Bioactivities of Halometabolites from Marine Actinobacteria

1
Integrated Laboratory, Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta 55281, Indonesia
2
Department of Fisheries, Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta 55281, Indonesia
*
Author to whom correspondence should be addressed.
Biomolecules 2019, 9(6), 225; https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060225
Submission received: 5 April 2019
/
Revised: 27 May 2019
/
Accepted: 28 May 2019
/
Published: 11 June 2019
(This article belongs to the Special Issue Bioactives from Marine Products)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Natural halogenated compounds (halometabolites) are produced mainly by marine organisms, including marine Actinobacteria. Many commercially important compounds for pharmaceuticals contain halogen, and the halogen is responsible for the physical and chemical properties as well as bioactivities and toxicities. In the exploration of marine environment that is supported by advanced structure elucidation, varied panel bioassays and high-throughput screening have accelerated number of halometabolites isolated from marine Actinobacteria to date. The metabolites exhibited unique structures and promising bioactivities. This review focuses on the chemodiversity and bioactivities of marine halometabolites from marine Actinobacteria reported in the last 15 years (2003–2018).
1. Introduction
Halometabolites are a group of compounds contain halogen substituents (F, Cl, Br, I). To date, there are more than 5000 halogenated compounds with high degree of structural variability containing a single or several halogen atoms [1]. Halogen elements are found in several forms in nature. Chloride, iodine, and bromide salts are present in the oceans, while the Earth’s crust is rich in fluorine. Natural organohalogens (or halometabolites) are produced from two sources: abiogenic and biogenic. Natural abiogenic organohalogen is formed during geothermal processes such as volcano, hot springs, or earthquake. Biomass burning and soil chemistry have also contributed to the enormous number of abiogenic halometabolites [2]. Biogenic halometabolites are produced by bacteria, fungi, plants, marine invertebrates, and macroalgae [1,2,3,4,5,6,7,8].
Halometabolites in nature have several functions in physiological, biochemical, or defensive role for their host including communication (quorum sensing) and production of growth hormones, sex pheromone, toxin, or antibiotics. The role of substituent halogen in organic compounds is related to the bioactivity, bioavailability, and stability of the compounds.
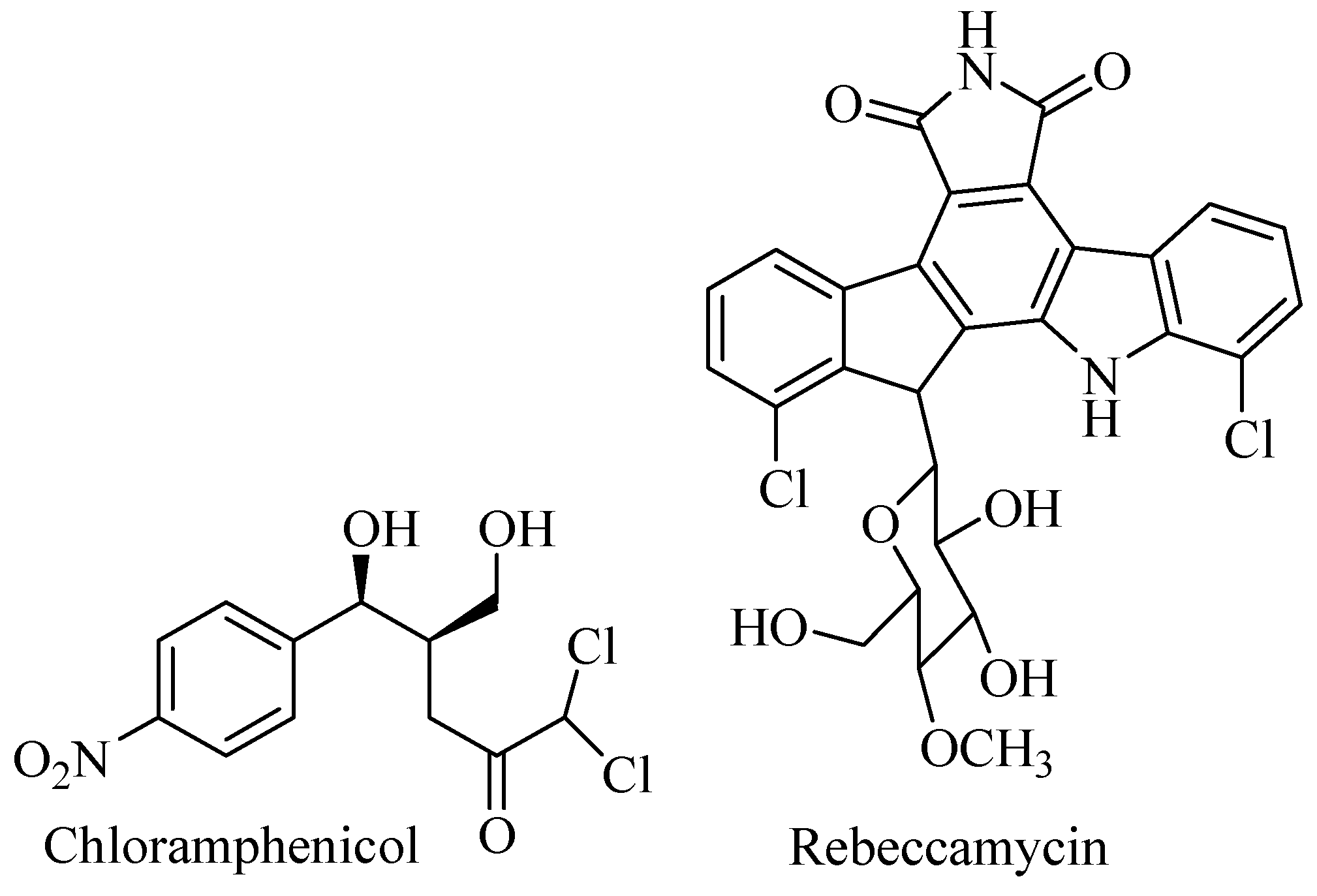
Chlorinated antibiotics were discovered from the exploration of soil Actinobacteria since the discovery of streptomycin from Streptomyces griseus. A number of drugs derived from Actinobacteria such as antibiotic and anticancer are on the market today. Chlorinated antibiotics such as chloramphenicol and vancomycin played important roles for the eradication of infectious diseases in human. Chloramphenicol is a broad-spectrum antibiotic used to treat bacterial infections. Chloramphenicol antibiotic is on the WHO (World Health Organization) list of essential medicine. Chlortetracycline is a member of the tetracycline family and produced by Streptomyces aureofaciens. Chlortetracycline was used clinically in 1948 and is used to prevent, control, and treat animal health problems and increase growth rate in chickens, turkeys, ducks, swine, calves, beef cattle, and others. Calicheamicin is a group of enediyne metabolite with iodine and has remarkable activity as anticancer produced by Micromonospora echinospora. Linking calicheamicin to monoclonal antibody is used for therapy of acute myeloid leukemia. Rebeccamycin is produced by Streptomyces sp. and is a derivate of staurosporine with chlor attached to it. Rebeccamycin showed in vitro antitumor activity at Inhibitory Concentration (IC50) 480 nM against P388 leukemia cells. Complestatin is a cyclic halogenated peptide produced by Streptomyces lavendulae anti and exhibited activity as HIV-1 integrase inhibitor. Vancomycin is a halogenated glycopeptide active against Staphylococcus aureus (including methicillin-resistant strains), S. epidermidis (including multiple-resistant strains), Streptococcus pneumoniae (including multiple-resistant strains), S. pyogenes, S. agalactiae, S. bovis, S. mutans, Clostridium spp., Listeria monocytogenes, Actinomyces spp., and Lactobacillus spp. [9,10,11].
2. Research Methodology
In the studies on bioactive marine natural products, we look for the potential of marine Actinobacteria as halometabolites producers. This comprehensive review illustrates the chemistry and biological activities of halometabolites produced by marine Actinobacteria reported in the last 15 years (2003–2018). Mining and searching for data of compounds and bioactivities were obtained from reports in the database MarinLit, Google Scholar, ScienceDirect, Dictionary of Marine Natural Products, and Marine Natural Product Review. Herein, we grouped the halometabolites based on class of compounds.
3. Halometabolites Isolated from Marine Actinobacteria
The marine environment is home for wide diversity of organisms and sources of structurally diverse secondary metabolites and drug leads. Halometabolites were produced mainly by marine organisms because seawater contained ion chloride and bromine. Marine organisms have the capability to oxidize bromide more easily than chlorine in the biosynthesis of organic compounds, thus bromometabolites are higher than chlorometabolites as observed in sponge and red algae [4,5,7].
The phylum Actinobacteria is Gram-positive bacteria with high G-C content in DNA. Terrestrial Actinobacteria has been explored for decades as sources of pharmacologically active compounds, and more than 70% of antibiotics used today are derived from Actinobacteria. Other bioactive compounds such as anticancer, antifungal, anthelminthic, antidiabetic, etc., were discovered from terrestrial Actinobacteria as well. Figure 1 shows diverse commercial halometabolites isolated from terrestrial Actinobacteria to prove Actinobacteria play important roles for biomedicine and biotechnology applications [9,12].
Marine environment is different from terrestrial so that marine Actinobacteria have special characteristic and adapted to stress in marine environment. As a result, marine Actinobacteria produce new type of secondary metabolites that differs from terrestrial one. Marine Actinobacteria can be found in any part of the ocean such as water column, sediment, deep sea, and in association with seaweed, sponges, and marine organisms [13,14,15]. Marine Actinobacteria have been explored and yielded structurally unique secondary metabolites with varied biological activities [15,16,17]. Searching for bioactive halometabolites was focused on many members of this genera such as Streptomycetes, Actinoplanes, Nocardia, and other rare Actinobacteria [18,19,20]. One study showed that marine Actinomycetes was a major producer of antibacterial compounds compared to Bacilli and Gammaproteobacteria [21].
