A Remote Sensing Algorithm of Column-Integrated Algal Biomass Covering Algal Bloom Conditions in a Shallow Eutrophic Lake


Abstract
:1. Introduction
2. Study Area and Data
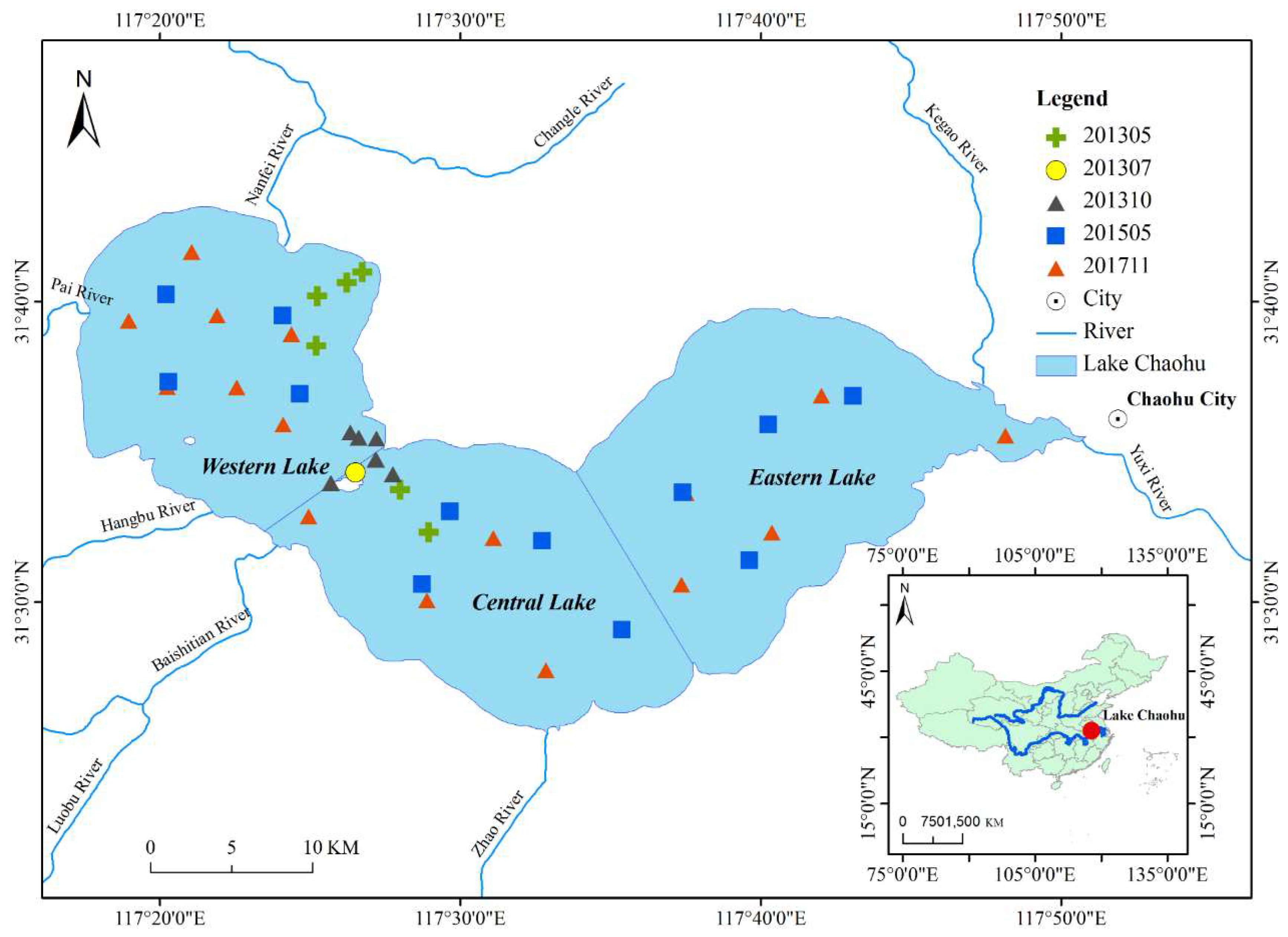
2.1. Study Site
2.2. In Situ Data
2.3. Remote Sensing Data
3. Development of Column-Integrated Algal Biomass Algorithm
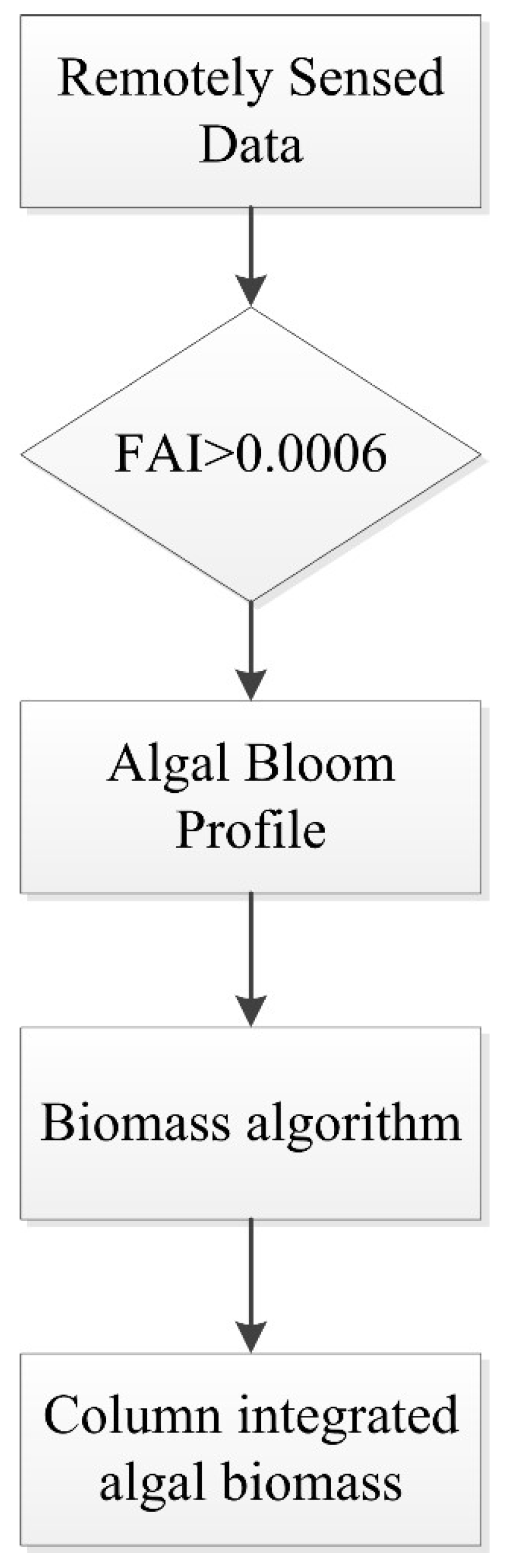
3.1. Algorithm Approach (Flow Chart)
3.2. Depth-Integrated Algal Biomass Retrieval from the Surface Investigation
Rrc(555)’ = Rrc(555) − [Rrc(469)] × (859 − 555)/(859 − 469) + Rrc(859) × (555 − 469)/(859 − 469)]
Rrc(645)’ = Rrc(645) − [Rrc(469)] × (859 − 645)/(859 − 469) + Rrc(859) × (645 − 469)/(859 − 469)]
b = 6.453 × z − 2.028
3.3. Performance Evaluation
4. Results
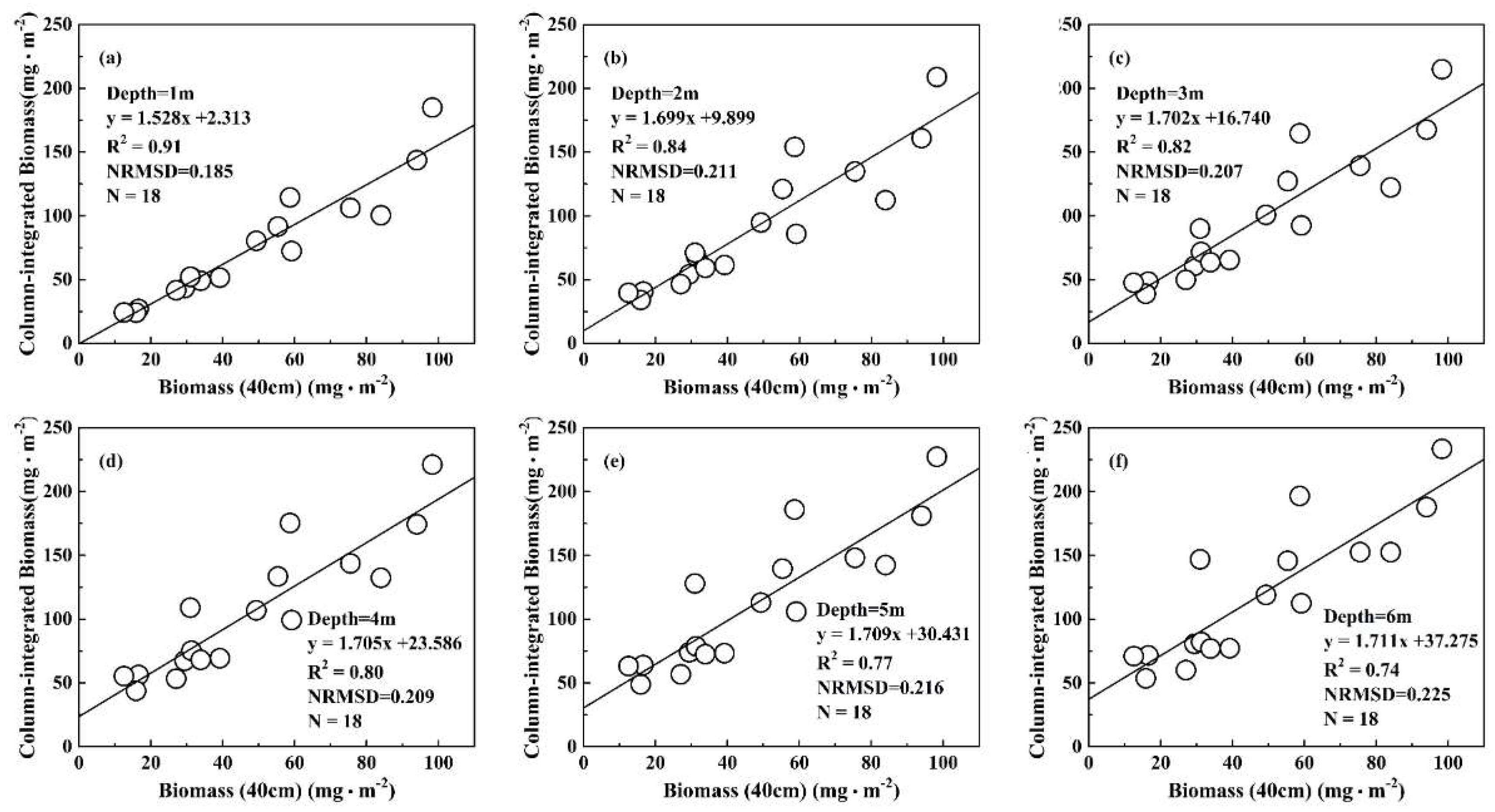
4.1. Field Observation of Column-Integrated Algal Biomass under Algal Bloom Conditions
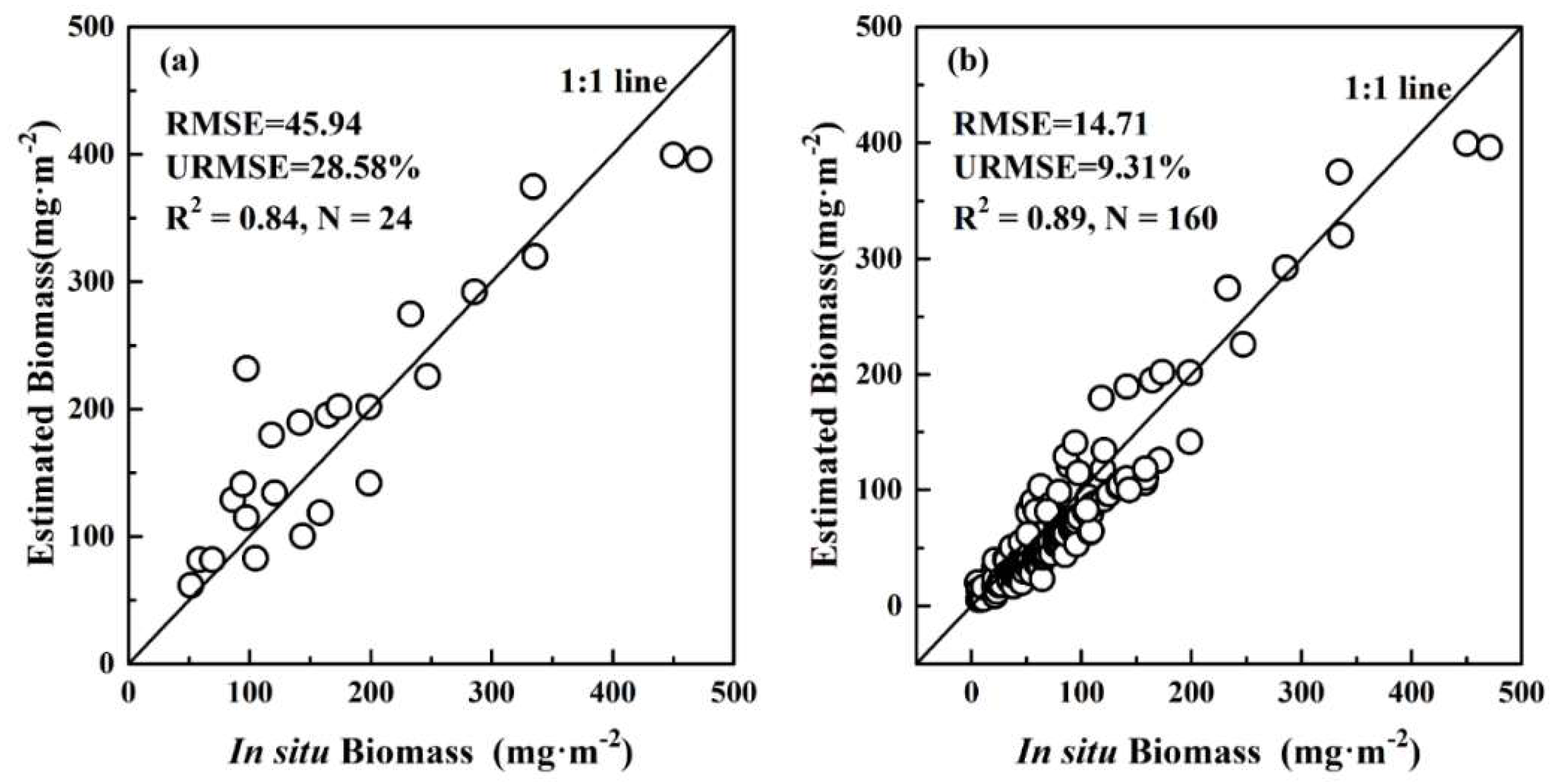
4.2. Algorithm Validation
4.2.1. Water Column Validation
4.2.2. Area-Integrated Validation
4.3. Sensitivity Analysis
5. Discussion
5.1. Comparison with Other Algorithms
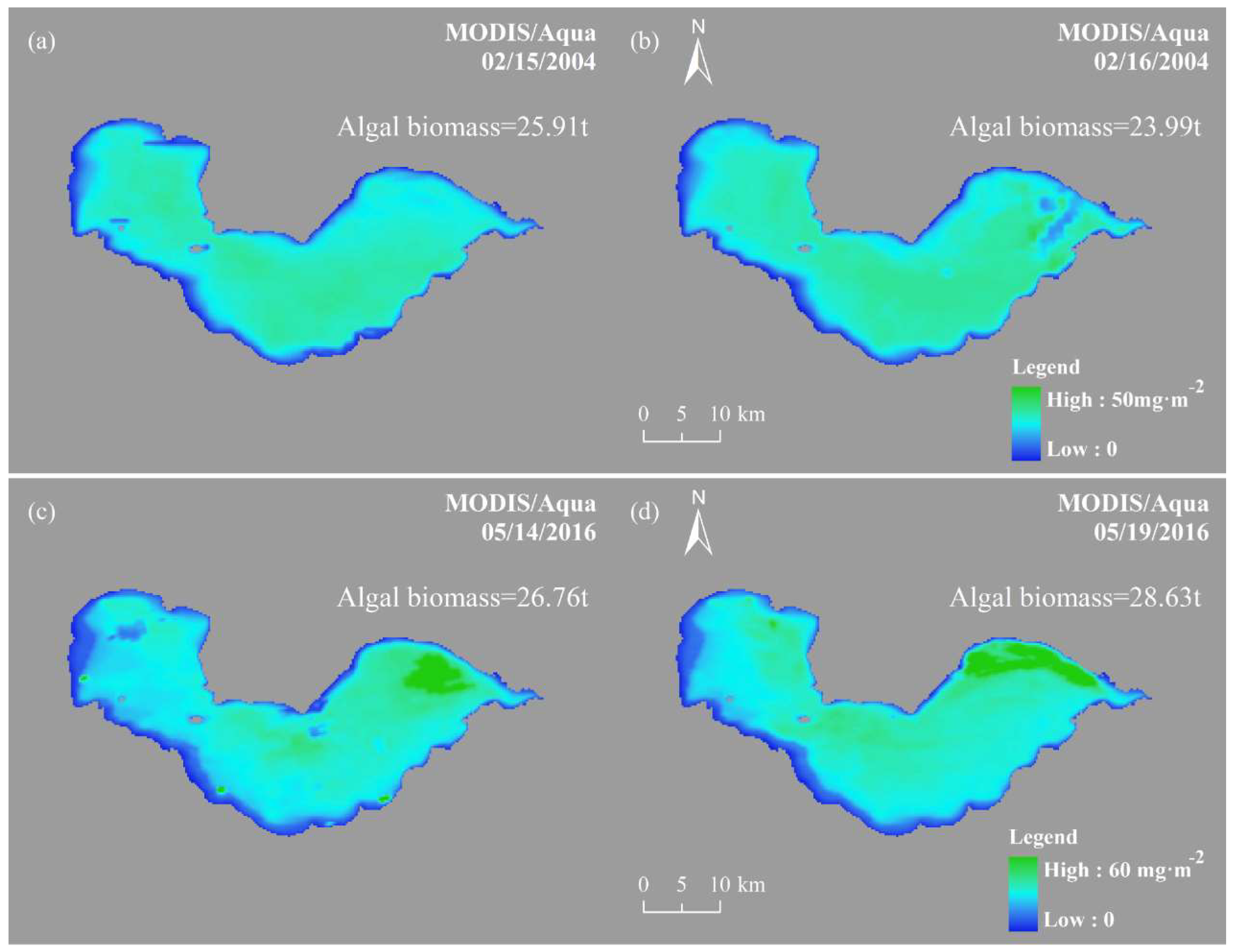
5.2. Analysis of Variation in Spatial Distribution
5.3. Extension and Application of the Algorithm
5.3.1. Extending Application to other Lakes
5.3.2. Algorithm for Data from Other Sensors
5.4. Algorithm Limitations
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L. Freshwater biodiversity: Importance, threats, status and conservation challenges. Boil. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr. 2006, 51, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K.; Smith, V.H. Nitrogen, phosphorus, and eutrophication in streams. Inland Waters 2016, 6, 155–164. [Google Scholar] [CrossRef]
- Padisák, J.; Reynolds, C.S. Shallow lakes: The absolute, the relative, the functional and the pragmatic. Hydrobiologia 2003, 506, 1–11. [Google Scholar] [CrossRef]
- Hunter, P.D.; Matthews, M.W.; Kutser, T.; Tyler, A.N. Remote Sensing of Cyanobacterial Blooms in Inland, Coastal, and Ocean Waters. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 89–99. [Google Scholar]
- Brivio, P.; Giardino, C.; Zilioli, E. Determination of chlorophyll concentration changes in Lake Garda using an image-based radiative transfer code for Landsat TM images. Int. J. Remote Sens. 2001, 22, 487–502. [Google Scholar] [CrossRef]
