
Honokiol and Magnolol: Insights into Their Antidermatophytic Effects

 ,
,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antifungal Susceptibility Results
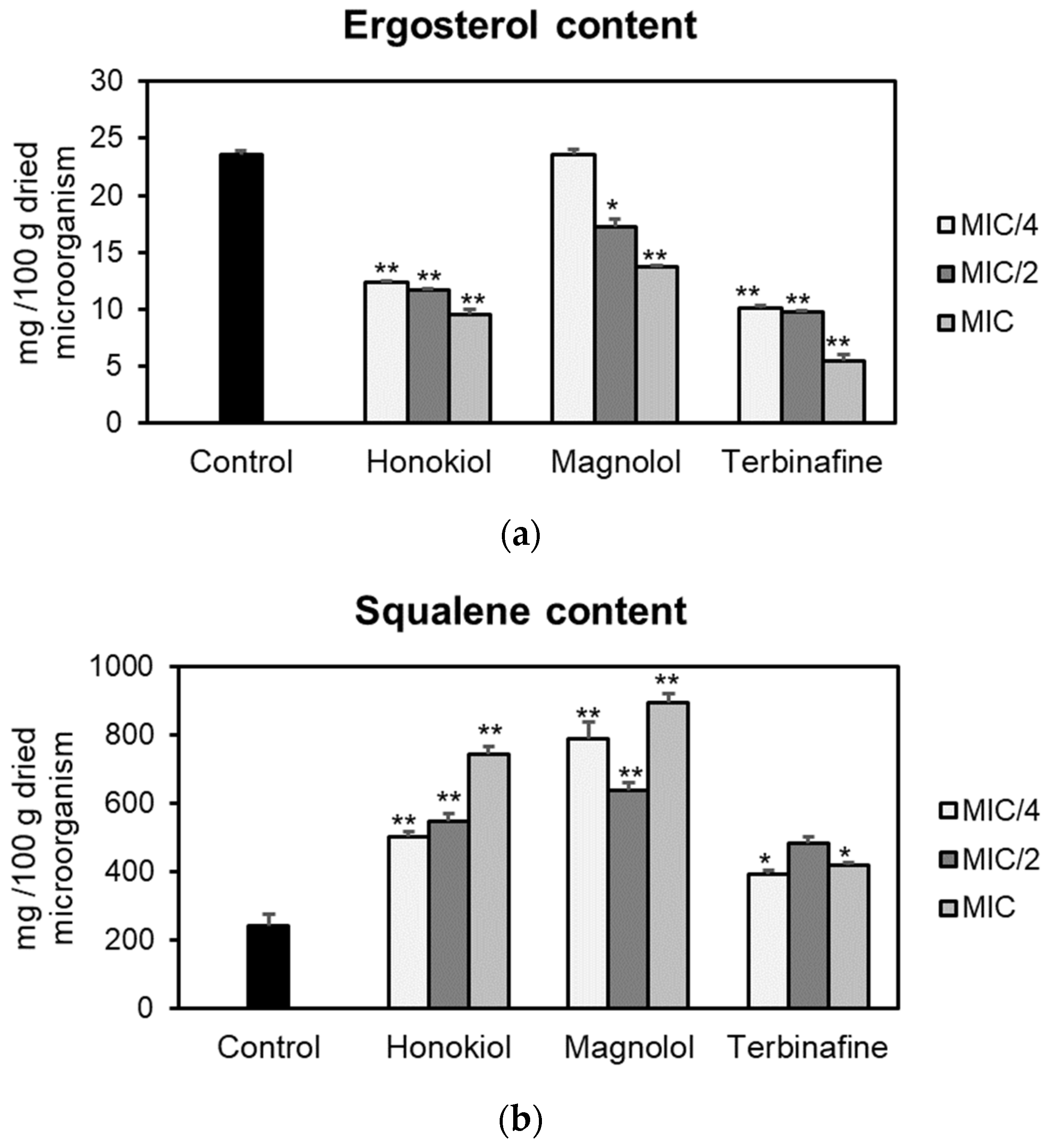
2.2. Effects on Ergosterol Biosynthesis
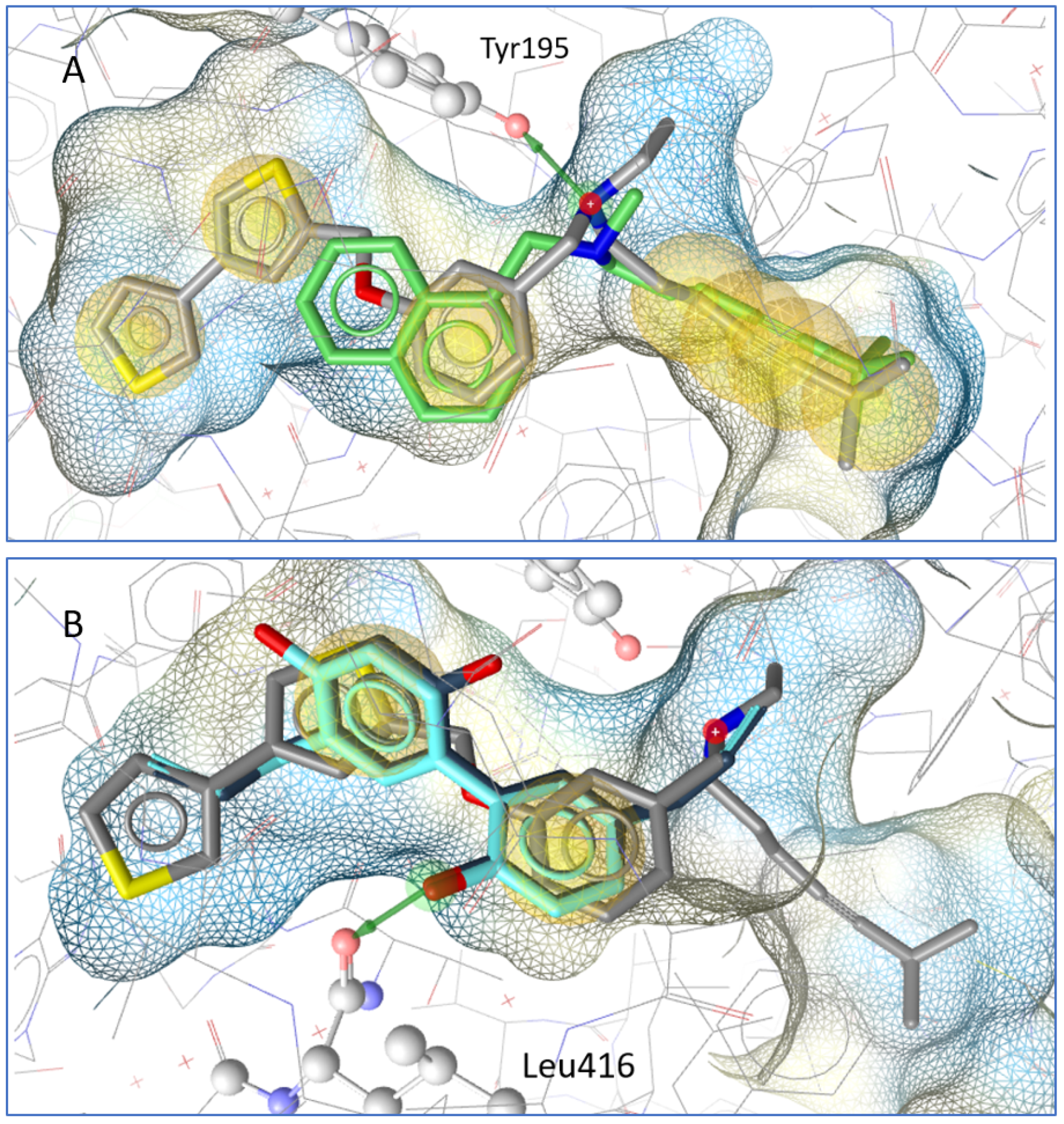
2.3. Molecular Docking Results
2.4. Checkerboard Assay Results
2.5. Effects on Cytokine Production in Human Neutrophils
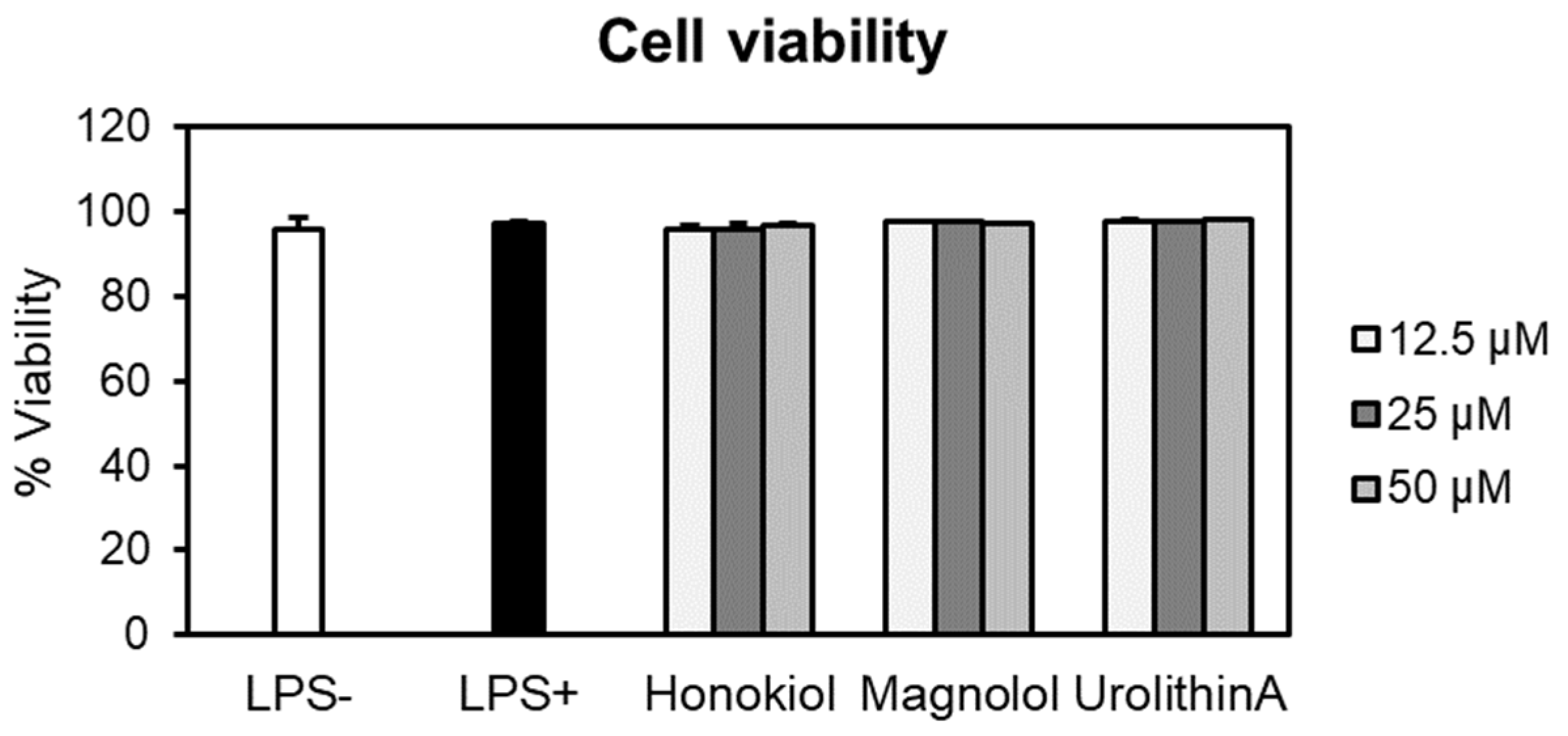
2.5.1. Effects on Neutrophils’ Viability
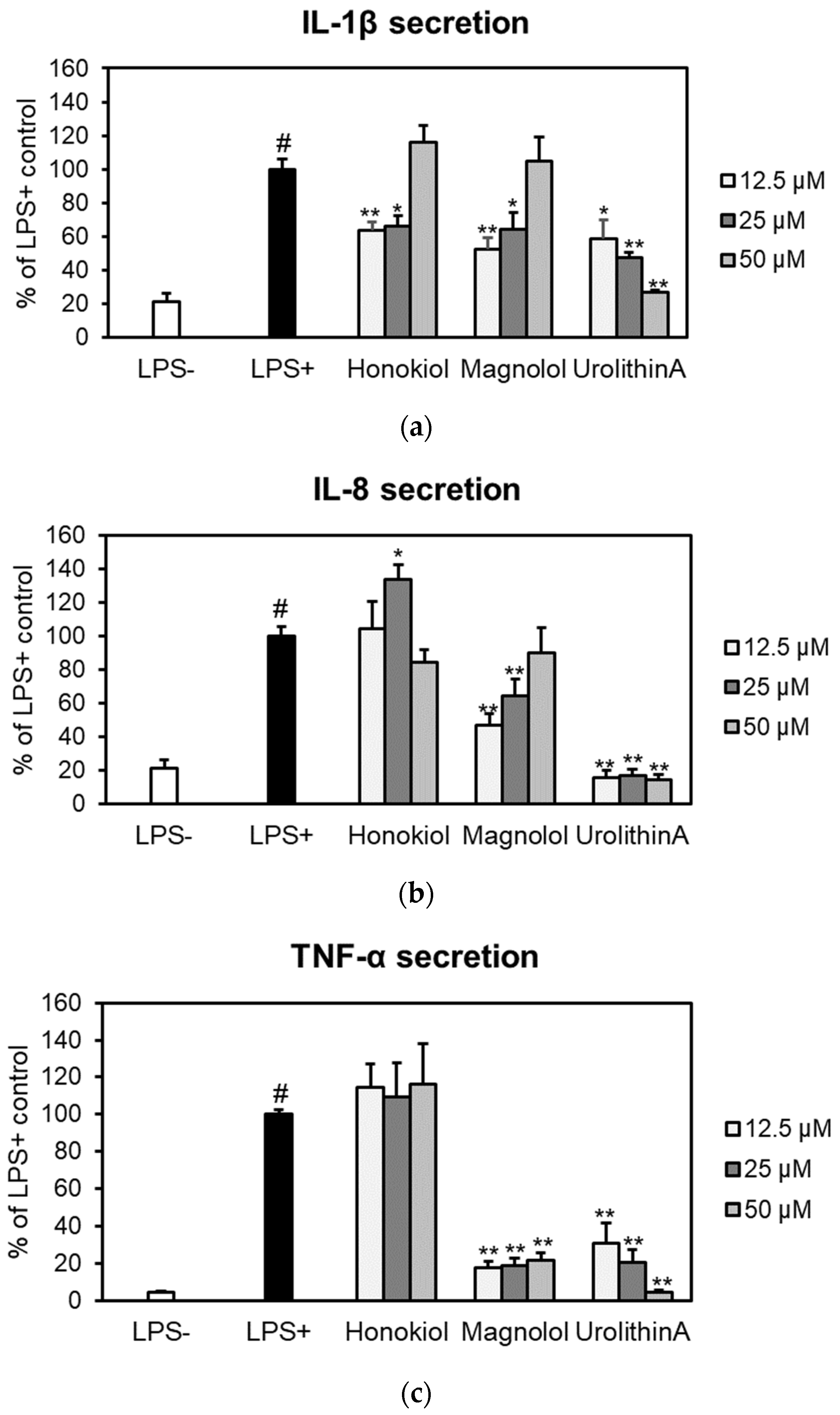
2.5.2. Effects on IL-1β, IL-8 and TNF-α Secretion
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material, Extraction
4.3. Isolation of Honokiol and Magnolol
4.4. Microbial Strains
4.5. Antifungal Susceptibility Testing
4.6. Evaluation of Effects on Ergosterol Biosynthesis
4.6.1. Sterols’ Extraction
4.6.2. HPLC-DAD Quantification of Ergosterol and Squalene
4.7. Molecular Docking Studies
4.8. Checkerboard Assay
4.9. Evaluation of Cytokine Production in Human Neutrophils
4.9.1. Neutrophils’ Isolation
4.9.2. Evaluation of Neutrophils’ Viability
4.9.3. Evaluation of IL-1β, IL-8 and TNF-α Secretion
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhan, P.; Liang, G.; Liu, W. Dermatophytes and dermatophytic infections worldwide. In Dermatophytes and Dermatophytoses; Bouchara, J.P., Nenoff, P., Gupta, A.K., Chaturvedi, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 15–40. [Google Scholar]
- Burstein, V.L.; Beccacece, I.; Guasconi, L.; Mena, C.J.; Cervi, L.; Chiapello, L.S. Skin immunity to dermatophytes: From experimental infection models to human disease. Front. Immunol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a novel multilocus phylogenetic taxonomy for the dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnat, S.; Łagowski, D.; Nowakiewicz, A. Major challenges and perspectives in the diagnostics and treatment of dermatophyte infections. J. Appl. Microbiol. 2020, 129, 212–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monod, M.; Feuermann, M.; Yamada, T. Terbinafine and itraconazole resistance in dermatophytes. In Dermatophytes and Dermatophytoses; Bouchara, J.P., Nenoff, P., Gupta, A.K., Chaturvedi, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 415–429. [Google Scholar]
