
Conservation of Micromeria browiczii (Lamiaceae), Endemic to Zakynthos Island (Ionian Islands, Greece)

 and
and
Abstract
:1. Introduction
2. Results
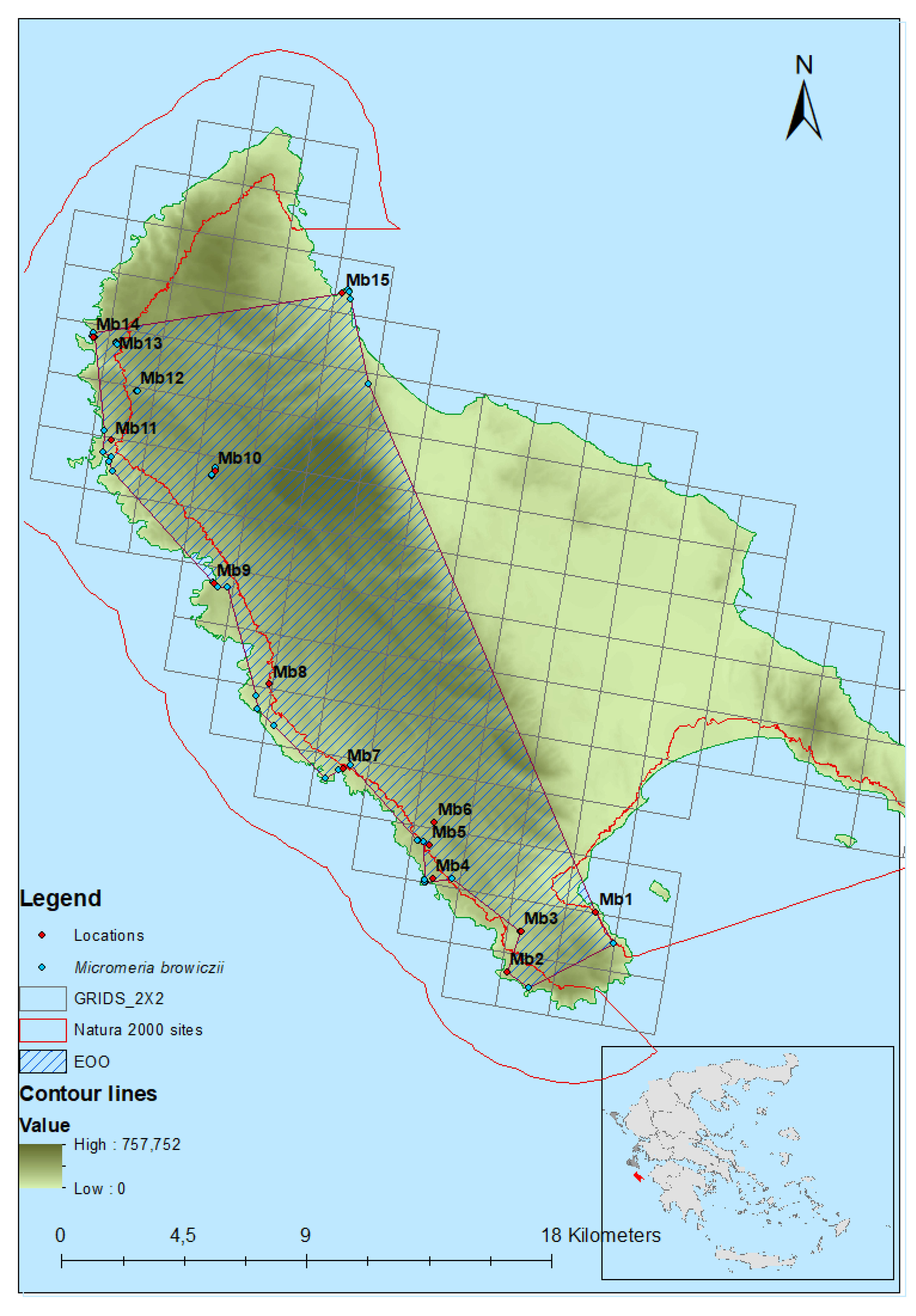
