
Phytogeographical Analysis and Ecological Factors of the Distribution of Orchidaceae Taxa in the Western Carpathians (Local study)


Abstract
:1. Introduction
2. Materials and Methods
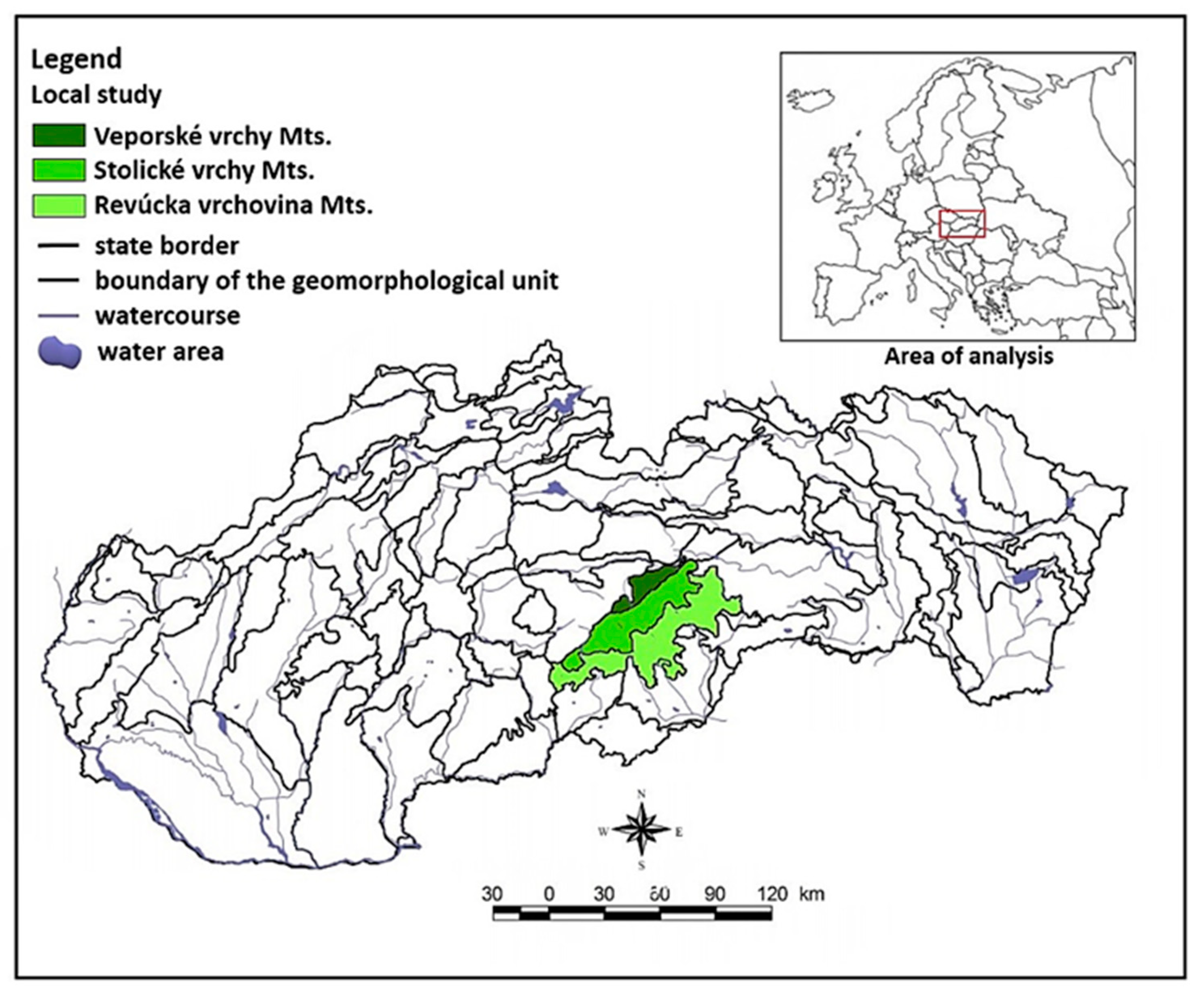
2.1. Study Area
2.2. Geographical Conditions
2.3. Landscape Ecology and Landscape Cover Analysis
- anthropogenic - intensification of agriculture; forestry management; drainage; land use for technical construction; harvesting for gardening and medical purposes; mineral extraction; application of fertilizers; application of herbicide;, large-scale application of insecticides; soil compaction; mulching—covering of the soil surface
- zoogenic - damage done by large ungulates, small subterrestrial mammals and insects
- phytogenic - displacement by non-native (invasive) plant species, succession—plant association change
- pedogenic - soil erosion, high pH, nitrogen and ion content in soil.