- Khurana, A.; Sardana, K.; Chowdhary, A. Antifungal resistance in dermatophytes: Recent trends and therapeutic implications. Fungal Genet. Biol. 2019, 132, 103255. [Google Scholar] [CrossRef]
- Zuzarte, M.; Lopes, G.; Pinto, E.; Salgueiro, L. Are natural products an alternative therapy for dermatophytosis? In Dermatophytes and Dermatophytoses; Bouchara, J.P., Nenoff, P., Gupta, A.K., Chaturvedi, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 473–519. [Google Scholar]
- Lopes, G.; Pinto, E.; Salgueiro, L. Natural products: An alternative to conventional therapy for dermatophytosis? Mycopathologia 2017, 182, 143–167. [Google Scholar] [CrossRef]
- Luo, H.; Wu, H.; Yu, X.; Zhang, X.; Lu, Y.; Fan, J.; Tang, L.; Wang, Z. A review of the phytochemistry and pharmacological activities of Magnoliae officinalis cortex. J. Ethnopharmacol. 2019, 236, 412–442. [Google Scholar] [CrossRef]
- Lin, H.-L.; Cheng, W.-T.; Chen, L.-C.; Ho, H.-O.; Lin, S.-Y.; Hsieh, C.-M. Honokiol/magnolol-loaded self-assembling lecithin-based mixed polymeric micelles (lbMPMs) for improving solubility to enhance oral bioavailability. Int. J. Nanomed. 2021, 16, 651–665. [Google Scholar] [CrossRef]
- Lin, Y.; Li, Y.; Zeng, Y.; Tian, B.; Qu, X.; Yuan, Q.; Song, Y. Pharmacology, toxicity, bioavailability, and formulation of magnolol: An update. Front. Pharmacol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Rauf, A.; Olatunde, A.; Imran, M.; Alhumaydhi, F.A.; Aljohani, A.S.; Khan, S.A.; Uddin, M.S.; Mitra, S.; Emran, T.B.; Khayrullin, M. Honokiol: A review of its pharmacological potential and therapeutic insights. Phytomedicine 2021, 90, 153647. [Google Scholar] [CrossRef]