- Giardino, C.; Brando, V.E.; Dekker, A.G.; Strömbeck, N.; Candiani, G. Assessment of water quality in Lake Garda (Italy) using Hyperion. Remote Sens. Environ. 2007, 109, 183–195. [Google Scholar] [CrossRef]
- Lou, X.; Hu, C. Diurnal changes of a harmful algal bloom in the East China Sea: Observations from GOCI. Remote Sens. Environ. 2014, 140, 562–572. [Google Scholar] [CrossRef]
- Duan, H.; Ma, R.; Hu, C. Evaluation of remote sensing algorithms for cyanobacterial pigment retrievals during spring bloom formation in several lakes of East China. Remote Sens. Environ. 2012, 126, 126–135. [Google Scholar] [CrossRef]
- Tebbs, E.; Remedios, J.; Harper, D. Remote sensing of chlorophyll-a as a measure of cyanobacterial biomass in Lake Bogoria, a hypertrophic, saline–alkaline, flamingo lake, using Landsat ETM+. Remote Sens. Environ. 2013, 135, 92–106. [Google Scholar] [CrossRef]
- Omondi, R.; Yasindi, A.; Magana, A. Spatial and temporal variations of zooplankton in relation to some environmental factors in Lake Baringo, Kenya. Egerton J. Sci. Technol. 2015, 11, 29–50. [Google Scholar]
- Hecky, R. The eutrophication of lake Victoria. Int. Ver. Theor. Angew. Limnol. Verh. 1992, 25, 39–48. [Google Scholar] [CrossRef]
- Mbonde, A.S.; Sitoki, L.; Kurmayer, R. Phytoplankton composition and microcystin concentrations in open and closed bays of Lake Victoria, Tanzania. Aquat. Ecosyst. Health Manag. 2015, 18, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scavia, D.; Allan, J.D.; Arend, K.K.; Bartell, S.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloğlu, I.; DePinto, J.V. Assessing and addressing the re-eutrophication of Lake Erie: Central basin hypoxia. J. Great Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef] [Green Version]
- Kane, D.D.; Conroy, J.D.; Richards, R.P.; Baker, D.B.; Culver, D.A. Re-eutrophication of Lake Erie: Correlations between tributary nutrient loads and phytoplankton biomass. J. Great Lakes Res. 2014, 40, 496–501. [Google Scholar] [CrossRef]
- LaBuhn, S.; Klump, J.V. Estimating summertime epilimnetic primary production via in situ monitoring in an eutrophic freshwater embayment, Green Bay, Lake Michigan. J. Great Lakes Res. 2016, 42, 1026–1035. [Google Scholar] [CrossRef]