2.4. Data Analyses
3. Results
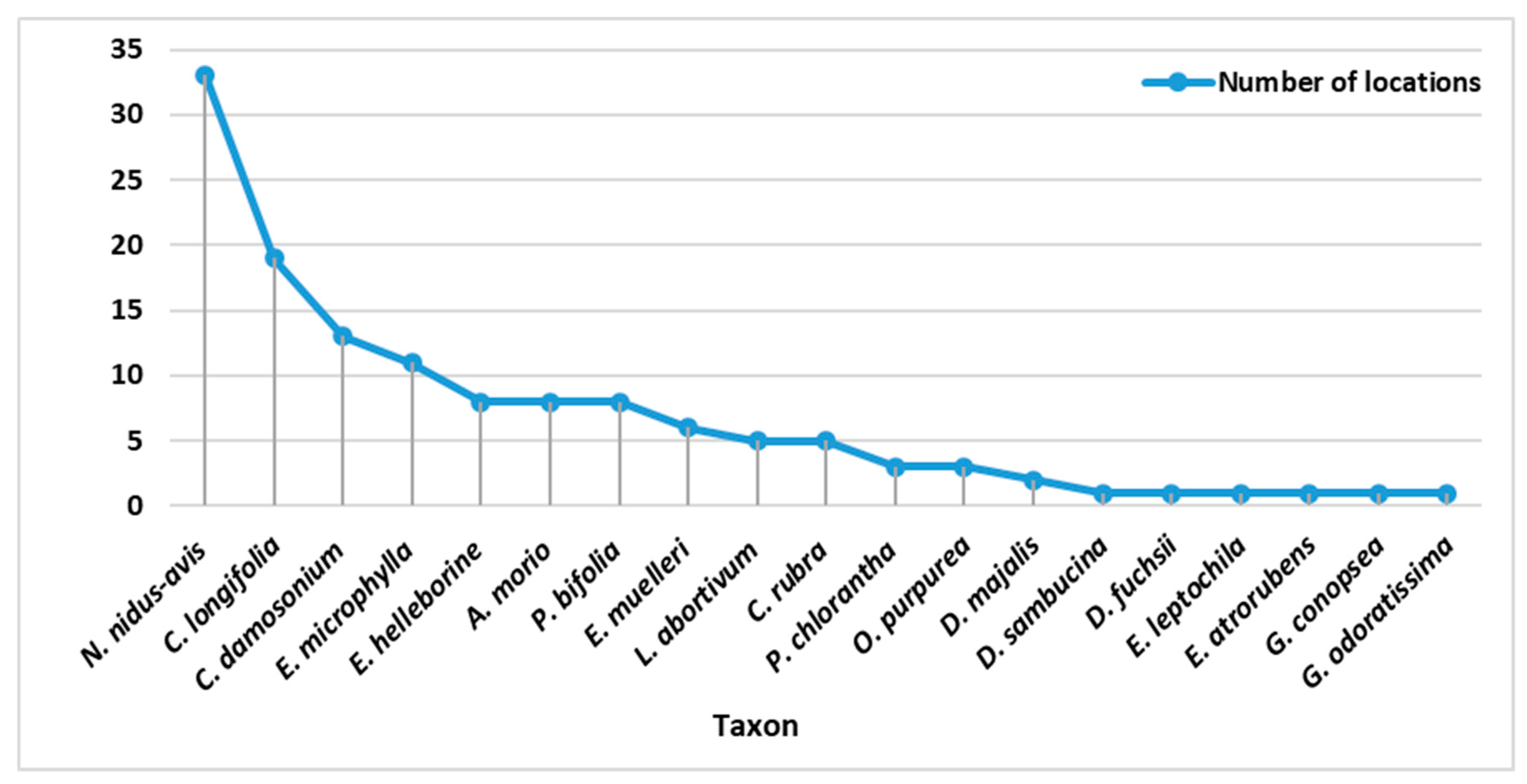
3.1. Monitored Taxa
- Cephalanthera damosonium (Mill.) Druce, NT / §
- Cephalanthera longifolia (L.) Fritsch, NT / §
- Cephalanthera rubra (L.) Rich., NT / §
- Anacamptis morio (L.) R. M. Bateman, NT / §
- Dactylorhiza majalis subsp. majalis (Rchb.) P. F. Hunt et Summerh, NT / §
- Dactylorhiza sambucina (L.) Soó, NT/ §
- Orchis purpurea Huds., NT / §
- Dactylorhiza fuchsii subsp. fuchsii (Druce) Soó, NT / §
- Platanthera bifolia subsp. latiflora (L.) Rich., LC
- Platanthera chlorantha (Custer) Rchb., NT / §
- Epipactis helleborine subsp. helleborine (L.) Crantz, LC
- Epipactis atrorubens (Hoffm.) Besser, LC / §
- Epipactis microphylla (Ehrh.) Sw., LC / §
- Epipactis muelleri Godfery, NT / §
- Epipactis leptochila Godfery, VU / §
- Limodorum abortivum (L.) Sw. NT / §
- Neottia nidus-avis (L.) L. C. M. Richard
- Gymnadenia conopsea (L.) R. Brown., LC / §
- Gymnadenia odoratissima (L.) Rich., NT / §
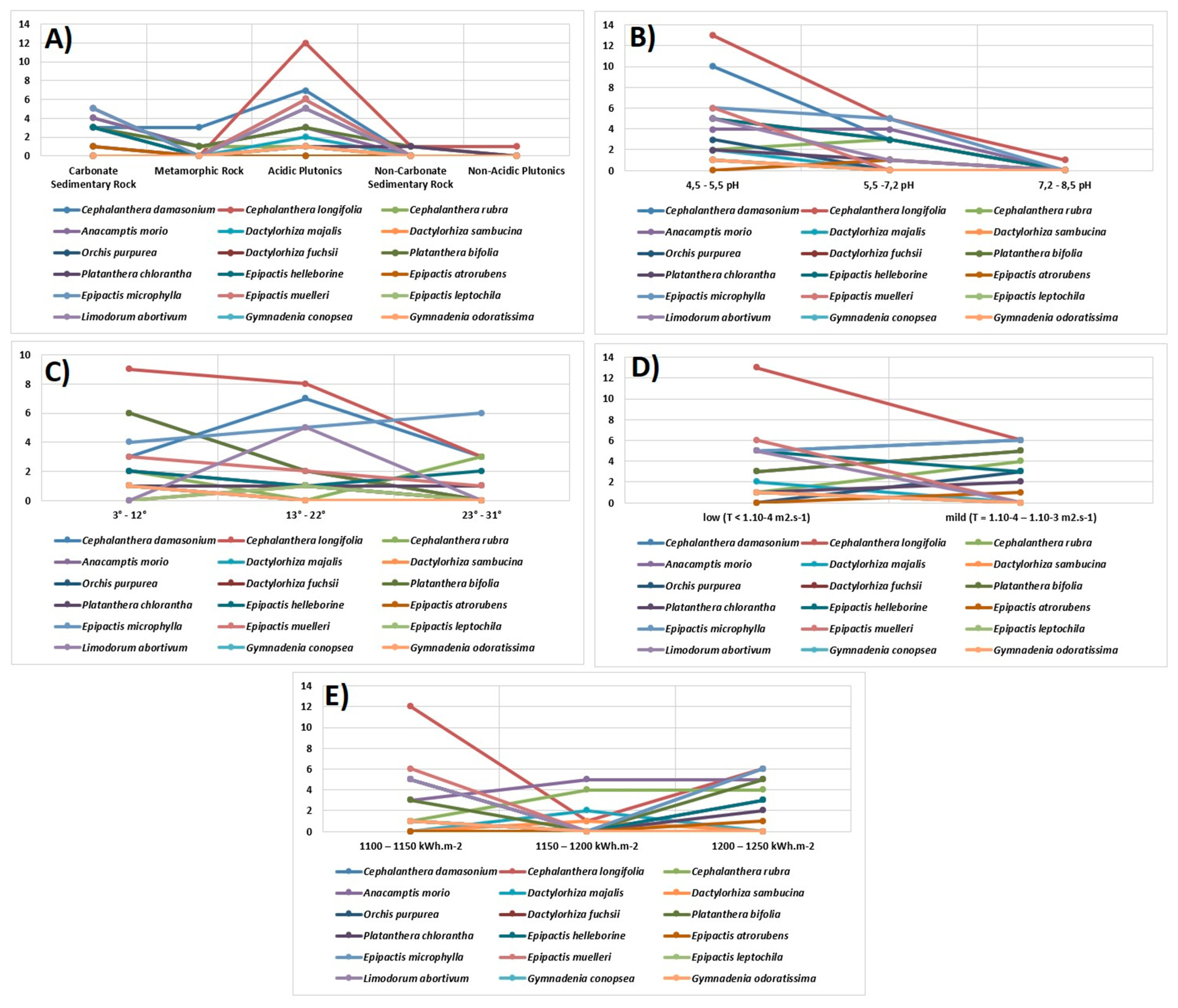
3.2. Ecological Factors and Diversity of Orchids
4. Discussion
Biogeography and Conservation Biology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eliáš, P.; Dítě, D.; Kliment, J.; Hrivnák, R.; Feráková, V. Red list of ferns and floweing plants of Slovakia. 5th edition. Biologia 2015, 70, 218–228. [Google Scholar] [CrossRef]
- Zákon, Č. 543/2002 z. z. o ochrane prírody a krajiny. Nature and Landscape Protection Act. 2002. Available online: http://www.sopsr.sk/news/file/ZZ_2002_543_20210101.pdf (accessed on 12 February 2021).
- Jasík, M. Legislatívna ochrana vstavačovitých v Slovenskej republike. In Európske Vstavačovité (Orchidaceae)—Výskum a Ochrana; Technická univerzita: Zvolen, Slovak, 1997; pp. 27–31. [Google Scholar]