- Ong, C.P.; Lee, W.L.; Tang, Y.-Q.; Yap, W.H. Honokiol: A review of its anticancer potential and mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and toxicology of magnolol and honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara-Lora, I.; Bras, G.; Karkowska-Kuleta, J.; González-González, M.; Ceballos, K.; Sidlo, W.; Rapala-Kozik, M. Plant-derived substances in the fight against infections caused by Candida species. Int. J. Mol. Sci. 2020, 21, 6131. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Fatima, Z.; Hameed, S. Plant phenolics for overcoming multidrug resistance in human fungal pathogen. In Plant Phenolics in Sustainable Agriculture; Lone, R., Shuab, R., Kamili, A.N., Eds.; Springer: Singapore, 2020; pp. 407–430. [Google Scholar]
- Oufensou, S.; Scherm, B.; Pani, G.; Balmas, V.; Fabbri, D.; Dettori, M.A.; Carta, P.; Malbrán, I.; Migheli, Q.; Delogu, G. Honokiol, magnolol and its monoacetyl derivative show strong anti-fungal effect on Fusarium isolates of clinical relevance. PLoS ONE 2019, 14, e0221249. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Lu, M.-H.; Guo, D.-S.; Zhai, Y.-Y.; Miao, D.; Yue, J.-Y.; Yuan, C.-H.; Zhao, M.-M.; An, D.-R. Antifungal effect of magnolol and honokiol from Magnolia officinalis on Alternaria alternata causing tobacco brown spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-S.; Lee, J.-Y.; Park, J.; Hwang, W.; Lee, J.; Park, D. Synthesis and microbiological evaluation of honokiol derivatives as new antimicrobial agents. Arch. Pharmacal Res. 2010, 33, 61–65. [Google Scholar] [CrossRef]
- Bang, K.H.; Kim, Y.K.; Min, B.S.; Na, M.K.; Rhee, Y.H.; Lee, J.P.; Bae, K.H. Antifungal activity of magnolol and honokiol. Arch. Pharmacal Res. 2000, 23, 46–49. [Google Scholar] [CrossRef]
- Lopes, A.I.R.; Tavaria, F.; Pintado, M.E. Conventional and natural compounds for the treatment of dermatophytosis. Med. Mycol. 2019, 58, 707–720. [Google Scholar] [CrossRef]