3.1. Alkaloid
3.1.1. Lynamicins
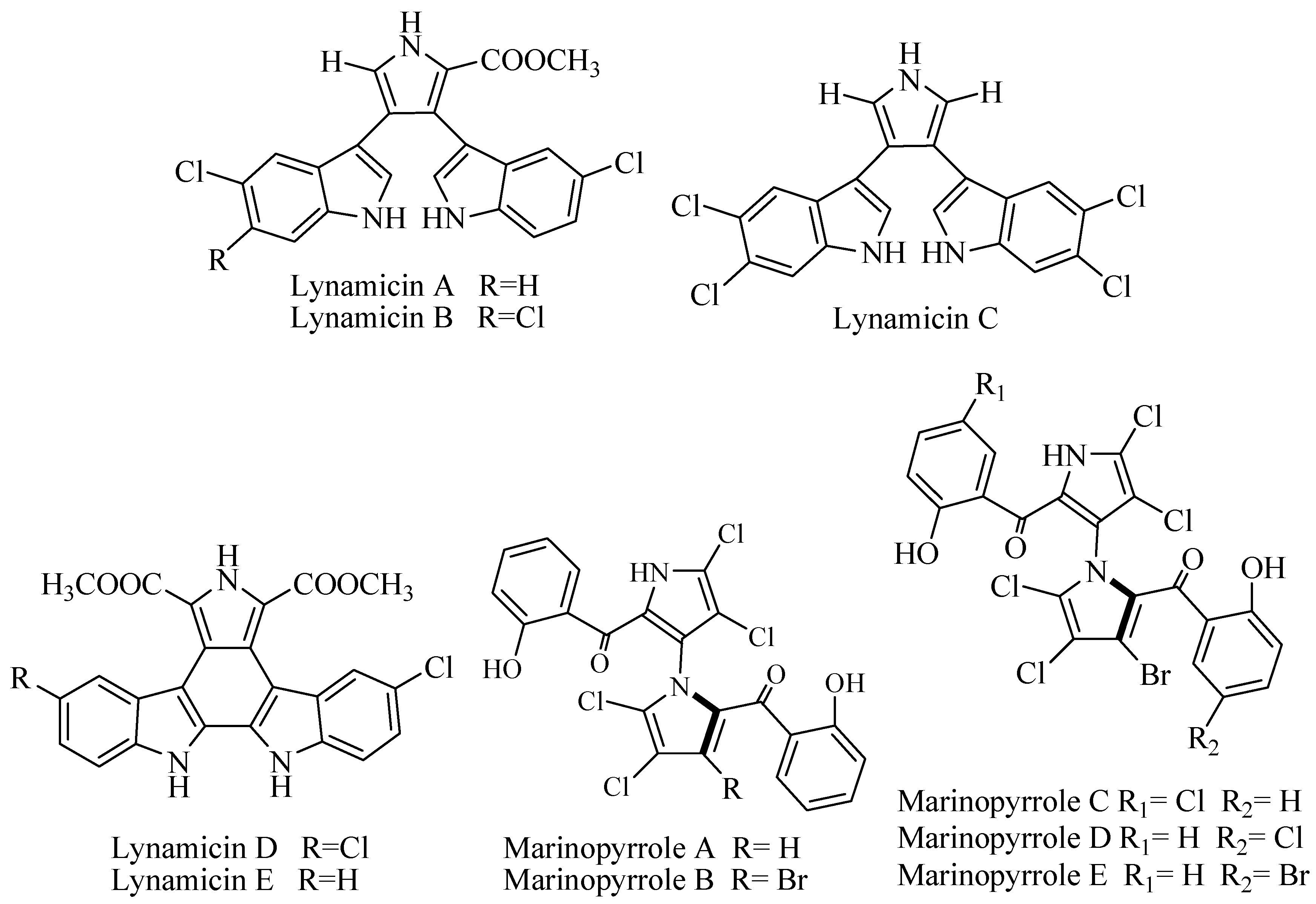
Marine Actinomycete NPS 12745 (Marinispora sp.) was isolated from marine sediment of Mission Bay, San Diego, coast yielded bisindole pyrrole compounds named lynamicins A–E (Figure 2). The series of compounds were tested against series of panel test bacteria that were resistant and sensitive to antibiotics. However, lynamicin E exhibited broad-spectrum activity and potency for treatment of nosocomial infection at Minimum Inhibition Concentration (MIC) 1.8–36 μg/mL [22].
3.1.2. Marinopyrroles
An obligate marine Actinomycetes CNQ-418 related to Streptomyces was isolated from 51 m sediment of La Jolla, California, produced two unique halogenated metabolites with uncommon 1,3′-bipyrrole pharmacophore called marinopyrrole A and B (Figure 2). The compounds were active against methicillin-resistant Staphylococcus aureus at MIC90 0.61 and 1.1 μM for marinopyrrole A and B, respectively. The IC50 against HCT-16 (human colon cancer cell line) for marinopyrrole A was 8.8 μM and marinopyrrole B was 9.0 μM [23]. Further examination of broth fermentation of Streptomyces strain CNQ-418 yielded marinopyrroles A–F. The compounds showed variation of substitution chlorine and bromine. Marinopyrrole A–C had significant activity against MRSA (methicillin-resistant Staphylococcus aureus) with MIC90 at less than 1 μg/mL [24]. Marinopyrrole A showed activity against S. aureus strains with MIC 0.188–1.5 μg/mL. This activity was better than available antibiotics vancomycin and linezolid. In addition, marinopyrrole A was active against H. influenzae at MIC 2 μg/mL. The toxicity against mammalian cell line was more than 20 times of the MIC value [25]. Marinopyrrole A is reported to be an antagonist of Myleoid Leukemia (Mcl-1), a member of the anti-apoptotic B-cell Lymphoma-2 (Bcl-2) family, which is a well-validated drug target for cancer treatment. The cell-based assay shows a high selectivity of marinopyrrole A. Treatment with marinopyrrole A inhibits the viability of K562 cells transfected with Mcl-1 gene with Effective concentration (EC50) value of 1.6 μM. The selectivity is more than 40-fold greater over the cells transfected with Bcl-XL gene. Moreover, marinopyrrole A can decrease Mcl-1 expression by increasing the cleavage of caspase-3 and Poly (ADP-ribose) polymerase (PARP). Marinopyrrole A is also reported to completely restore the sensitivity of multidrug-resistant leukemia cells to ABT-737 [26].
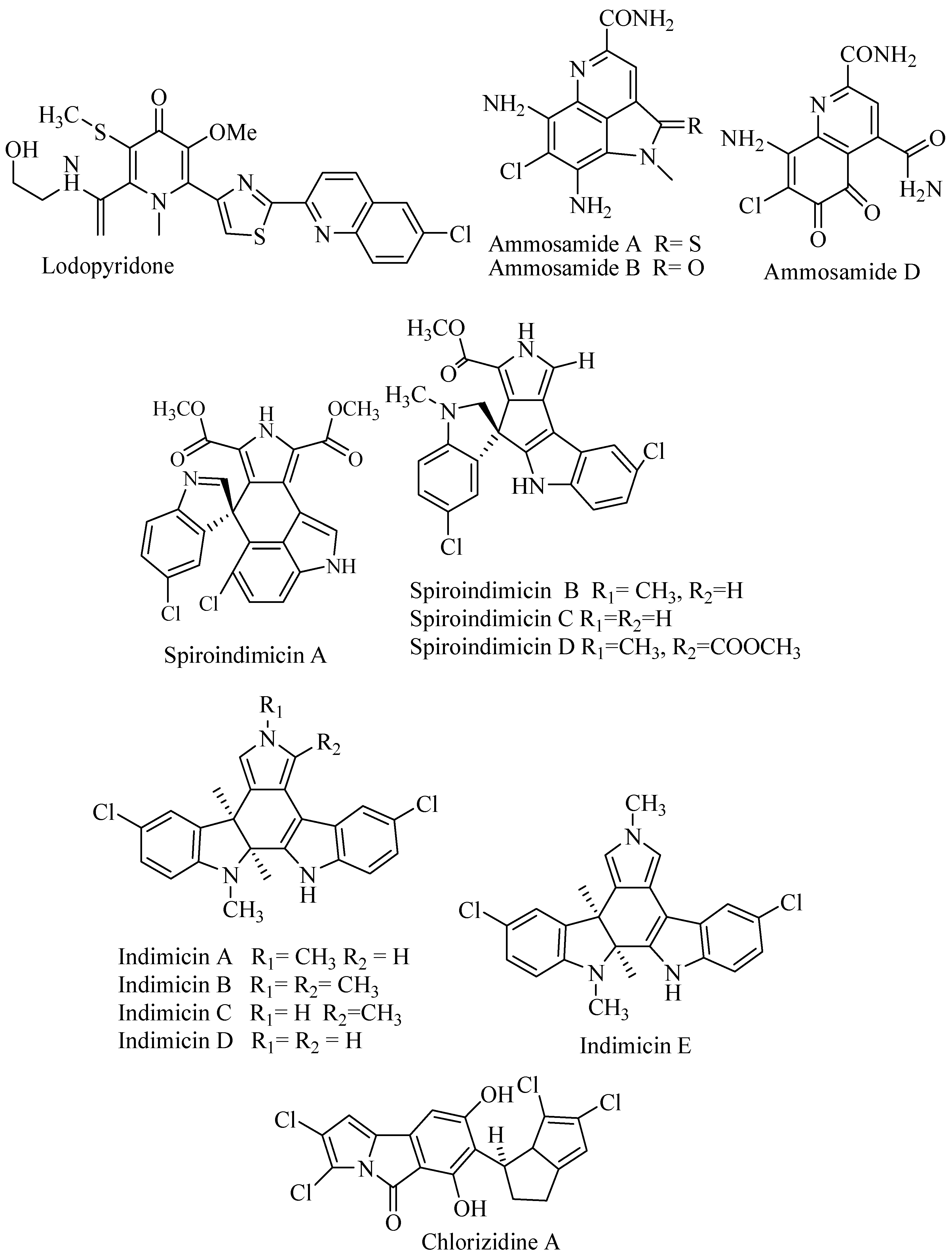
3.1.3. Lodopyridone
Lodopyridone (Figure 2) is a unique alkaloid isolated from an obligate marine Saccharomonospora CNQ-490 collected from sediment in La Jolla Submarine Canyon, California. The compound has interesting carbon skeleton properties with ethanolamine, thiomethyl with substitution of pyridine, thiazole, and chloroquinoline. The bioactivity was modest against cancer cell line HCT-116 at IC50 3.6 μM, but there was no activity against MRSA [27].