- Sawyers, J.E.; McNaught, A.S.; King, D.K. Recent and historic eutrophication of an island lake in northern Lake Michigan, USA. J. Paleolimnol. 2016, 55, 97–112. [Google Scholar] [CrossRef]
- Kulkarni, A. Water quality retrieval from Landsat TM imagery. Procedia Comput. Sci. 2011, 6, 475–480. [Google Scholar] [CrossRef]
- Cumming, B.F.; Laird, K.R.; Gregory-Eaves, I.; Simpson, K.G.; Sokal, M.A.; Nordin, R.; Walker, I.R. Tracking past changes in lake-water phosphorus with a 251-lake calibration dataset in British Columbia: Tool development and application in a multiproxy assessment of eutrophication and recovery in Osoyoos Lake, a transboundary lake in Western North America. Front. Ecol. Evol. 2015, 3, 84. [Google Scholar]
- Spetter, C.V.; Popovich, C.A.; Arias, A.; Asteasuain, R.O.; Freije, R.H.; Marcovecchio, J.E. Role of nutrients in phytoplankton development during a winter diatom bloom in a eutrophic South American estuary (Bahía Blanca, Argentina). J. Coast. Res. 2013, 31, 76–87. [Google Scholar] [CrossRef]
- Guinder, V.; Popovich, C.; Perillo, G. Phytoplankton and physicochemical analysis on the water system of the temperate estuary in South America: Bahía Blanca Estuary, Argentina. Int. J. Environ. Res. 2012, 6, 547–556. [Google Scholar]
- Brodie, J.; Devlin, M.; Haynes, D.; Waterhouse, J. Assessment of the eutrophication status of the Great Barrier Reef lagoon (Australia). Biogeochemistry 2011, 106, 281–302. [Google Scholar] [CrossRef]
- Blondeau-Patissier, D.; Schroeder, T.; Brando, V.E.; Maier, S.W.; Dekker, A.G.; Phinn, S. ESA-MERIS 10-year mission reveals contrasting phytoplankton bloom dynamics in two tropical regions of Northern Australia. Remote Sens. 2014, 6, 2963–2988. [Google Scholar] [CrossRef]
- Otten, T.G.; Crosswell, J.R.; Mackey, S.; Dreher, T.W. Application of molecular tools for microbial source tracking and public health risk assessment of a Microcystis bloom traversing 300 km of the Klamath River. Harmful Algae 2015, 46, 71–81. [Google Scholar] [CrossRef]
- Meneely, J.P.; Elliott, C.T. Microcystins: Measuring human exposure and the impact on human health. Biomarkers 2013, 18, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.; Tyler, A.; Willby, N.; Gilvear, D. The spatial dynamics of vertical migration by Microcystis aeruginosa in a eutrophic shallow lake: A case study using high spatial resolution time—Series airborne remote sensing. Limnol. Oceanogr. 2008, 53, 2391–2406. [Google Scholar] [CrossRef]