- D’Agostino, M.; Tesse, N.; Frippiat, J.P.; Machouart, M.; Debourgogne, A.; Agostino, D. Essential oils and their natural active compounds presenting antifungal properties. Molecules 2019, 24, 3713. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Venkataraman, M.; Quinlan, E.M. New antifungal agents and new formulations against dermatophytes. In Dermatophytes and Dermatophytoses; Bouchara, J.P., Nenoff, P., Gupta, A.K., Chaturvedi, V., Eds.; Springer: Cham, Switzerland, 2021; pp. 433–471. [Google Scholar]
- Darkes, M.J.; Scott, L.J.; Goa, K.L. Terbinafine. Am. J. Clin. Dermatol. 2003, 4, 39–65. [Google Scholar] [CrossRef]
- Ryder, N. Terbinafine: Mode of action and properties of the squalene epoxidase inhibition. Br. J. Dermatol. 1992, 126, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Ryder, N.S. Specific inhibition of fungal sterol biosynthesis by SF 86-327, a new allylamine antimycotic agent. Antimicrob. Agents Chemother. 1985, 27, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brescini, L.; Fioriti, S.; Morroni, G.; Barchiesi, F. Antifungal combinations in dermatophytes. J. Fungi 2021, 7, 727. [Google Scholar] [CrossRef] [PubMed]
- Cuenca-Estrella, M. Combinations of antifungal agents in therapy–what value are they? J. Antimicrob. Chemother. 2004, 54, 854–869. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.; Lopes, L.; Rossi, A. Dermatophyte resistance to antifungal drugs: Mechanisms and prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.; da Silva, L.G.; Rossi, A.; Sanches, P.R.; Souza, L.D.R.; Martinez-Rossi, N.M. Global analysis of cell wall genes revealed putative virulence factors in the dermatophyte Trichophyton rubrum. Front. Microbiol. 2019, 10, 2168. [Google Scholar] [CrossRef]
- Durdu, M.; Ilkit, M.; Tamadon, Y.; Tolooe, A.; Rafati, H.; Seyedmousavi, S. Topical and systemic antifungals in dermatology practice. Expert Rev. Clin. Pharmacol. 2017, 10, 225–237. [Google Scholar] [CrossRef]
- Kramer, O.; Albrecht, J. Clinical presentation of terbinafine-induced severe liver injury and the value of laboratory monitoring: A Critically Appraised Topic. Br. J. Dermatol. 2017, 177, 1279–1284. [Google Scholar] [CrossRef]