3.1.4. Ammosamide
Heteroaromatic alkaloids contain pyrroloquinoline ammosamide A and B (Figure 2) were isolated from Streptomyces CNR-698 from Bahama Island at 1618 m. Bioassay-guided fractionation yielded ammosamide A and B that showed cytotoxicity against HCT-116 at IC50 320 nM [28]. Ammosamide D was isolated from marine Streptomyces variabilis SNA-020 from Sweetings Cay, Bahamas. The cytotoxicity of ammosamide D to human cell line HCT-116 was at IC50 3.2–4.9 µM [29].
3.1.5. Spiroindimicins
PCR-based screening of tryptophan dimerization gene has enabled to select the deep-sea Streptomyces SCS10 03032 from South China Sea produced bisindole alkaloid with unique spiro ring spiroindimicin A–D (Figure 2) along with lynamicin A and D. The compounds were evaluated for cytotoxic activity against cancer cell lines MCF-7, HepG2, B16, H460, and CCRF-CEM. Spiroindimicin A showed no inhibitory activity, while spiroindimicin B showed cytotoxicity against CCRF-CEM (IC50 4 μg/mL), B16 (IC50 5 μg/mL), and H460 (IC50 12 μg/mL). Spiroindimicin C exhibited inhibition against HepG2 and H460 at IC50 6 and 15 μg/mL respectively, while spiroindimicin B and D with property [5,5]spiro-ring showed moderate activity against HepG2, B16, and H460 [30].
3.1.6. Indimicin
Deep sea Streptomyces sp. SCS10 03032 from South China produced chlorinated bisindole alkaloid indimicin A–E (Figure 2) along with lynamicin F and G. The antibacterial activity was tested against E. coli ATCC 25922, S. aureus ATCC 29213, B. thuringiensis SCSIO BT01, B. subtilis SCSIO BS01, and C. albicans ATCC10231 with MIC > 128 μg/mL and was considered as inactive. The cytotoxicity was tested against cancer cell line, and only indimicin B was active against MCF-7 cell line at IC50 10 μM [31].
3.1.7. Chlorizidine
An obligate marine Streptomyces strain CNH-287 was isolated from marine sediment San Clemente, California, produced chlorizidine (Figure 2). The compound revealed feature structure on 5H-pyrrolo[2,1-a]isoindol-5-one ring system that never been reported before. The cytotoxicity against HCT-116 was reported at IC50 3.2–4.9 μM [32].
3.2. Terpene and Meroterpene
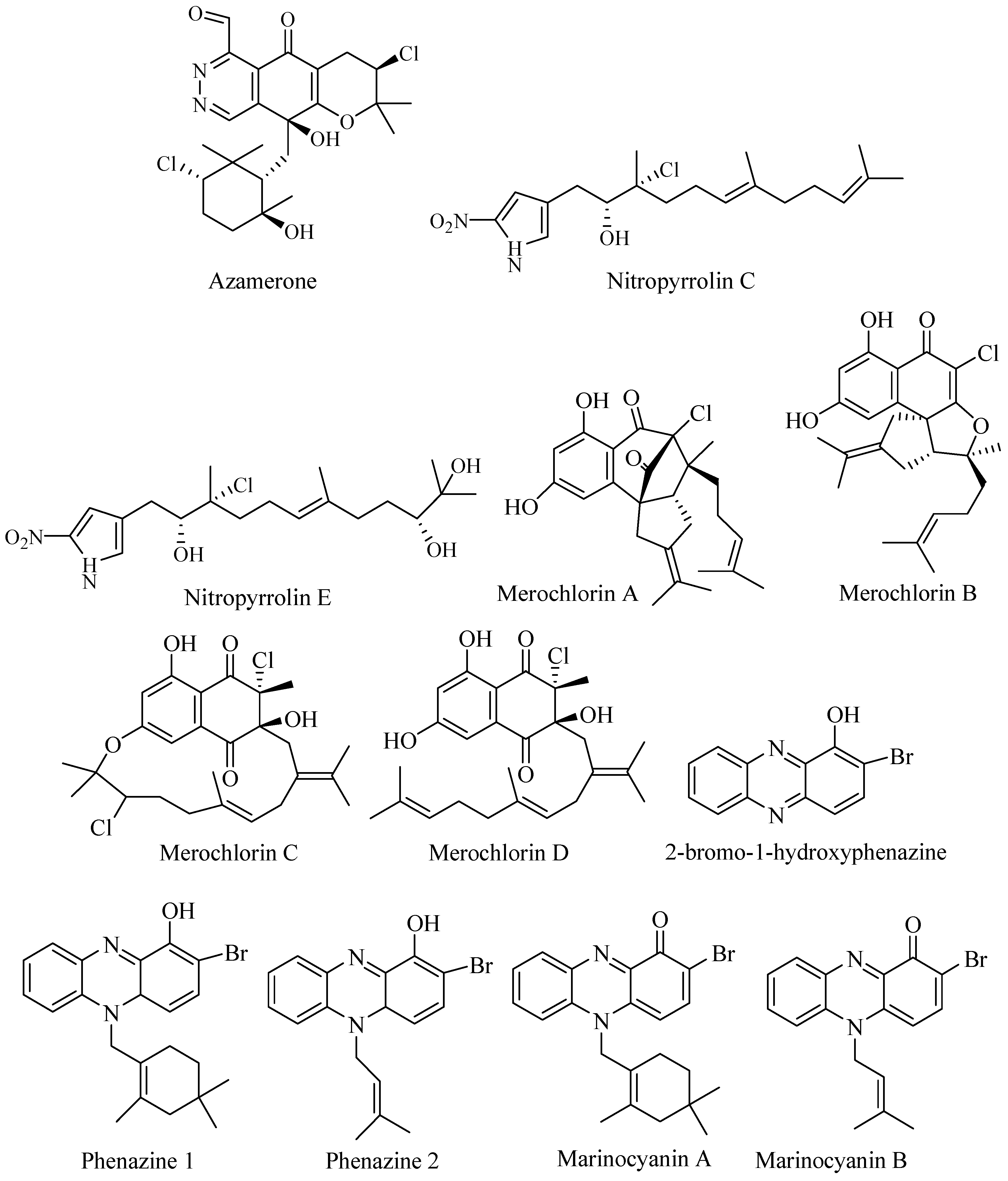
3.2.1. Azamerone
Azamerone (Figure 3) was isolated from saline culture of two strain Streptomyces MAR-4 and CNQ-766. The compound is meroterpenoid with phthalazinone ring with a side chain of 3-chloro-6-hydroxy-2,2,6-trimethyl-cyclohexyl-methyl (Figure 3). In vitro cytotoxicity assay against mouse splenocyte population of T-cells and macrophage showed weak potency at IC50 40 μM [33].
3.2.2. Nitropyrrolins
Marine Actinomycetes strain CNQ-509 isolated from marine sediment of La Jolla, California, produced a new set of nitrophyrrolins (Figure 3). The compounds were hybrid isoprenoid composed of linear sesquiterpenoid and α farnesyl nitropyrrole. Two compounds had chlorin moieties Nitropyrolins C and E. The compounds showed no activity against MRSA and cancer cell line HCT-116 [34].
3.2.3. Merochlorins
Bioassay-guided fractionation has led to discover novel meroterpen merochlorins (Figure 3) from marine Streptomyces CNH-189 from coastal sediment in California. The compounds displayed unrelated skeleton to available antibacterial agents [35]. Merochlorin A was active against Gram-positive bacteria but inactive against Gram-negative bacteria and showed no cross-resistance to Gram-positive bacteria. Merochlorins A was active against MRSA, MSSA (methicillin-sensitive Staphylococcus aureus), VSSA (vancomycin-sensitive Staphylococcus aureus), and VRSA (vancomycin- resistance Staphylococcus aureus) at concentration 2–4 μg/mL, and merochlorin A was active against Clostridium difficile [36].
3.2.4. Terpenoid Phenazines
Bioassay-guided fractionation of fermentation broth of Streptomyces sp. CNS-284 and CNY-90 led to isolation of brominated terpenoid phenazine 2-bromo-1-hydroxyphenazine and two novel brominated phenazines (marinocyanin A and B) along with lavanducyanin (nonbrominated phenazine) (Figure 3). Strain CNS-284 was isolated from Palau and strain CNY-90 was isolated from the Solomon Islands. The brominated compounds were active as anti-inflammatory agents and inhibited Tumor Necrosis Factor-α (TNF-α) induced Nuclear factor-kB (NF-kB at IC50 4.1, 24.2, and 16.3 µM, respectively. The compounds also showed activity against Lipopolysaccharide (LPS)-induced NO production at IC50 > 48.6, 15.1, and 8.0 μM, respectively. The production Protaglandin E2 (PGE2) was blocked at IC50 7.5, 0.89, and 0.63 μM, respectively. In addition, 2-bromo-1-hydroxyphenazine showed activity in NF-kB-luciferase assay at IC50 73 μM [37].
