signaling pathways affecting skeletal health. Curr. Osteoporos. Rep. 2012, 10, 190–198. [Google Scholar] [CrossRef]
- Chen, J.; Crawford, R.; Chen, C.; Xiao, Y. The key regulatory roles of the PI3K/Akt signaling pathway in the functionalities of mesenchymal stem cells and applications in tissue regeneration. Tissue Eng. Part B Rev. 2013, 19, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Dong, K.; Hao, P.; Xu, S.; Liu, S.; Zhou, W.; Yue, X.; Rausch-Fan, X.; Liu, Z. Alpha-Lipoic Acid Alleviates High-Glucose Suppressed Osteogenic Differentiation of MC3T3-E1 Cells via Antioxidant Effect and PI3K/Akt Signaling Pathway. Cell. Physiol. Biochem. 2017, 42, 1897–1906. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, Q.; Wang, J.; Xi, Q.; Zhu, H.; Gu, M. Gene expression changes in human mesenchymal stem cells from patients with osteoporosis. Mol. Med. Rep. 2015, 12, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, N.; Ma, J.; Chen, X.C.; Li, Z.; Zhao, W. Expression profile analysis of new candidate genes for the therapy of primary osteoporosis. Eur. Rev. Med. Pharm. Sci. 2016, 20, 433–440. [Google Scholar]
- Li, X.J.; Zhu, Z.; Han, S.L.; Zhang, Z.L. Bergapten exerts inhibitory effects on diabetes-related osteoporosis via the regulation of the PI3K/AKT, JNK/MAPK and NF-kappaB signaling pathways in osteoprotegerin knockout mice. Int. J. Mol. Med. 2016, 38, 1661–1672. [Google Scholar] [CrossRef]
- Hogan, B. Bone morphogenetic proteins: Multifunctional regulators of vertebrate development. Genes Dev. 1996, 10, 1580–1594. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, T.; Otsuka, T.; Fujita, K.; Sakai, G.; Matsushima-Nishiwaki, R.; Kozawa, O.; Tokuda, H. (-)-Epigallocatechin gallate but not chlorogenic acid suppresses EGF-stimulated migration of osteoblasts via attenuation of p38 MAPK activity. Int. J. Mol. Med. 2018, 42, 3149–3156. [Google Scholar] [CrossRef] [Green Version]
- Thouverey, C.; Caverzasio, J. Focus on the p38 MAPK signaling pathway in bone development and maintenance. Bonekey Rep. 2015, 4. [Google Scholar] [CrossRef]
- Pathare, G.; Föller, M.; Michael, D.; Walker, B.; Hierlmeier, M.; Mannheim, J.G.; Pichler, B.J.; Lang, F. Enhanced FGF23 serum concentrations and phosphaturia in gene targeted mice expressing WNK-resistant SPAK. Kidney Blood Press. Res. 2012, 36, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Xuan, S.; Bouxsein, M.L.; Von Stechow, D.; Akeno, N.; Faugere, M.C.; Malluche, H.; Zhao, G.; Rosen, C.J.; Efstratiadis, A. Osteoblast-specific knockout of the insulin-like growth factor (IGF) receptor gene reveals an essential role of IGF signaling in bone matrix mineralization. J. Biol. Chem. 2002, 277, 44005–44012. [Google Scholar] [CrossRef] [Green Version]
- Shang, Q.; Yu, X.; Ren, H.; Shen, G.; Zhao, W.; Zhang, Z.; Huang, J.; Yu, P.; Liang, D.; Tang, J. Effect of Plastrum Testudinis Extracts on the Proliferation and Osteogenic Differentiation of rBMSCs by Regulating p38 MAPK-Related Genes. Evid. Based Complement. Altern. Med. 2019, 2019, 6815620. [Google Scholar] [CrossRef]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. Regulation of the T cell response. Clin. Exp. Allergy 2006, 36, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Vanamee, E.S.; Faustman, D.L. TNFR2: A Novel Target for Cancer Immunotherapy. Trends Mol. Med. 2017, 23, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Faustman, D.; Davis, M. TNF receptor 2 pathway: Drug target for autoimmune diseases. Nat. Rev. Drug Discov. 2010, 9, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Primer Sequence (5′-3′) | Product Size (bp) | Tm (°C) |
|---|---|---|---|---|
| AKT | AF039943 | F: TGATGGCACATTCATTGGCTAC R: TGTTTGGTTTAGGTCGTTCTGTCT | 122 | 58 |
| PI3K | NM001004410 | F: CGGATGTTGCCTTACGGTTGT R: GTTCTTGTCCTTGAGCCACTGAT | 162 | 58 |
| GAPDH | XM_019960295 | F: CCTTCATTGACCTTCACTACATGGTCTA R: TGGAAGATGGTGATGGCCTTTCCATTG | 127 | 58 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qamar, H.; Waqas, M.; Li, A.; Iqbal, M.; Mehmood, K.; Li, J. Plastrum Testudinis Extract Mitigates Thiram Toxicity in Broilers via Regulating PI3K/AKT Signaling. Biomolecules 2019, 9, 784. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120784
Qamar H, Waqas M, Li A, Iqbal M, Mehmood K, Li J. Plastrum Testudinis Extract Mitigates Thiram Toxicity in Broilers via Regulating PI3K/AKT Signaling. Biomolecules. 2019; 9(12):784. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120784
Chicago/Turabian StyleQamar, Hammad, Muhammad Waqas, Aoyun Li, Mudassar Iqbal, Khalid Mehmood, and Jiakui Li. 2019. "Plastrum Testudinis Extract Mitigates Thiram Toxicity in Broilers via Regulating PI3K/AKT Signaling" Biomolecules 9, no. 12: 784. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120784