- Sun, L.-M.; Liao, K.; Liang, S.; Yu, P.-H.; Wang, D.-Y. Synergistic activity of magnolol with azoles and its possible antifungal mechanism against Candida albicans. J. Appl. Microbiol. 2015, 118, 826–838. [Google Scholar] [CrossRef]
- Jin, J.; Guo, N.; Zhang, J.; Ding, Y.; Tang, X.; Liang, J.; Li, L.; Deng, X.; Yu, L. The synergy of honokiol and fluconazole against clinical isolates of azole-resistant Candida albicans. Lett. Appl. Microbiol. 2010, 51, 351–357. [Google Scholar] [CrossRef]
- Calderon, R.A.; Hay, R.J. Fungicidal activity of human neutrophils and monocytes on dermatophyte fungi, Trichophyton quinckeanum and Trichophyton rubrum. Immunology 1987, 61, 289–295. [Google Scholar] [PubMed]
- van der Linden, M.; Meyaard, L. Fine-tuning neutrophil activation: Strategies and consequences. Immunol. Lett. 2016, 178, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.; Fleckman, P.; Olerud, J.E. Biofilms and inflammation in chronic wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef]
- Sychrová, A.; Koláriková, I.; Žemlička, M.; Šmejkal, K. Natural compounds with dual antimicrobial and anti-inflammatory effects. Phytochem. Rev. 2020, 19, 1471–1502. [Google Scholar] [CrossRef]
- Wang, X.-D.; Wang, Y.-L.; Gao, W.-F. Honokiol possesses potential anti-inflammatory effects on rheumatoid arthritis and GM-CSF can be a target for its treatment. Int. J. Clin. Exp. Pathol. 2015, 8, 7929–7936. [Google Scholar]
- Li, C.-Y.; Chao, L.K.; Wang, S.-C.; Chang, H.-Z.; Tsai, M.-L.; Fang, S.-H.; Liao, P.-C.; Ho, C.-L.; Chen, S.-T.; Cheng, W.-C.; et al. Honokiol inhibits LPS-induced maturation and inflammatory response of human monocyte-derived dendritic cells. J. Cell. Physiol. 2011, 226, 2338–2349. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-M.; Chen, S.-H.; Liao, Y.-W.; Yu, C.-H.; Yu, C.-C.; Hsieh, P.-L. Magnolol ameliorates the accumulation of reactive oxidative stress and inflammation in diabetic periodontitis. J. Formos. Med. Assoc. 2021, 120, 1452–1458. [Google Scholar] [CrossRef]