Marinocyanin A–F were identified from Streptomyces CNY-90. Marinocyanin A had potency as antifungal against amphotericin-resistant Candida albicans with MIC 0.95 μM. Marinocyanin A and B showed inhibition against cancer cell line HCT-116 at IC50 0.049 μM and 0.029 μM, respectively [38].
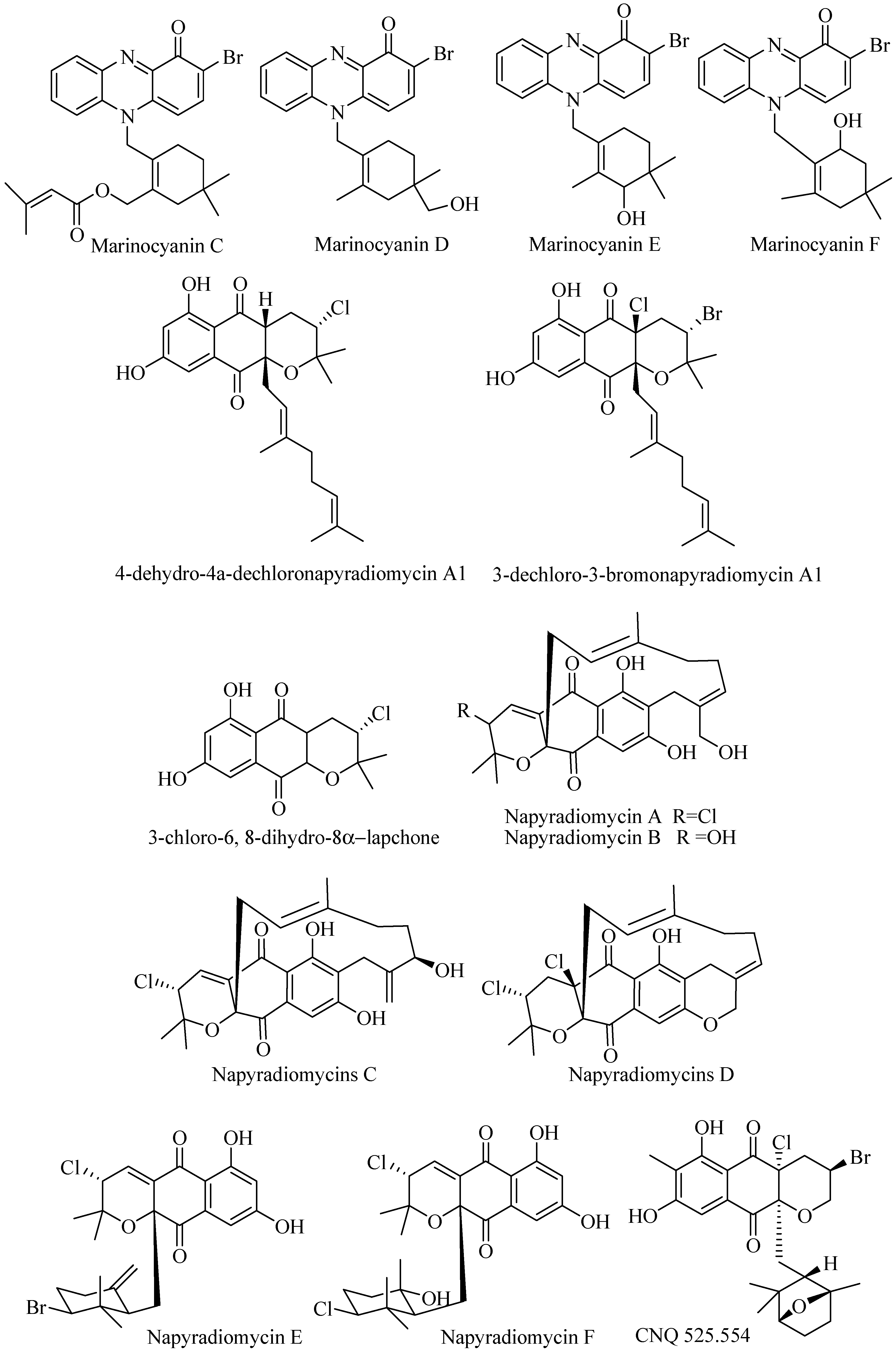
3.2.5. Napyradiomycins and Related Compounds
Napyradiomycins are the group of hybrid terpene and polyketide compounds that consist of napthoquinone ring system with halogen substitution. Investigation of broth culture of marine-derived Streptomyces SCSIO 10428 from Xieyang Island, China, yielded three new napyradiomycins- related compounds (Figure 3) 4-dehydro-4a-dechloronapyradiomycin A1, 3-dechloro-3-bromo napyradiomycin A1, and 3-chloro-6,8-dihydroxy-8-α-lapachone, isolated along with main products napyradiomycin A1, 18-oxonapyradiomycin A1, napyradiomycin B1, napyradiomycin B3, naphthomevalin, and napyradiomycin SR. Evaluation of their bioactivities showed that they had antibacterial activities with MIC values ranging from 0.5 to 32 μg/mL against Staphylococcus aureus ATCC 29213, Bacillus subtilis SCSIO BS01, and Bacillus thuringiensis SCSIO BT01 but no activity against Gram-negative bacteria. Some displayed activity to human cancer cell line SF-268, MCF-7, NCI-H460, and HepG-2 with IC50 values below 20 μM, but some showed activity above 20 μM [39].
Six novel napyradiomycins A–F (Figure 3) along with napyradiomycins B2–B4 were isolated from Streptomyces strain CNQ-329 and CNH-070 from sediment in San Diego, California. The strains hade similarity to Streptomyces aculeolatus. The napyradiomycins were evaluated against MRSA and HCT-116. Napyradiomycin A was active against MRSA at MIC 16 μg/mL while napyradiomycin B-F were inactive. Napyradiomycin B3 was the most active at MIC 2 μg/mL. Napyradiomycins inhibited HCT-116 at IC50 range 4.19–16 μg/mL [40].
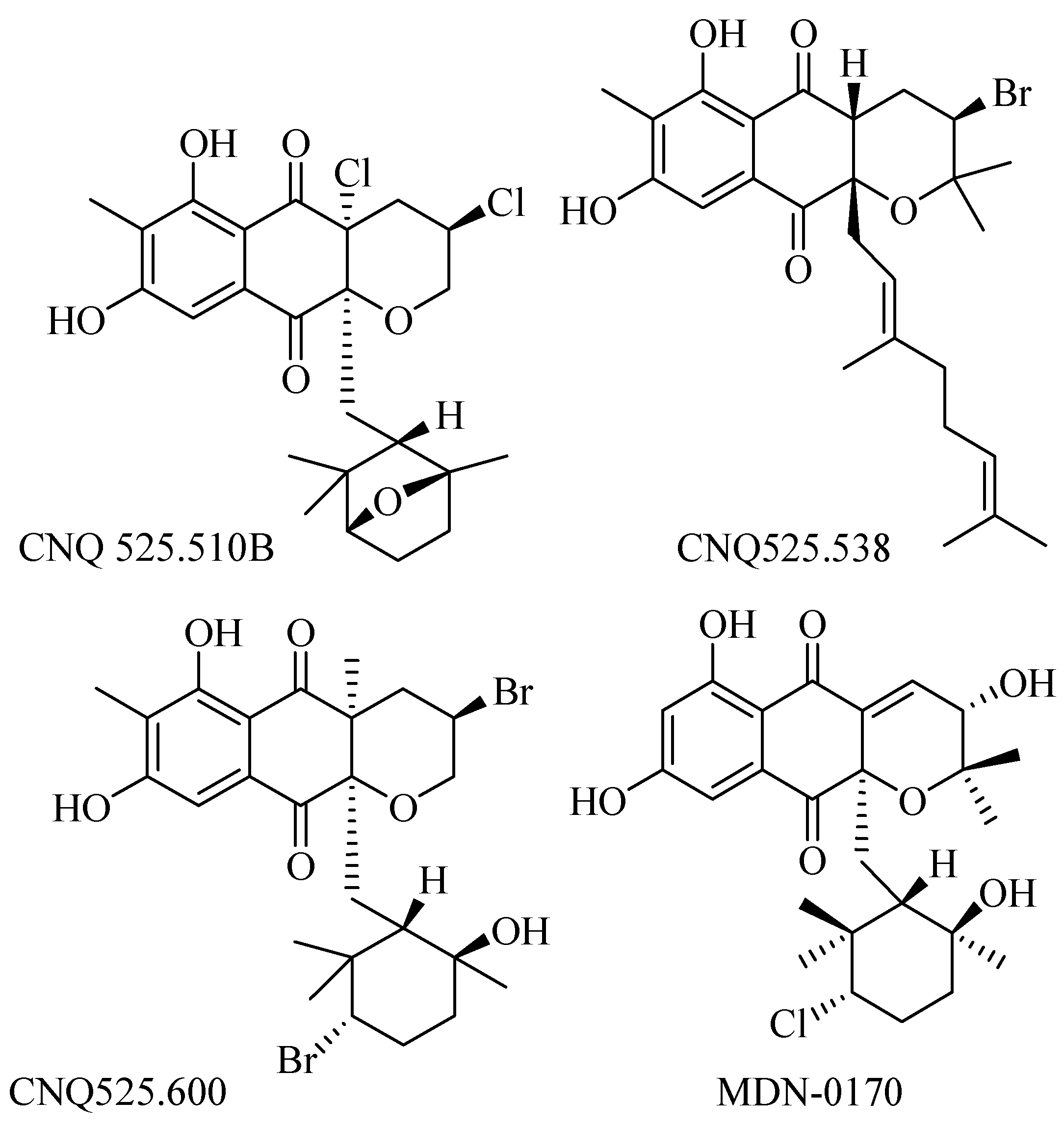
Strain Streptomyces CNQ-525 produced varied types of napyradiomycin that have been reported. Further investigation of the strain has yielded four new napyradiomycin called CNQ525.510B, CNQ525.538, CNQ525.554, and CNQ525.600 (Figure 3) along with known napyradiomycins such as B1, B3, B4, A80915A, A80915B, A80915C, A80915D, CNQ525.512, and SF2415B3. The compounds were tested against HCT-116 colon carcinoma and showed activity at range less than 1 μM to more than 100 μM [41].