- Hu, C.; Lee, Z.; Ma, R.; Yu, K.; Li, D.; Shang, S. Moderate resolution imaging spectroradiometer (MODIS) observations of cyanobacteria blooms in Taihu Lake, China. J. Geophys. Res. Oceans 2010, 115. [Google Scholar] [CrossRef]
- Wynne, T.; Stumpf, R.; Tomlinson, M.; Warner, R.; Tester, P.; Dyble, J.; Fahnenstiel, G. Relating spectral shape to cyanobacterial blooms in the Laurentian Great Lakes. Int. J. Remote Sens. 2008, 29, 3665–3672. [Google Scholar] [CrossRef]
- Gower, J.; Doerffer, R.; Borstad, G. Interpretation of the 685 nm peak in water-leaving radiance spectra in terms of fluorescence, absorption and scattering, and its observation by MERIS. Int. J. Remote Sens. 1999, 20, 1771–1786. [Google Scholar] [CrossRef]
- Gower, J.; King, S.; Borstad, G.; Brown, L. Detection of intense plankton blooms using the 709 nm band of the MERIS imaging spectrometer. Int. J. Remote Sens. 2005, 26, 2005–2012. [Google Scholar] [CrossRef]
- Matthews, M.W.; Bernard, S.; Robertson, L. An algorithm for detecting trophic status (chlorophyll-a), cyanobacterial-dominance, surface scums and floating vegetation in inland and coastal waters. Remote Sens. Environ. 2012, 124, 637–652. [Google Scholar] [CrossRef]
- Li, L.; Li, L.; Song, K. Remote sensing of freshwater cyanobacteria: An extended IOP Inversion Model of Inland Waters (IIMIW) for partitioning absorption coefficient and estimating phycocyanin. Remote Sens. Environ. 2015, 157, 9–23. [Google Scholar] [CrossRef]
- Ogashawara, I.; Moreno-Madriñán, M.J. Improving inland water quality monitoring through remote sensing techniques. ISPRS Int. J. Geo-Inf. 2014, 3, 1234–1255. [Google Scholar] [CrossRef]
- Ha, N.T.T.; Thao, N.T.P.; Koike, K.; Nhuan, M.T. Selecting the Best Band Ratio to Estimate Chlorophyll-a Concentration in a Tropical Freshwater Lake Using Sentinel 2A Images from a Case Study of Lake Ba Be (Northern Vietnam). ISPRS Int. J. Geo-Inf. 2017, 6, 290. [Google Scholar] [CrossRef]