- Song, Z.; Sun, J.; Deng, P.; Zhou, F.; Xu, H.; Wen, Y.; Teng, F.; Ge, D.; Feng, R. Oligochitosan-pluronic 127 conjugate for delivery of honokiol. Artif. Cells Nanomed. Biotechnol. 2018, 46, 740–750. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Deng, L.; Cai, L.; Zhang, X.; Zheng, H.; Deng, C.; Duan, X.; Zhao, X.; Wei, Y.; Chen, L. Preparation, characterization, pharmacokinetics, and bioactivity of honokiol-in-hydroxypropyl-β-cyclodextrin-in-liposome. J. Pharm. Sci. 2011, 100, 3357–3364. [Google Scholar] [CrossRef]
- Arendrup, M.; Meletiadis, J.; Mouton, J.; Lagrou, K.; Hamal, P.; Guinea, J. EUCAST Definitive Document E. DEF. 9.3.1. Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. Lond. Eur. Comm. Antimicrob. Susceptibility Test. 2017, 7, 1. [Google Scholar]
- Saunte, D.M.L.; Hare, R.K.; Jørgensen, K.M.; Jørgensen, R.; Deleuran, M.; Zachariae, C.O.; Thomsen, S.F.; Bjørnskov-Halkier, L.; Kofoed, K.; Arendrup, M.C. Emerging terbinafine resistance in Trichophyton: Clinical characteristics, squalene epoxidase gene mutations, and a reliable EUCAST method for detection. Antimicrob. Agents Chemother. 2019, 63, e01126-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markantonatou, A.-M.; Samaras, K.; Zachrou, E.; Vyzantiadis, T.-A. Comparison of four methods for the in vitro susceptibility testing of dermatophytes. Front. Microbiol. 2020, 11, 1593. [Google Scholar] [CrossRef]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of ergosterol content: Novel method for determination of fluconazole susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.; Pinto, E.; Andrade, P.; Valentão, P. Antifungal activity of phlorotannins against dermatophytes and yeasts: Approaches to the mechanism of action and influence on Candida albicans virulence factor. PLoS ONE 2013, 8, e72203. [Google Scholar] [CrossRef]