3.3. Peptides
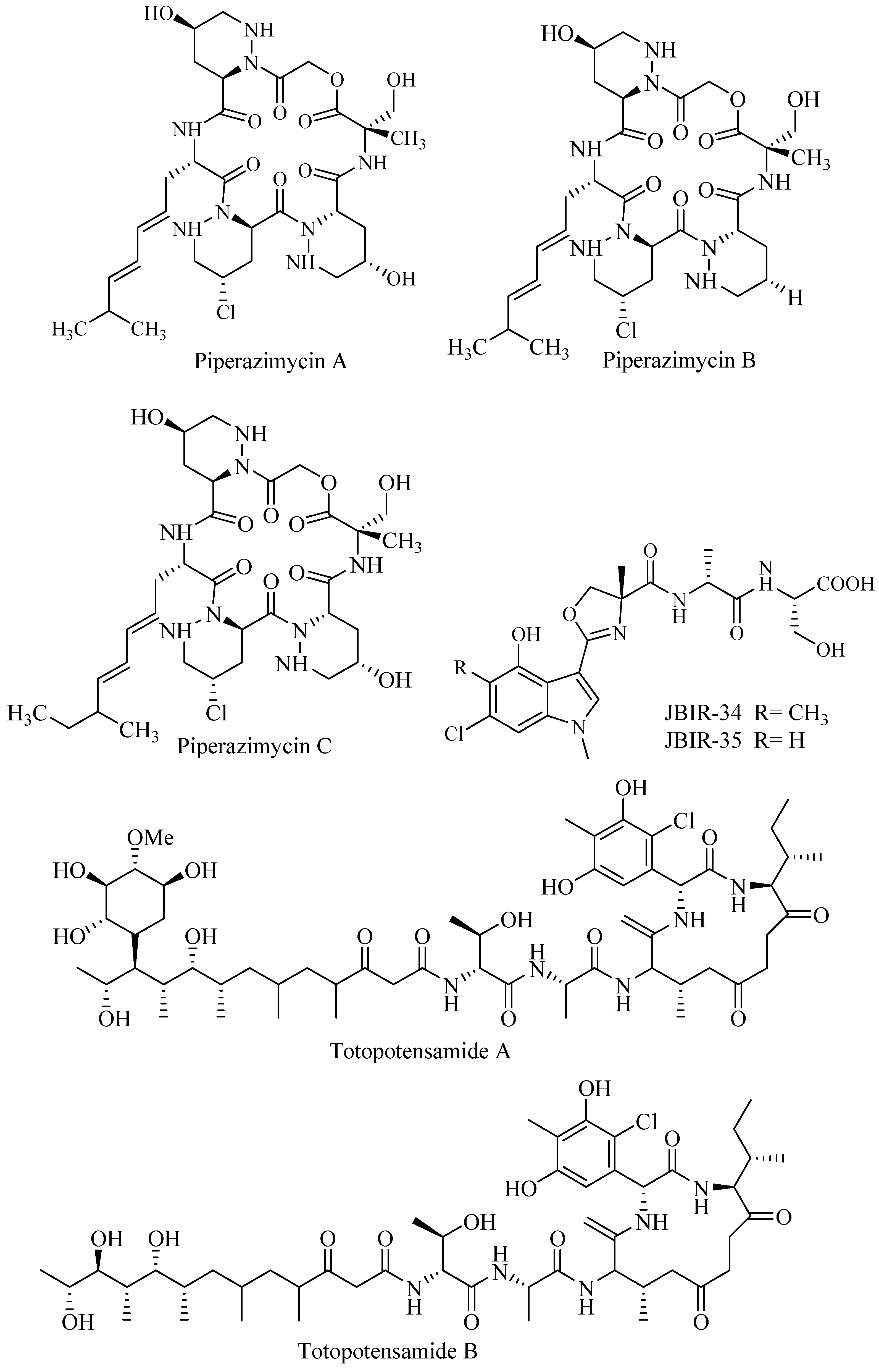
3.3.1. Piperazimycins
Cyclic chlorinated hexadepsipeptide piperazimycin A–C (Figure 4) were isolated and purified from an ethyl acetate extract of culture fermentation of Streptomyces sp. isolated from Guam. The compounds were assayed for bioactivity against human colon carcinoma and 60 cancer cell lines. Each compound exhibited significant cytotoxicity with an average GI (Growth Inhibition)50 76 ng/mL against HCT-16 (human colon carcinoma). Piperazimycin A was the most potent and 3 times more active against solid tumor compared to other piperazimycins [43].
3.3.2. JBIR
3.3.3. Totopotensamides
Streptomyces sp. 1053 U.I. Ia.Ib cultivated from gastropod Lienardia totopotens collected near Mactan Island, Cebu, Philippines, produced hybrid peptide-polyketide glycoside totopotensamide A and B (Figure 4). The compounds had interesting features but showed no activity in wide-range bioassay including DRG panel assay for neurological activity [45].
3.4. Polyketides
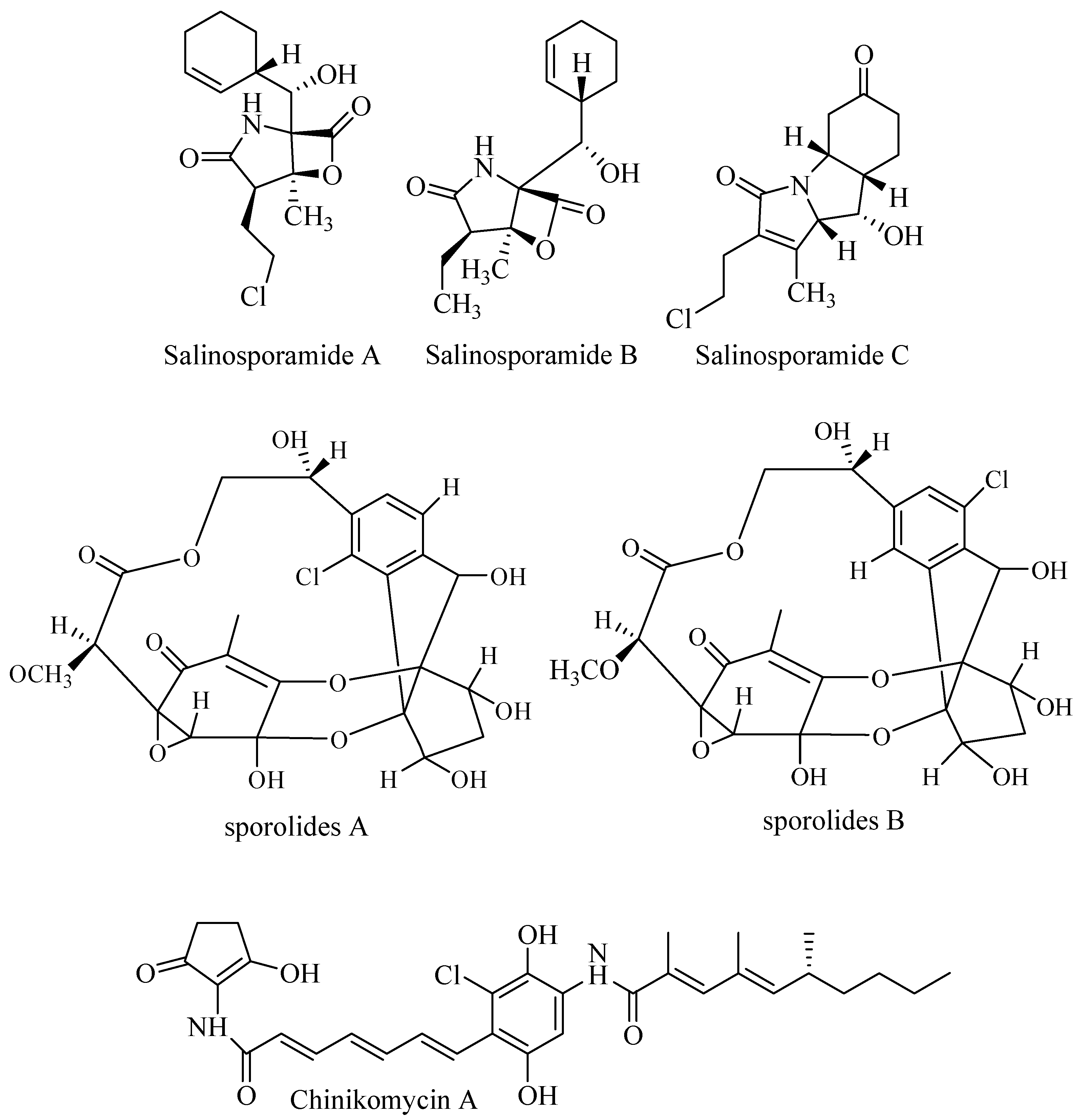
3.4.1. Salinosporamides
Salinosporamide A (Figure 5) was discovered from culture broth of Salinispora tropica CNB. The compound has unique and unusual structure consists of fused γ-lactam β-lactone ring structure. Salinosporamide A and B inhibited selectively the proteolytic activity of the 20S subunit of the proteasome. Both compounds also inhibited human colon carcinoma HCT-116 but had no activity against antibiotic-resistant strain Staphylococcus aureus, Enterococcus faecium, Candida albicans, and herpes simplex virus. Salinosporamide A inhibited proteasomal chymotrypsin-like proteolytic at IC50 1.3 nM. Cytotoxicity of salinosporamide A was observed against HCT-116 at IC50 11 ng/mL. The strong potency was examined against NCI-H226 (non-small cell lung cancer), SF-539 (CNS cancer), SK-MEL-28 (melanoma), and MDA-MB-435 (breast cancer) LC50 less than 10 nM. Salinosporamide A shares structure similarity to omuralide A but is more potent than omuralide A. This is due to methylation at C-3, chloroethyl group at C-2, and cyclohexene at C-5. Β-lactone moiety is the key for bioactivity. Mechanism of action of salinosporamide A induces apoptosis, suppresses osteoclastogenesis, and inhibits invasion through down-modulation of NF-ĸB regulated gene products [46,47].