- Qi, L.; Hu, C.; Duan, H.; Cannizzaro, J.; Ma, R. A novel MERIS algorithm to derive cyanobacterial phycocyanin pigment concentrations in a eutrophic lake: Theoretical basis and practical considerations. Remote Sens. Environ. 2014, 154, 298–317. [Google Scholar] [CrossRef]
- Randolph, K.; Wilson, J.; Tedesco, L.; Li, L.; Pascual, D.L.; Soyeux, E. Hyperspectral remote sensing of cyanobacteria in turbid productive water using optically active pigments, chlorophyll a and phycocyanin. Remote Sens. Environ. 2008, 112, 4009–4019. [Google Scholar] [CrossRef]
- Gorham, T.; Jia, Y.; Shum, C.; Lee, J. Ten-year survey of cyanobacterial blooms in Ohio’s waterbodies using satellite remote sensing. Harmful Algae 2017, 66, 13–19. [Google Scholar] [CrossRef]
- Hu, L.; Hu, C.; Ming-Xia, H. Remote estimation of biomass of Ulva prolifera macroalgae in the Yellow Sea. Remote Sens. Environ. 2017, 192, 217–227. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Oliver, R.L.; Walsby, A.E. Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. N. Z. J. Mar. Freshw. Res. 1987, 21, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Walsby, A.; Utkilen, H.; Johnsen, I. Buoyancy changes of a red coloured Oscillatoria agardhii in Lake Gjersjoen, Norway. Arch. Hydrobiol. 1983, 97, 18–38. [Google Scholar]
- Walsby, A.; Schanz, F. Light—Dependent growth rate determines changes in the population of Planktothrix rubescens over the annual cycle in Lake Zürich, Switzerland. New Phytol. 2002, 154, 671–687. [Google Scholar] [CrossRef]
- Walsby, A.E. Stratification by cyanobacteria in lakes: A dynamic buoyancy model indicates size limitations met by Planktothrix rubescens filaments. New Phytol. 2005, 168, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Hu, C.; Visser, P.M.; Ma, R. Diurnal changes of cyanobacteria blooms in Taihu Lake as derived from GOCI observations. Limnol. Oceanogr. 2018, 63, 1711–1726. [Google Scholar] [CrossRef] [Green Version]
- Sathyendranath, S.; Prieur, L.; Morel, A. A three-component model of ocean colour and its application to remote sensing of phytoplankton pigments in coastal waters. Int. J. Remote Sens. 1989, 10, 1373–1394. [Google Scholar] [CrossRef]
- Frolov, S.; Ryan, J.; Chavez, F. Predicting euphotic-depth-integrated chlorophyll-afrom discrete-depth and satellite-observable chlorophyll-a off central California. J. Geophys. Res. Oceans 2012, 117. [Google Scholar] [CrossRef]