- Prochádzka, F.; Velísek, V. Orchideje Naší Přírody; Academia: Prague, Czech Republic, 1983; p. 284. [Google Scholar]
- Ujházy, K.; Máliš, F.; Križová, E.; Nič, J. Fytocenológia a lesnícka typológia, ekológia (návody na cvičenia). TU in Zvolen. 2018. 119 S. Available online: https://kf.tuzvo.sk/sites/default/files/ujhazy_et-al_2018_navody_na_cvicenia_prac-verz.pdf (accessed on 14 February 2021).
- Hrivnák, M.; Slezák, M.; Galvánek, D.; Vlčko, J.; Belanová, E.; Rízová, V.; Senko, D.; Hrivnák, R. Species Richness, Ecology, and Prediction of Orchids in Central Europe: Local-Scale Study. Diversity 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Bateman, M.R. Evolutionary classification of European orchids: The crucial importance of maximising explicit evidence and minimising authoritarian speculation. J. Eur. Orch. 2009, 41, 243–318. [Google Scholar]
- Gaskett, C.A.; Gallagher, V.R. Orchid diversity: Spatial and climatic patterns from herbarium records. Ecol. Evol. 2018, 8, 11235–11245. [Google Scholar] [CrossRef] [Green Version]
- Kreutz, C.A.J.K. Kompendium der Europäischen Orchideen/Catalogue of European Orchids, 1st ed.; Kreutz Publishers: Landgraa, The Netherland, 2004; p. 240. ISBN 90-806626-4-X. [Google Scholar]
- Štípková, Z.; Romportl, D.; Kindlmann, P. Which Environmental Factors Drive Distribution of Orchids? A Case Study from South Bohemia, Czech Republic. In Orchids Phytochemistry, Biology and Horticulture; Springer: Cham, Switzerland, 2020; pp. 1–33. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 14, 355–370. [Google Scholar] [CrossRef]