3.4.2. Sporolides
Detailed examination of fermentation broth of Salinispora tropica CNB-392 (salinosporamide producer) turn out to discover unique polyketide sporolides (Figure 5). These interesting chemical structures were inactive when tested in assay against cancer cell line HCT-116, bacteria MRSA, and Vancomycin Resistance Enterococcus (VRE) [48].
3.4.3. Chinikomycins
Two novel antitumor antibiotic chinikomycins A and B (Figure 5) were isolated from marine Streptomyces griseoauranticus M045 along with manumycin A. Chinikomycins were inactive in antibacterial, antiviral, and phytotoxicity assays. Chinikomycin A showed antitumor activity against cell lines MAXF401 NL (mammary), MEXF462 NL (melanoma), and REX (renal cancer) at IC50 2.41, 4.15, and 4.02 μg/mL, respectively. Chinikomycin B was active against MAXF401 at IC50 3.04 μg/mL [49].
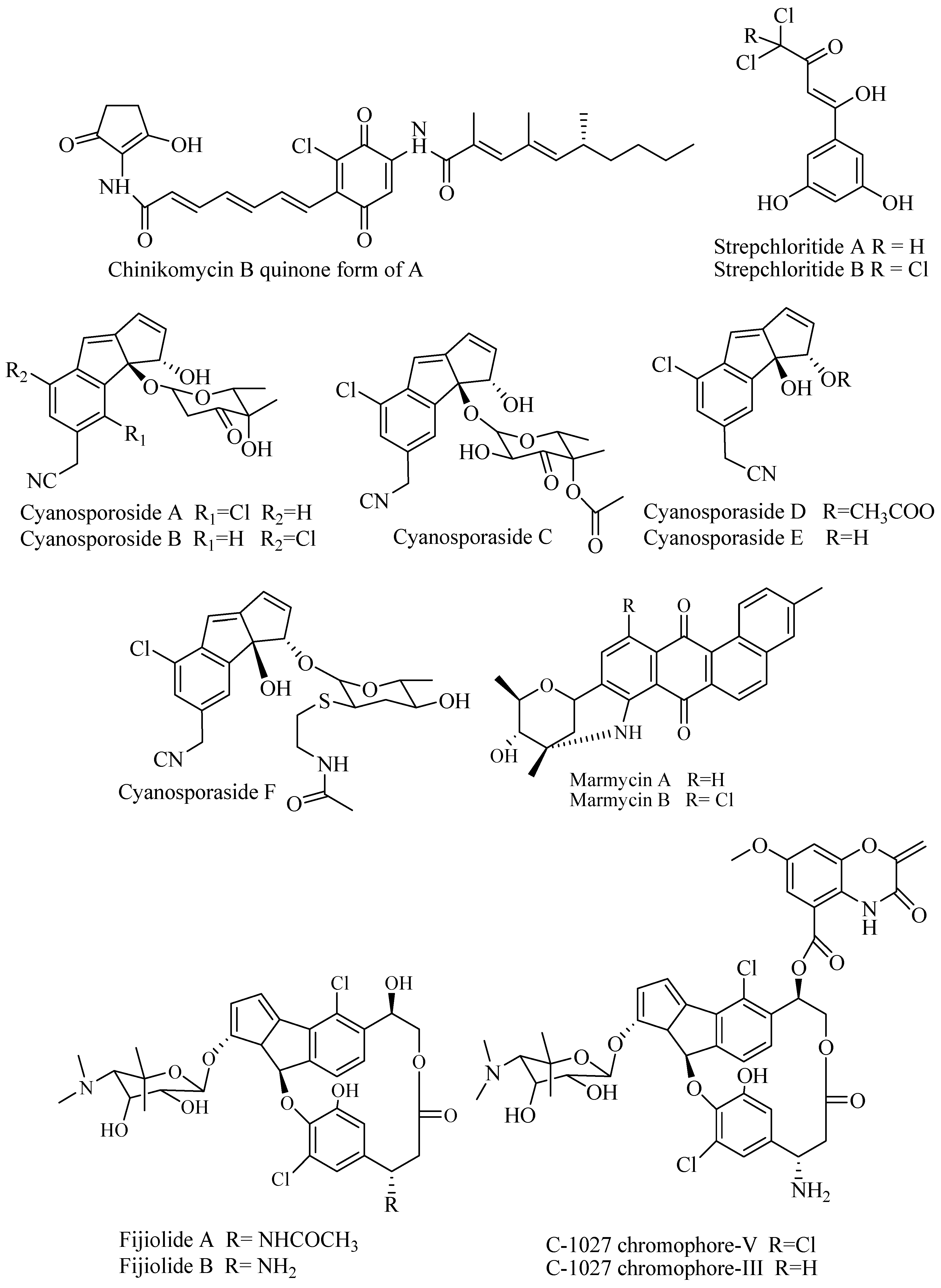
3.4.4. Cyanosporasides
Two novel compounds cyanosporaside A and B (Figure 5) were isolated from Salinispora pacifica CNS 103 collected from deep-sea sediment in Palau. The cyanosporasides contain 3-keto-pyranohexose sugar and a cyano- and chloro-substituted cyclopenta [a] indene ring. In the bioassay against resistant strain, cyanosporoside A was inactive as antibacterial agent against MRSA, VREF (vancomycin-resistance Enterococcus faecalis), and amphotericin-resistant Candida albicans (ARCA). The activity against HCT-116 was weak at IC50 30 μg/mL [50]. New derivates (cyanosporasides C–F) were isolated from the marine Actinomycetes Salinispora pacifica CNS-143 and Streptomyces sp. CNT-179 [51].
3.4.5. Marmycins
Marmycins, an angucycline class of compounds, were isolated from marine Actinomycetes belonging to Streptomyces CNH-990. The compounds have no significant activity as antibiotic against MRSA and VREF and antifungal against ARCA. Marmycin A (Figure 5) showed activity against HCT-16 at IC50 60.5 nM, but the chlorinated analog (marmycin B) was less potent at IC50 1.09 μM [52]. This fact is in opposite that halogenation usually responsible for bioactivity and enhance the bioactivity.
3.4.6. Fijiolides
Marine-derived Actinobacteria genus Nocardiopsis isolated from sediment near Beqa Island in Beqa Lagoon, Fiji, produce Fijiolide A and B (Figure 5). Fijiolide A enhanced the activity of quinone reductase 1 (QR1), an enzyme that converts quinone to hydroquinone at concentration 28.4 μM. In addition, Fijiolide A reduced TNF-α-induced NF-ĸB activation to 70.3% and IC50 0.57 μM. In contrast, Fijiolide B did not exhibit activities suggesting that substitution on the nitrogen atom affects activity [53]. Arctic marine Actinomycetes identified as Streptomyces strain ART 5 was isolated from the arctic region, eastern Siberia during the RV Araon Arctic Expedition (ARA 03B). Profiling chemistry of fermentation broth yielded identification of fijiolide A and B along with articoside, C-1027-chromophore V, and C-1027-chromophore-III. The compounds were tested for bioactivity against Candida albicans and antiproliferative activities against human carcinoma cell lines. C-1027-chromophore V and C-1027-chromophore-III showed bioactivity against C. albicans at IC50 37.9 μM and 25.6 μM, respectively, but fijiolides showed no activity. The difference in the benzoxazine that counts for the activity. Antiproliferative activities of compounds were ranged from moderate to strong against cancer cell lines HCT-116, A549, SNU638, SK-HEP1, K562, and MDA-MB231 at IC50 0.6–44 μM [54].
3.4.7. Streptochloride
Chlorinated polyketide compound streptochlorides (Figure 5) were isolated from ethyl acetate extract of fermentation broth of Streptomyces sp. OUCMDZ-1703 associated with unidentified soft coral. Both compounds have modest antimicrobial activity against P. aeruginosa, E. coli, and S. aureus but no activity against MRSA. Streptochloride A and B demonstrated cytotoxicity against MCF-7 cell line at IC50 9.9 and 20.2 μmol/L, respectively [55].
4. Future Direction and Conclusions
Marine Actinobacteria have shown as producer of varied diversity of halometabolites compounds. The compounds range from simple to complicated structures with group under polyketides, peptides, alkaloid, and terpenoid. Some compounds demonstrate intriguing structure special for marine compounds. The compounds exhibited enormous potential for the discovery of new therapeutic leads in the development of drugs to fight the current antibiotic resistance threats, anticancer, and other bioactivities. Marine Actinobacteria produce more chlorometabolites than bromometabolites in contrast with sponges and red algae which are rich in bromometabolites.