- Padyana, A.K.; Gross, S.; Jin, L.; Cianchetta, G.; Narayanaswamy, R.; Wang, F.; Wang, R.; Fang, C.; Lv, X.; Biller, S.A.; et al. Structure and inhibition mechanism of the catalytic domain of human squalene epoxidase. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, P. Methods for determining bactericidal activity and antimicrobial interactions: Synergy testing, time-kill curves, and population analysis. In Antimicrobial Susceptibility Testing Protocols; Schwalbe, R., Steele-Moore, L., Goodwin, A.C., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 275–298. [Google Scholar]
- van Vuuren, S.; Viljoen, A. Plant-based antimicrobial studies–methods and approaches to study the interaction between natural products. Planta Med. 2011, 77, 1168–1182. [Google Scholar] [CrossRef] [Green Version]
- Trifan, A.; Luca, S.V.; Greige-Gerges, H.; Miron, A.; Gille, E.; Aprotosoaie, A.C. Recent advances in tackling microbial multidrug resistance with essential oils: Combinatorial and nano-based strategies. Crit. Rev. Microbiol. 2020, 46, 1–20. [Google Scholar] [CrossRef]
- Czerwińska, M.; Dudek, M.K.; Pawłowska, K.; Pruś, A.; Ziaja, M.; Granica, S. The influence of procyanidins isolated from small-leaved lime flowers (Tilia cordata Mill.) on human neutrophils. Fitoterapia 2018, 127, 115–122. [Google Scholar] [CrossRef]
- Trifan, A.; Skalicka-Woźniak, K.; Granica, S.; Czerwińska, M.E.; Kruk, A.; Marcourt, L.; Wolfender, J.-L.; Wolfram, E.; Esslinger, N.; Grubelnik, A.; et al. Symphytum officinale L.: Liquid-liquid chromatography isolation of caffeic acid oligomers and evaluation of their influence on pro-inflammatory cytokine release in LPS-stimulated neutrophils. J. Ethnopharmacol. 2020, 262, 113169. [Google Scholar] [CrossRef]
- Rønning, S.B.; Voldvik, V.; Bergum, S.K.; Aaby, K.; Borge, G.I.A. Ellagic acid and urolithin A modulate the immune response in LPS-stimulated U937 monocytic cells and THP-1 differentiated macrophages. Food Funct. 2020, 11, 7946–7959. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Gong, L.F.; Wu, Y.F.; Lin, Z.; Jiang, B.J.; Wu, L.; Yu, K.H. Urolithin A targets the PI3K/Akt/NF-κB pathways and prevents IL-1β-induced inflammatory response in human osteoarthritis: in vitro and in vivo studies. Food Funct. 2019, 10, 6135–6146. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Honokiol | Magnolol | Terbinafine | |||