- Slaviero, A.; Del Vecchio, S.; Pierce, S.; Fantinato, E.; Buffa, G. Plant community attributes affect dry grassland orchid establishment. Plant Ecol. 2016, 217, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Pollination mechanisms are driving orchid distribution in space. Sci. Rep. 2020, 10, 850. [Google Scholar] [CrossRef] [Green Version]
- Tsiftsis, S.; Štípková, Z.; Kindlmann, P. Role of way of life, latitude, elevation and climate on the richness and distribution of orchid species. Biodivers. Conserv. 2019, 28, 75–96. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I. Temporal and spatial patterns of orchid species distribution in Greece: Implications for conservation. Biodivers. Conserv. 2020, 29, 3461–3489. [Google Scholar] [CrossRef]
- Perazza, G.; Decarli, M. Monitoring of Cypripedium calceolus (Orchidaceae) in the Adamello-Brenta Natural Park (Italy). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 178–184. [Google Scholar] [CrossRef]
- Tatarenko, I.; Dodd, M.; Wallace, H.; Bellamy, G.; Fleckney, A. Protecting small populations of rare species. Case study on Dactylorhiza viridis (Orchidaceae) in Fancott Woods and Meadows SSSI, Bedfordshire, UK. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 165–171. [Google Scholar] [CrossRef]
- Kirillova, I.A.; Kirillov, D.V. Impact of weather conditions on seasonal development, population structure and reproductive success on Dactylorhiza traunsteineri (Orchidaceae) in the Komi Republic (Russia). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 77–89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, J.; He, J.S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2020, 181, 64–72. [Google Scholar] [CrossRef]
- Kočický, D.; Ivanič, B. Geomorfologické Členenie Slovenska. 2014. Available online: https://apl.geology.sk/temapy/ (accessed on 23 March 2020).