- Uitz, J.; Claustre, H.; Morel, A.; Hooker, S.B. Vertical distribution of phytoplankton communities in open ocean: An assessment based on surface chlorophyll. J. Geophys. Res. Oceans 2006, 111. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.R.; Cullen, J.J.; Platt, T. Phytoplankton and thermal structure in the upper ocean: Consequences of nonuniformity in chlorophyll profile. J. Geophys. Res. Oceans 1983, 88, 2565–2570. [Google Scholar] [CrossRef]
- Odermatt, D.; Gitelson, A.; Brando, V.E.; Schaepman, M. Review of constituent retrieval in optically deep and complex waters from satellite imagery. Remote Sens. Environ. 2012, 118, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.; Aiken, J.; Lavender, S. The atmospheric correction of water colour and the quantitative retrieval of suspended particulate matter in Case II waters: Application to MERIS. Int. J. Remote Sens. 1999, 20, 1713–1733. [Google Scholar] [CrossRef]
- Nouchi, V.; Odermatt, D.; Wüest, A.; Bouffard, D. Effects of non-uniform vertical constituent profiles on remote sensing reflectance of oligo-to mesotrophic lakes. Eur. J. Remote Sens. 2018, 51, 808–821. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Ma, R.; Duan, H.; Loiselle, S.; Xue, K.; Liang, Q. Satellite-based estimation of column-integrated algal biomass in nonalgae bloom conditions: A case study of Lake Chaohu, China. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 450–462. [Google Scholar] [CrossRef]
- Matsumoto, K.; Honda, M.C.; Sasaoka, K.; Wakita, M.; Kawakami, H.; Watanabe, S. Seasonal variability of primary production and phytoplankton biomass in the western Pacific subarctic gyre: Control by light availability within the mixed layer. J. Geophys. Res. Oceans 2014, 119, 6523–6534. [Google Scholar] [CrossRef] [Green Version]
- Sverdrup, H. On conditions for the vernal blooming of phytoplankton. J. Cons. 1953, 18, 287–295. [Google Scholar] [CrossRef]
- Chiswell, S.M. Annual cycles and spring blooms in phytoplankton: Don’t abandon Sverdrup completely. Mar. Ecol. Prog. Ser. 2011, 443, 39–50. [Google Scholar] [CrossRef]
- Xue, K.; Zhang, Y.; Duan, H.; Ma, R.; Loiselle, S.; Zhang, M. A remote sensing approach to estimate vertical profile classes of phytoplankton in a eutrophic lake. Remote Sens. 2015, 7, 14403–14427. [Google Scholar] [CrossRef]