|---|---|---|---|---|---|---|
| MIC | MFC | MIC | MFC | MIC | MFC | |
| Trichophyton rubrum ATCC 28188 | 8 | 16 | 8 | 16 | 0.031 | 0.031 |
| Trichophyton mentagrophytes ATCC 9533 | 8 | 16 | 8 | 16 | 0.031 | 0.031 |
| Trichophyton rubrum RTCC 1348 * | 8 | 16 | 8 | 16 | 0.031 | 0.031 |
| Trichophyton rubrum RTCC 2158 * | 8 | 16 | 8 | 16 | 0.062 | 0.062 |
| Trichophyton ajelloi RTCC 1356 * | 8 | 16 | 8 | 16 | 0.062 | 0.062 |
| Microsporum gypseum RTCC 2172 * | 8 | 16 | 8 | 16 | 0.062 | 0.062 |
| Microsporum canis RTCC 1883 * | 8 | 16 | 8 | 16 | 0.062 | 0.062 |
| Combination | MIC * in combination (mg/L) | FIC | FICI | Interpretation |
|---|---|---|---|---|
| Honokiol | 0.5 | 0.06 | 0.56 | Add |
| Terbinafine | 0.015 | 0.50 | ||
| Magnolol | 2 | 0.25 | 0.50 | Syn |
| Terbinafine | 0.007 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifan, A.; Bostănaru, A.-C.; Luca, S.V.; Temml, V.; Akram, M.; Herdlinger, S.; Kulinowski, Ł.; Skalicka-Woźniak, K.; Granica, S.; Czerwińska, M.E.; et al. Honokiol and Magnolol: Insights into Their Antidermatophytic Effects. Plants 2021, 10, 2522. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112522
Trifan A, Bostănaru A-C, Luca SV, Temml V, Akram M, Herdlinger S, Kulinowski Ł, Skalicka-Woźniak K, Granica S, Czerwińska ME, et al. Honokiol and Magnolol: Insights into Their Antidermatophytic Effects. Plants. 2021; 10(11):2522. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112522
Chicago/Turabian StyleTrifan, Adriana, Andra-Cristina Bostănaru, Simon Vlad Luca, Veronika Temml, Muhammad Akram, Sonja Herdlinger, Łukasz Kulinowski, Krystyna Skalicka-Woźniak, Sebastian Granica, Monika E. Czerwińska, and et al. 2021. "Honokiol and Magnolol: Insights into Their Antidermatophytic Effects" Plants 10, no. 11: 2522. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112522