- Plesník, P. Fytogeograficko-vegetačné členenie. M 1: 1 000 000. In Atlas Krajiny Slovenskej Republiky, Ministerstvo Životného Prostredia SR, Slovenská Agentúra Životného Prostredia; Atlas krajiny, S.R., Hrnčiarová, T., Eds.; MŽP: Bratislava, Slovakia; SAŢP: Banská Bystrica, Slovakia, 2002; ISBN 80-88833-27-2. [Google Scholar]
- Vass, D. Geologická Mapa Lučenskej Kotliny a Cerovej Vrchoviny; ŠGÚDS: Bratislava, Slovakia, 1992; ISBN 79-603-92-18/20. [Google Scholar]
- Kočický, D.; Ivanič, B. Klimatickogeografické Typy; Štátny Geologický Ústav Dionýza Štúra: Bratislava, Slovakia, 2014; Available online: http://apl.geology.sk/temapy (accessed on 18 July 2020).
- Šály, R.; Šurina, B. Pôdy. In Atlas Krajiny Slovenskej Republiky; SAŽP: Bratislava, Slovakia, 2002; pp. 106–111. [Google Scholar]
- Michalko, J.; Magic, D.; Maglocký, Š. Geobotanická mapa ČSSR, SSR, M 1: 200 000. Veda. 1986. Available online: http://www.kgrr.fpv.ukf.sk/index.php/ostatne/na-stiahnutie (accessed on 14 February 2021).
- ESRI. Corine Land Cover. 2018. Available online: https://image.discomap.eea.europa.eu/arcgis/rest/services/Corine/CLC2018_WM/MapServer (accessed on 18 July 2020).
- Wittlinger, L.; Šolcová, L. Geografická charakteristika územia so zameraním na vybrané antropogénne vplyvy ťažby. Geogr. Inf. 2018, 22, 539–551. [Google Scholar] [CrossRef]
- ESRI. Basemap. Earthstar Geographics, Esri, HERE, Garmin, FAO, NOAA, USGS. Available online: https://maps.arcgis.com (accessed on 18 July 2020).
- Wittlinger, L. Botanické Výskytové Dáta Mapovateľa. Komplexný Informačný a Monitorovací Systém—ŠOP SR. 2019. Available online: https://www.biomonitoring.sk/ (accessed on 20 July 2020).