To date, there are several bioprospecting programs with target marine biodiversity for novel bioactive metabolites including halometabolites. FADH2-dependent halogenase is the biggest group of halogenating enzymes, thus can be used as target in the bioprospecting of halometabolites from marine Actinobacteria. Genome mining by employing gene that encodes FADH2-dependent halogenase as an indicator has enabled to screen 555 genetic potentials of actinomycetes for halogenated natural products [56]. Gao and Huang employed the same approach to screen 228 Actinomycetes to find distribution of the gene and secondary metabolites [57]. Screening mangrove-derived Actinomycetes using FADH2-dependent halogenase resulted in 26 halogenase-positive strain among 163 isolates [58]. PCR-based marker gene screening was employed to detect FADH2-dependent halogenase gene of Arctic marine Actinobacteria. The study concluded that Arctic marine Actinobacteria are potential in halometabolites production [59]. Three novel halogenase gene clusters were identified in microbial metagenome of marine sponge indicated that the microbial consortia of sponges including marine Actinobacteria are a valuable resource for novel halogenation [60]. There is a correlation between the distribution of FADH2-dependent halogenase gene in filamentous actinomycetes and the potential for producing halometabolites.
Comparative genome studies showed that Actinobacteria are rich in secondary metabolites genes that never been explored, so the chance to discover new bioactive halometabolites is still wide open. Further research into mechanisms of biological halogenation will provide insight and a greater understanding of biosynthesis of halometabolites. Furthermore, understanding the genes encoding halogenase enzymes may be used to generate recombinant organisms to produce derivative new natural product. Advance technology in exploration and collection, compound isolation, purification, structure elucidation, bioassay, and high-throughput screening will ensure and enable to identify potential halometabolites from marine Actinobacteria for benefit to humankind.
Author Contributions
N.K. contributed in writing manuscript and funding; T.T. contributed in conceptualizing and reviewing the article.
Funding
This research was funded by TWAS for Individual Scientific Research Grant 11-189RG/CHE/AS to NK and Higher Education Research Grant Indonesia (Hibah Penelitian Unggulan Perguruan Tinggi Negeri) 2017 for Triyanto.
Acknowledgments
Maria Ulfah and Chandra Praditaningrum for their assistance in the preparation of references.
Conflicts of Interest
The authors declare no conflict of interest. And the funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gribble, G.W. Biological activity of recently discovered halogenated marine natural product. Mar. Drugs 2015, 13, 4044–4136. [Google Scholar] [CrossRef] [PubMed]
- Gribble, G.W. Amazing organohalogens. Am. Scientist 2004, 92, 342–349. [Google Scholar] [CrossRef]
- Gribble, G.W. The diversity of naturally produced organohalogen. Chemosphere 2003, 52, 289–297. [Google Scholar] [CrossRef]
- Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.-G.; Gloer, J.B.; Ji, N.-Y.; Zhao, J.-C. Halogenated organic molecules of Rhodomelaceae origin: Chemistry and Biology. Chem. Rev. 2013, 113, 3632–3683. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzer, R.A.; Prinsep, M.R. Marine natural product. Nat. Prod. Rep. 2018, 34, 235–294. [Google Scholar] [CrossRef] [PubMed]
- El-Demerdash, A.; Atanasov, A.G.; Horbanczuk, O.K.; Tammam, M.A.; Abdel-Mogib, M.; Hooper, J.N.A.; Sekeroglu, N.; Al-Mourabit, A.; Kijjoa, A. Chemical Diversity and Biological Activities of Marine Sponges of the Genus Suberea: A Systematic Review. Mar. Drugs 2019, 17, 115. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, S.K.; Rajendran, N.M.; Marino, A. Natural Products Diversity of Marine Ascidians (Tunicates; Ascidiacea) and Successful Drugs in Clinical Development. Nat. Prod. Biopros. 2017, 7, 1–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; Wezel, G.P. Taxonomy, Physiology, and Natural Product of Actinobacteria. Microb. Mol. Biol. Rev. 2016, 80, 1–44. [Google Scholar] [CrossRef]
- Rezanka, T.; Spizek, J. Halogen containing antibiotics from Streptomycetes. Stud. Nat. Prod. Chem. 2003, 29, 309–353. [Google Scholar]
- Berdy, J. Bioactive Microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Geniloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.C.; Bora, N. Diversity and biogeography of marine actinobacteria. Curr. Opin. Microbiol. 2006, 9, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Ulfah, M.; Kasanah, N.; Handayani, N.S.N. Bioactivity and genetic screening of marine Actinobacteria associated with red algae Gelidiella acerosa. Ind. J. Biotechnol. 2017, 22, 13–21. [Google Scholar] [CrossRef]
- Subramani, R.; Aalbersberg, W. Marine actinomycetes: An ongoing source of novel bioactive metabolites. Microbiol. Res. 2012, 167, 571–580. [Google Scholar] [CrossRef]
- Rahman, H.; Austin, B.; Mitchell, W.J.; Morris, P.C.; Jamieson, D.J.; Adams, D.R.; Spragg, A.M.; Schweizer, M. Novel Antiinfective compounds from marine Actinobacteria. Mar. Drugs 2010, 8, 498–518. [Google Scholar] [CrossRef]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.-K. Pharmaceutically active secondary metabolites. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef]
- Genilloud, O. Mining Actinomycetes for Novel Antibiotics in the Omics Era: Are We Ready to Exploit This New Paradigm? Antibiotics 2018, 7, 85. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nakashima, T. Actinomycetes, an Inexhaustible Source of Naturally Occurring Antibiotics. Antibiotics 2018, 7, 45. [Google Scholar] [CrossRef]
- Dhakal, D.; Pokhrel, A.R.; Shresta, B.; Sohng, J.K. Marine rare Actinobacteria: Isolation, Characterization, and Strategies for Harnessing Bioactive Compounds. Front. Microbiol. 2017, 8, 1106. [Google Scholar] [CrossRef]
- Schinke, C.; Martins, T.; Queiroz, S.C.N.; Melo, I.S.; Reyes, F.G. Antibacterial Compounds from marine bacteria 2010–2015. J. Nat. Prod. 2017, 80, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- McArthur, K.A.; Mitchell, S.S.; Tsueng, G.; Rheingold, A.; White, D.J.; Grodberg, J.; Lam, K.S.; Potts, B.C. Lynamicins A-E, chlorinated bisindole pyrrole antibiotics from a novel marine actinomycete. J. Nat. Prod. 2008, 71, 1732–1737. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.C.; Prieto-Davo, A.; Jensen, P.R.; Fenical, W. The marinopyrroles, antibiotics of an unpre- cedented structure class from a marine Streptomyces sp. Org. Lett. 2008, 10, 629–731. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.C.; Fenical, W. Structures, reactivities, and antibiotic properties of the marinopyrroles A–F. J. Org. Chem. 2010, 75, 3240–3250. [Google Scholar] [CrossRef] [PubMed]
- Haste, N.M.; Hughes, C.C.; Tran, D.N.; Fenical, W.; Jensen, P.R.; Nizet, V.; Hensler, M.E. Pharmacological properties of the marine natural product marinopyrrole A against meticillin-resistant Staphylococcus aureus. Antimic. Agents Chemother. 2011, 55, 3305–3312. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Li, R.; Sung, S.-S.; Wu, H.; Liu, Y.; Manieri, W.; Krishnegowda, G.; Awwad, A.; Dewey, A.; Liu, X.; et al. Discovery of marinopyrrole A (Maritoclax) as a selective Mcl-1 antagonist that overcomes ABT-737 resistance by binding to and targeting Mcl-1 for proteasomal degradation. J. Biol. Chem. 2012, 287, 10224–10235. [Google Scholar] [CrossRef]
- Maloney, K.N.; MacMillan, J.B.; Kauffman, C.A.; Jensen, P.R.; DiPasquale, A.G.; Rheingold, A.L.; Fenical, W. Lodopyridone, a structurally unprecedented alkaloid from a marine actinomycete. Org. Lett. 2009, 11, 5422–5424. [Google Scholar] [CrossRef]
- Hughes, C.C.; MacMillan, J.B.; Gaudêncio, S.P.; Jensen, P.R.; Fenical, W. The ammosamides: Structures of cell cycle modulators from a marine-derived streptomyces species. Angew. Chem. Int. Ed. Engl. 2009, 48, 725–730. [Google Scholar] [CrossRef]
- Pan, E.; Jamison, M.; Yousufuddin, M.; MacMillan, J.B. Ammosamide D, an oxidatively ring opened ammosamide analog from a marine-derived Streptomyces variabilis. Org. Lett. 2012, 14, 2390–2393. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Li, S.; Liu, Z.; Chen, Y.; Zhang, H.; Zhang, G.; Zhang, Q.; Tian, X.; Yuan, C. Spiroindimicin A–D: New bisindole alkaloids from the deep sea derived actinomycetes. Org. Lett. 2014, 14, 3364–3367. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Li, S.; Liu, Z.; Chen, Y.; Zhang, H.; Zhang, G.; Zhang, Q.; Tian, X.; Yuan, C. Indimicin A-E, bisindole alkaloids from the deep sea derived Streptomyces sp. SCSIO 03032. J. Nat. Prod. 2014, 77, 1887–18892. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Mico, X.; Jensen, P.R.; Fenical, W.; Hughes, C.C. Chlorizidine, a cytotoxic 5 h-pyrrolo [2,1-a] iso indol-5-one-containing alkaloid from a marine Streptomyces sp. Org. Lett. 2013, 15, 988–991. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kwon, H.C.; Williams, P.G.; Jensen, P.R.; Fenical, W. Azamerone, a terpenoid phthalazinone from a marine-derived bacterium related to the genus Streptomyces (Actinomycetales). Org. Lett. 2006, 8, 2471–2474. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.C.; Espindola, A.P.D.; Park, J.S.; Prieto-Davó, A.; Rose, M.; Jensen, P.R.; Fenical, W. Nitropyrrolins A–E, cytotoxic farnesyl-α-nitropyrroles from a marine-derived bacterium within the actinomycete family Streptomycetaceae. J. Nat. Prod. 2010, 73, 2047–2052. [Google Scholar] [CrossRef] [PubMed]