- Chen, J.; Quan, W. An improved algorithm for retrieving chlorophyll-a from the Yellow River Estuary using MODIS imagery. Environ. Monit. Assess. 2013, 185, 2243–2255. [Google Scholar] [CrossRef]
- Yin, H.; Deng, J.; Shao, S.; Gao, F.; Gao, J.; Fan, C. Distribution characteristics and toxicity assessment of heavy metals in the sediments of Lake Chaohu, China. Environ. Monit. Assess. 2011, 179, 431–442. [Google Scholar] [CrossRef]
- Jiang, Y.-J.; He, W.; Liu, W.-X.; Qin, N.; Ouyang, H.-L.; Wang, Q.-M.; Kong, X.-Z.; He, Q.-S.; Yang, C.; Yang, B. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Zhang, M.; Duan, H.; Loiselle, S.; Xu, J. Fourteen-year record (2000–2013) of the spatial and temporal dynamics of floating algae blooms in Lake Chaohu, observed from time series of MODIS images. Remote Sens. 2015, 7, 10523–10542. [Google Scholar] [CrossRef]
- Cai, Y.; Kong, F.; Shi, L.; Yu, Y. Spatial heterogeneity of cyanobacterial communities and genetic variation of Microcystis populations within large, shallow eutrophic lakes (Lake Taihu and Lake Chaohu, China). J. Environ. Sci. 2012, 24, 1832–1842. [Google Scholar] [CrossRef]
- Jiang, Y.; Cheng, B.; Liu, M.; Nie, Y. Spatial and temporal variations of taste and odor compounds in surface water, overlying water and sediment of the Western Lake Chaohu, China. Bull. Environ. Contam. Toxicol. 2016, 96, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.; Du, M. The dynamics of Microcystis genotypes and microcystin production and associations with environmental factors during blooms in Lake Chaohu, China. Toxins 2014, 6, 3238–3257. [Google Scholar] [CrossRef]
- Zhou, Y.; Jin, J.; Liu, L.; Zhang, L.; He, J.; Wang, Z. Inference of reference conditions for nutrient concentrations of Chaohu Lake based on model extrapolation. Chin. Geogr. Sci. 2013, 23, 35–48. [Google Scholar] [CrossRef]
- Mueller, J.L.; Fargion, G.S.; McClain, C.R.; Pegau, S.; Zanefeld, J.; Mitchell, B.G.; Kahru, M.; Wieland, J.; Stramska, M. Ocean Optics Protocols for Satellite Ocean Color Sensor Validation, Revision 4, Volume IV: Inherent Optical Properties: Instruments, Characterizations, Field Measurements and Data Analysis Protocols; Aeronautics and Space Administration of USA: Greenbelt, MD, USA, 2003.
- Welschmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Hu, C.; Duan, H.; Barnes, B.B.; Ma, R. An EOF-based algorithm to estimate chlorophyll a concentrations in Taihu Lake from MODIS land-band measurements: Implications for near real-time applications and forecasting models. Remote Sens. 2014, 6, 10694–10715. [Google Scholar] [CrossRef]
- Franz, B.A.; Werdell, P.J.; Meister, G.; Kwiatkowska, E.J.; Bailey, S.W.; Ahmad, Z.; McClain, C.R. MODIS land bands for ocean remote sensing applications. In Proceedings of the Ocean Optics XVIII, Montreal, QC, Canada, 9–13 October 2006. [Google Scholar]
- Zhang, Y.; Ma, R.; Duan, H.; Loiselle, S.; Zhang, M.; Xu, J. A novel MODIS algorithm to estimate chlorophyll a concentration in eutrophic turbid lakes. Ecol. Indic. 2016, 69, 138–151. [Google Scholar] [CrossRef]
- Morel, A.; Berthon, J.F. Surface pigments, algal biomass profiles, and potential production of the euphotic layer: Relationships reinvestigated in view of remote-sensing applications. Limnol. Oceanogr. 1989, 34, 1545–1562. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.G.; Xie, P.; Zhou, Q.; Yang, H.; Guo, L.G. Studies on temporal and spatial variations of phytoplankton in Lake Chaohu. J. Integr. Plant Boil. 2007, 49, 409–418. [Google Scholar] [CrossRef]
- Wynne, T.T.; Stumpf, R.P.; Tomlinson, M.C.; Dyble, J. Characterizing a cyanobacterial bloom in western Lake Erie using satellite imagery and meteorological data. Limnol. Oceanogr. 2010, 55, 2025–2036. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chl-a | Algal Biomass | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Depth (m) | Mean | Max | Min | SD | CV(%) | Mean | Max | Min | SD | CV(%) |
| 0.1 | 136.40 | 313.85 | 55.43 | 71.44 | 52.37 | 14.82 | 33.10 | 5.85 | 7.51 | 50.69 |
| 0.2 | 96.70 | 217.50 | 24.67 | 59.73 | 61.77 | 11.65 | 26.57 | 4.01 | 6.42 | 55.09 |
| 0.4 | 70.83 | 195.62 | 20.91 | 43.13 | 60.89 | 14.45 | 33.14 | 4.54 | 8.37 | 57.97 |
| 0.7 | 47.77 | 113.88 | 15.20 | 29.59 | 61.94 | 17.79 | 46.43 | 6.49 | 10.54 | 59.25 |
| 1 | 32.56 | 90.16 | 12.39 | 20.96 | 64.36 | 12.05 | 27.91 | 4.14 | 7.29 | 60.51 |
| 1.5 | 23.52 | 59.40 | 6.19 | 13.93 | 59.22 | 14.02 | 37.39 | 6.61 | 8.56 | 61.08 |
| 2 | 12.95 | 36.54 | 3.39 | 8.88 | 68.59 | 9.12 | 20.97 | 2.40 | 5.32 | 58.39 |
| 3 | 9.31 | 36.54 | 3.39 | 8.42 | 90.48 | 11.13 | 36.54 | 3.39 | 8.25 | 74.12 |
| Total Algal Biomass Variation (%) | |||
|---|---|---|---|
| Parameter variation | 5% | 10% | 20% |
| Bio(40) | 4.10 | 8.20 | 16.41 |
| z | 1.03 | 2.05 | 4.07 |
| Algorithms | R2 | RMSE | URMSE(100%) | Reference |
|---|---|---|---|---|
| MB89 | 0.6204 | 278.9908 | 0.8503 | [72] |
| Uitz proposed | 0.6201 | 599.0637 | 1.1607 | [49] |
| Critical depth based | 0.0721 | 252.8243 | 1.6160 | [57] |
| This study | 0.84 | 45.94 | 0.2858 | -- |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, R.; Xue, K.; Zhang, Y.; Loiselle, S. A Remote Sensing Algorithm of Column-Integrated Algal Biomass Covering Algal Bloom Conditions in a Shallow Eutrophic Lake. ISPRS Int. J. Geo-Inf. 2018, 7, 466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7120466
Li J, Ma R, Xue K, Zhang Y, Loiselle S. A Remote Sensing Algorithm of Column-Integrated Algal Biomass Covering Algal Bloom Conditions in a Shallow Eutrophic Lake. ISPRS International Journal of Geo-Information. 2018; 7(12):466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7120466
Chicago/Turabian StyleLi, Jing, Ronghua Ma, Kun Xue, Yuchao Zhang, and Steven Loiselle. 2018. "A Remote Sensing Algorithm of Column-Integrated Algal Biomass Covering Algal Bloom Conditions in a Shallow Eutrophic Lake" ISPRS International Journal of Geo-Information 7, no. 12: 466. https://0-doi-org.brum.beds.ac.uk/10.3390/ijgi7120466