- ESRI World Lithology. Available online: https://landscape6.arcgis.com/arcgis/rest/services/World_Lithology/ImageServer (accessed on 20 July 2020).
- ESRI. World Soils Harmonized World Soil Database—Chemistry. Available online: https://landscape6.arcgis.com/arcgis/rest/services/World_Soils_HWSD_Chemistry/ImageServer (accessed on 20 July 2020).
- ESRI. Terrain: Slope Map. Available online: https://elevation.arcgis.com/arcgis/rest/services/WorldElevation/Terrain/ImageServer (accessed on 20 July 2020).
- Tomlain, J.; Hrvoľ, J.; Globálne Žiarenie a Relatívne Trvanie Slnečného Svitu. Atlas Krajiny Slovenskej Republiky. 2002. Available online: https://app.sazp.sk/atlassr/ (accessed on 18 July 2020).
- Malík, P.; Švasta, J.; Jetel, J.; Hanzel, V.; Gedeon, M.; Scherer, S.; Fendek, M. Hydrogeologické Pomery. Atlas Krajiny Slovenskej Republiky. 2002. Available online: https://app.sazp.sk/atlassr/ (accessed on 18 July 2020).
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, W.; Werner, W.; Pauliben, D. Zeigen von Pflanzen in Mitteleuropa. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- Stanová, V.; Valachovič, M. Katalóg Biotopov Slovenska; DAPHNE—Inštitút Aplikovanej Ekológie: Bratislava, Slovakia, 2002; p. 225. [Google Scholar]
- Dykyjová, D. Ekologie Středoevropských Orchidejí, Kopp, Jihočeská Univerzita; Kopp, 2003 České Budějovice: České Budějovice, Czech Republic, 2003; p. 115. [Google Scholar]
- Vlčko, J.; Dítě, D.; Kolník, M. Vstavačovité Slovenska; ZO SZOPK Orchidea: Zvolen, Slovakia, 2003; pp. 1–120. [Google Scholar]
- Ujházy, K.; Vlčko, J.; Uhliarová, E. Nová lokalita vstavačovitých (Orchis coriophora, O. ustulata, O. morio) na severozápadnom úpätí Poľany. Bull. Slov. Bot. Spoločn. Bratisl. 2003, 25, 61–68. [Google Scholar]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Communities: Luxembourg, 2011; p. 130. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. 2020. Available online: https://www.gbif.org/citation-guidelines (accessed on 15 January 2021).
- Molnár, V.A. Magyarország Orchideáinak Atlasza; Kossuth Kiadó: Budapest, Hungary, 2011; pp. 1–510. [Google Scholar]
- Kirca, S.; Kreutz, K.C.A.J.; Çolak, A.H. A biogeographical and ecological classification of orchids in Turkey. Phytocoenologia 2020, 50, 65–77. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S. Patterns of orchid species richness and composition in relation to geological substrates. Wulfenia 2019, 26, 1–21. Available online: http://biore.bio.bg.ac.rs/handle/123456789/3546 (accessed on 8 March 2021).