- Sakoulas, G.; Nam, S.J.; Loesgen, S.; Fenical, W.; Jensen, P.R.; Nizet, V.; Hensler, M. Novel bacterial metabolite merochlorin A demonstrates in vitro activity against multi-drug resistant methicillin-resistant Staphylococcus aureus. PLoS ONE 2012, 7, 1–57. [Google Scholar] [CrossRef]
- Kaysser, L.; Bernhardt, P.; Nam, S.J.; Loesgen, S.; Ruby, J.G.; Skewes-Cox, P.; Jensen, P.R.; Fenical, W.; Moore, B.S. Merochlorins A–D, cyclic meroterpenoid antibiotics biosynthesized in divergent pathways with vanadium-dependent chloroperoxidases. J. Am. Chem. Soc. 2012, 134, 11988–11991. [Google Scholar] [CrossRef] [PubMed]
- Kondratyuk, T.P.; Park, E.-J.; van Breemen, R.B.; Asolkar, R.N.; Murphy, B.T.; Fenical, W.; Pezzutto, J.M. Novel Marine Phenazines as potential cancer chemopreventive and antiinflamatory agents. Mar. Drugs 2012, 10, 451–464. [Google Scholar] [CrossRef]
- Asolkara, R.N.; Singh, A.; Jensen, P.R.; Aalbersbergb, W.; Cartéb, B.K.; Feussnerb, K.-D.; Subramani, R.; DiPasquale, Antonio.; Rheingold, A.L.; Fenicala, W. Marinocyanins, cytotoxic bromo-phenazinone meroterpenoids from a marine bacterium from the streptomycete clade MAR4. Tetrahedron 2017, 73, 2234–2241. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Li, S.; Li, J.; Chen, Y.; Saurav, K.; Zhang, Q.; Zhand, H.; Zhang, W.; Zhang, S.; Zhang, C. Antibacterial and cytotoxic new napyradiomycins from the marine-derived Streptomyces sp. SCSIO 10428. Mar. Drugs 2013, 11, 2113–2125. [Google Scholar] [CrossRef]
- Cheng, Y.-B.; Jensen, P.; Fenical, W. Cytotoxic and Antimicrobial Napiradiomycins from two marine derived MAR 4 Streptomyces Strain. Eur. J. Org. Chem. 2013, 18, 3751–3757. [Google Scholar] [CrossRef]
- Farnes, L.; Coufal, G.N.; Kauffman, C.A.; Rheingold, A.L.; Dipasquale, A.G.; Jensen, P.R.; Fenical, W. Napyradiomycin derivates, produced by a merine derived Actinomycete, illustrate cytotoxicity by induction of apoptosis. J. Nat. Prod. 2013, 77, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lacret, R.; Perez-Victoria, I.; Oves-Costales, D.; de la Cruz, M.; Domingo, E.; Martin, J.; Diaz, C.; Vicente, F.; Geniloud, O.; Reyes, F. MDN-0170, a New napyradiomycin from Streptomyces sp. strain CA-271078. Mar. Drugs 2016, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.D.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Piperazimycins: Cytotoxic hexapeptides from a marine-derived bacterium of the genus Streptomyces. J. Org. Chem. 2007, 72, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, K.; Takagi, M.; Shin-Ya, K. Tetrapeptides possessing a unique skeleton, JBIR-34 and JBIR-35, isolated from a sponge-derived actinomycete, Streptomyces sp. Sp080513GE-23. J. Nat. Prod. 2010, 73, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Flores, M.; Forteza, I.; Henriksen, N.M.; Concepcion, G.P.; Rosenberg, G.; Haygood, M.G.; Oliver, B.M.; Light, A.R.; Cheatham, I.T.E.; et al. Totopotensamides, polyketide–cyclic peptide hybrids from a mollusk-associated acterium Streptomyces sp. J. Nat. Prod. 2012, 75, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Feling, R.H.; Buchanan, G.O.; Mincer, T.J.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Salinosporamide A: A highly cytotoxic proteasome inhibitor from a novel microbial source, a marine bacterium of the new genus Salinospora. Angew. Chem. Int. Ed. Eng. 2003, 42, 355–357. [Google Scholar] [CrossRef]
- Williams, P.G.; Buchanan, G.O.; Feling, R.H.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. New cytotoxic salinosporamides from the marine actinomycete Salinispora tropica. J. Org. Chem. 2005, 70, 6193–6203. [Google Scholar] [CrossRef]
- Buchanan, G.O.; Williams, P.G.; Feling, R.H.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Sporolides A and B: Structurally unprecedented halogenated macrolides from the marine actinomycete Salinispora tropica. Org. Lett. 2005, 7, 2731–2734. [Google Scholar] [CrossRef]
- Li, F.; Maskey, R.P.; Qin, S.; Sattler, I.; Fiebig, H.H.; Maier, A.; Zeeck, A.; Laatsch, H. Chinikomycins A and B: Isolation, structure elucidation, and biological activity of novel antibiotics from a marine Streptomyces sp. isolate M045. J. Nat. Prod. 2005, 68, 349–353. [Google Scholar] [CrossRef]
- Oh, D.-C.; Williams, P.G.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Cyanosporasides A and B, cyano- and chloro-cyclopenta [a]indene glycosides from the deep-sea marine actinomycete “Salinispora pacifica”. Org. Lett. 2006, 8, 1021–1024. [Google Scholar] [CrossRef]
- Lane, A.L.; Nam, S.J.; Fukuda, T.; Yamanaka, K.; Kauffman, C.A.; Jensen, P.R.; Fenical, W.; Moore, B.S. Structures and comparative characterization of biosynthetic gene clusters for cyanosporoides, enediyne-derived natural products from marine actinomycetes. J. Am. Chem. Soc. 2013, 135, 4171–4174. [Google Scholar] [CrossRef]
- Martin, G.D.; Tan, L.T.; Jensen, P.R.; Dimayuga, R.E.; Fairchild, C.R.; Raventos-Suarez, C.; Fenical, W. Marmycins A and B, cytotoxic pentacyclic C-glycosides from a marine sediment-derived actinomycete related to the genus Streptomyces. J. Nat. Prod. 2007, 70, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.J.; Gaudêncio, S.P.; Kauffman, C.A.; Jensen, P.R.; Kondratyuk, T.P.; Marler, L.E.; Pezzuto, J.M.; Fenical, W. Fijiolides A and B, inhibitors of TNF-α-induced NF-κB activation, from a marine-derived sediment bacterium of the genus Nocardiopsis. J. Nat. Prod. 2010, 73, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Ahn, C.H.; Shin, Y.; Won, T.H.; Ko, K.; Lee, S.K.; Oh, K.B.; Shin, J.; Nam, S.I.; Oh, D.C. New benzoxazine secondary metabolites from an arctic actinomycete. Mar. Drugs 2014, 12, 2526–2538. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Kong, F.; Wang, Y.; Wang, Y.; Liu, P.; Zuo, G.; Zhu, W. Antibiotic metabolites from the coral-associated actinomycete Streptomyces sp. OUCMDZ-1703. Chin. J. Chem. 2013, 31, 100–104. [Google Scholar] [CrossRef]
- Hornung, A.; Bertazzo, M.; Dziarnowski, A.; Schneider, K.; Welzel, K.; Wohlert, S.E.; Holzenkampfer, M.; Nicholson, G.J.; Bechthold, A.; Sussmuth, R.D.; et al. Genomic screening approach to the structure-guided identification of drug candidates from natural sources. Chem. Bio. Chem. 2007, 8, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Huang, Y. Detection, distribution, and organohalogen compound discovery implications of the reduced flavin adenine dinucleotide-dependent halogenase gene in major filamentous actinomycete taxonomic groups. Appl. Environ. Microbiol. 2009, 75, 4813–4820. [Google Scholar] [CrossRef]
- Li, X.-G.; Tang, X.-M.; Xiao, J.; Ma, G.-H.; Xu, L.; Xie, S.-J.; Xu, M.-J.; Xiao, X.; Xu, J. Harnessing the potential of halogenated natural product biosynthesis by mangrove-derived actinomycetes. Mar. Drugs 2013, 11, 3875–3890. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Chen, R.; Jiang, M.; Tian, X.; Liu, H.; Yu, Y.; Fan, C.; Chen, B. Bioprospecting potential of halogenases from Arctic marine Actinomycetes. BMC Microbiol. 2016, 16, 1–9. [Google Scholar] [CrossRef]
- Bayer, K.; Scheuermayer, M.; Fieseler, L.; Hentschel, U. Genomic mining for novel FADH2-dependent halo genases in marine sponge-associated microbial consortia. Mar. Biotechnol. 2013, 15, 63–72. [Google Scholar] [CrossRef]
Figure 1.
Halometabolites from terrestrial Actinobacteria.


Figure 2.
Halogenated alkaloid.


Figure 3.
Halogenated terpene and monoterpene.



Figure 4.
Halogenated peptides.

Figure 5.
Halogenated polyketide.


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kasanah, N.; Triyanto, T. Bioactivities of Halometabolites from Marine Actinobacteria. Biomolecules 2019, 9, 225. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060225
AMA Style
Kasanah N, Triyanto T. Bioactivities of Halometabolites from Marine Actinobacteria. Biomolecules. 2019; 9(6):225. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060225
Chicago/Turabian StyleKasanah, Noer, and Triyanto Triyanto. 2019. "Bioactivities of Halometabolites from Marine Actinobacteria" Biomolecules 9, no. 6: 225. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9060225
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.