- Tsiftsis, S.; Tsiripidis, I.; Karagiannakidou, V.; Alifragis, D. Niche analysis and conservation of the orchids of east Macedonia (NE Greece). Acta Oecol. 2018, 33, 27–35. [Google Scholar] [CrossRef]
- Shevchyk, V.L.; Solomakha, I.; Dvirna, S.T. A new finding of a uniquely large population of Anacamptis morio (Orchidaceae) on the Left-Bank Ukraine. Biodivers. Res. Conserv. 2019, 55, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Kirby, S. Geological processes and orchid biogeography with applications to southeast Central America. Lankesteriana 2015, 7, 53–55. [Google Scholar] [CrossRef] [Green Version]
- Rocha, F.S.; Waechter, J.L. Ecological distribution of terrestrial orchids in a south Brazilian Atlantic region. Nord. J. Bot. 2009, 28, 112–118. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Localities Code | Abiotic Environmental Factors | ||||
|---|---|---|---|---|---|---|
| Substrate: Ecological Land Unit Value | Soil Reaction (pH) | Terrain: Slope (°) | Flow and Hydrogeological Productivity (m2.s−1) | Average Annual Amounts of Global Radiation (kWh.m–2) | ||
| Cephalanthera damasonium | CD_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10−4–1.10-3) | 1100–1150 |
| CD_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CD_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CD_04 | Metamorphic Rock | 4.5–5.5 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CD_05 | Metamorphic Rock | 4.5–5.5 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CD_06 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CD_07 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| CD_08 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| CD_09 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| CD_10 | Metamorphic Rock | 4.5–5.5 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CD_11 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| CD_12 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CD_13 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| Cephalanthera longifolia | CL_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| CL_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CL_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CL_04 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CL_05 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CL_06 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| CL_07 | Non-Carbonate Sedimentary Rock | 4.5–5.5 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CL_08 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_09 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| CL_10 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_11 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| CL_12 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_13 | Non-Acidic Plutonics | 7.2–8.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_14 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_15 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| CL_16 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| CL_17 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| CL_18 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1150–1200 | |
| CL_19 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| Cephalanthera rubra | CR_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| CR_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CR_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CR_04 | Metamorphic Rock | 4.5–5.5 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| CR_05 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| Anacamptis morio | AM_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| AM_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| AM_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 3 - 12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| AM_04 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| AM_05 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| AM_06 | Metamorphic Rock | 4.5–5.5 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| AM_07 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| AM_08 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| Dactylorhiza majalis | DR_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1150–1200 |
| DR_02 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1150–1200 | |
| Dactylorhiza sambucina | DS_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1150–1200 |
| Orchis purpurea | OP_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| OP_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| OP_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| Dactylorhizafuchsii | DF_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 |
| Platanthera bifolia | PB_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| PB_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| PB_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| PB_04 | Non-Carbonate Sedimentary Rock | 4.5–5.5 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| PB_05 | Metamorphic Rock | 4.5–5.5 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| PB_06 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| PB_07 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| PB_08 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| Platanthera chlorantha | PCH_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| PCH_02 | Non-Carbonate Sedimentary Rock | 4.5–5.5 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| PCH_03 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| Epipactis helleborine | EH_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| EH_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EH_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EH_04 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| EH_05 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EH_06 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| EH_07 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| EH_08 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| Epipactis atrorubens | EA_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| Epipactis microphylla | EM_01 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 |
| EM_02 | Carbonate Sedimentary Rock | 5.5–7.2 | 23–31 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EM_03 | Carbonate Sedimentary Rock | 5.5–7.2 | 3–12 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EM_04 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EM_05 | Carbonate Sedimentary Rock | 5.5–7.2 | 13–22 | mild (T = 1.10-4–1.10-3) | 1100–1150 | |
| EM_06 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EM_07 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EM_08 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EM_09 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| EM_10 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| EM_11 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| Epipactis muelleri | EMu_01 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 |
| EMu_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EMu_03 | Acidic Plutonics | 4.5–5.5 | 23–31 | low (T < 1.10-4) | 1200–1250 | |
| EMu_04 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| EMu_05 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| EMu_06 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 | |
| Epipactis leptochila | EL_01 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 |
| Limodorum abortivum | LA_01 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 |
| LA_02 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| LA_03 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| LA_04 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| LA_05 | Acidic Plutonics | 4.5–5.5 | 13–22 | low (T < 1.10-4) | 1200–1250 | |
| Gymnadenia conopsea | GC_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 |
| Gymnadenia odoratissima | GO_01 | Acidic Plutonics | 4.5–5.5 | 3–12 | low (T < 1.10-4) | 1200–1250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wittlinger, L.; Petrikovičová, L. Phytogeographical Analysis and Ecological Factors of the Distribution of Orchidaceae Taxa in the Western Carpathians (Local study). Plants 2021, 10, 588. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030588
Wittlinger L, Petrikovičová L. Phytogeographical Analysis and Ecological Factors of the Distribution of Orchidaceae Taxa in the Western Carpathians (Local study). Plants. 2021; 10(3):588. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030588
Chicago/Turabian StyleWittlinger, Lukáš, and Lucia Petrikovičová. 2021. "Phytogeographical Analysis and Ecological Factors of the Distribution of Orchidaceae Taxa in the Western Carpathians (Local study)" Plants 10, no. 3: 588. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030588