2.1. Geographical Distribution
2.2. Population Size
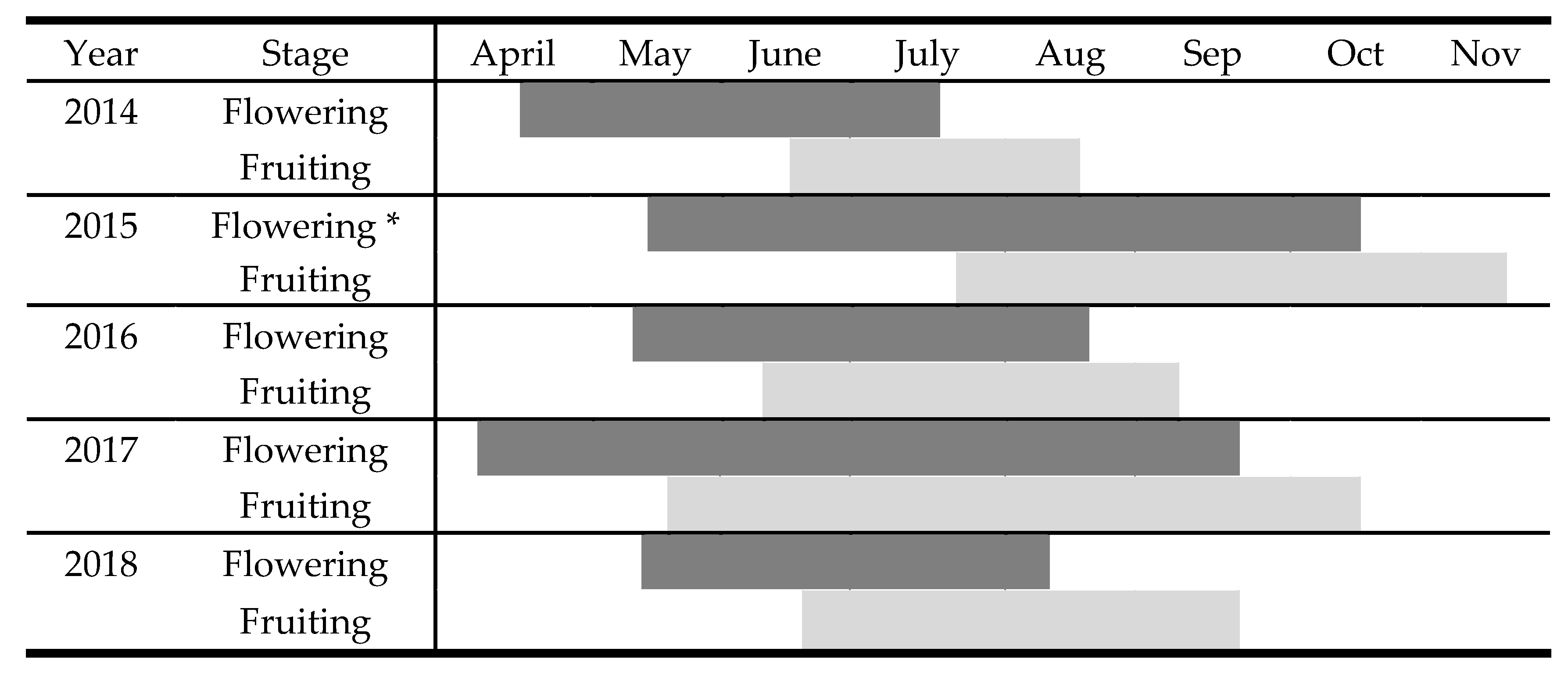
2.3. Reproductive Biology
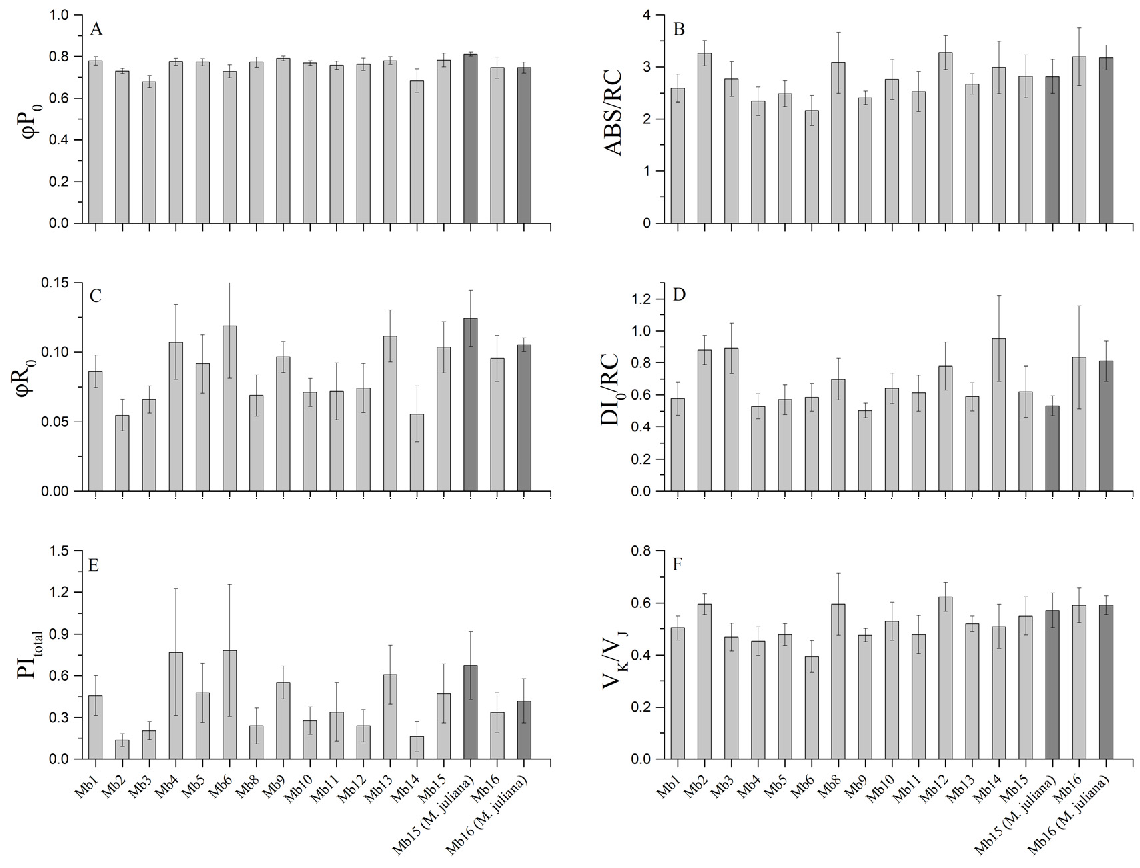
2.4. Chlorophyll Fluorescence Measurements
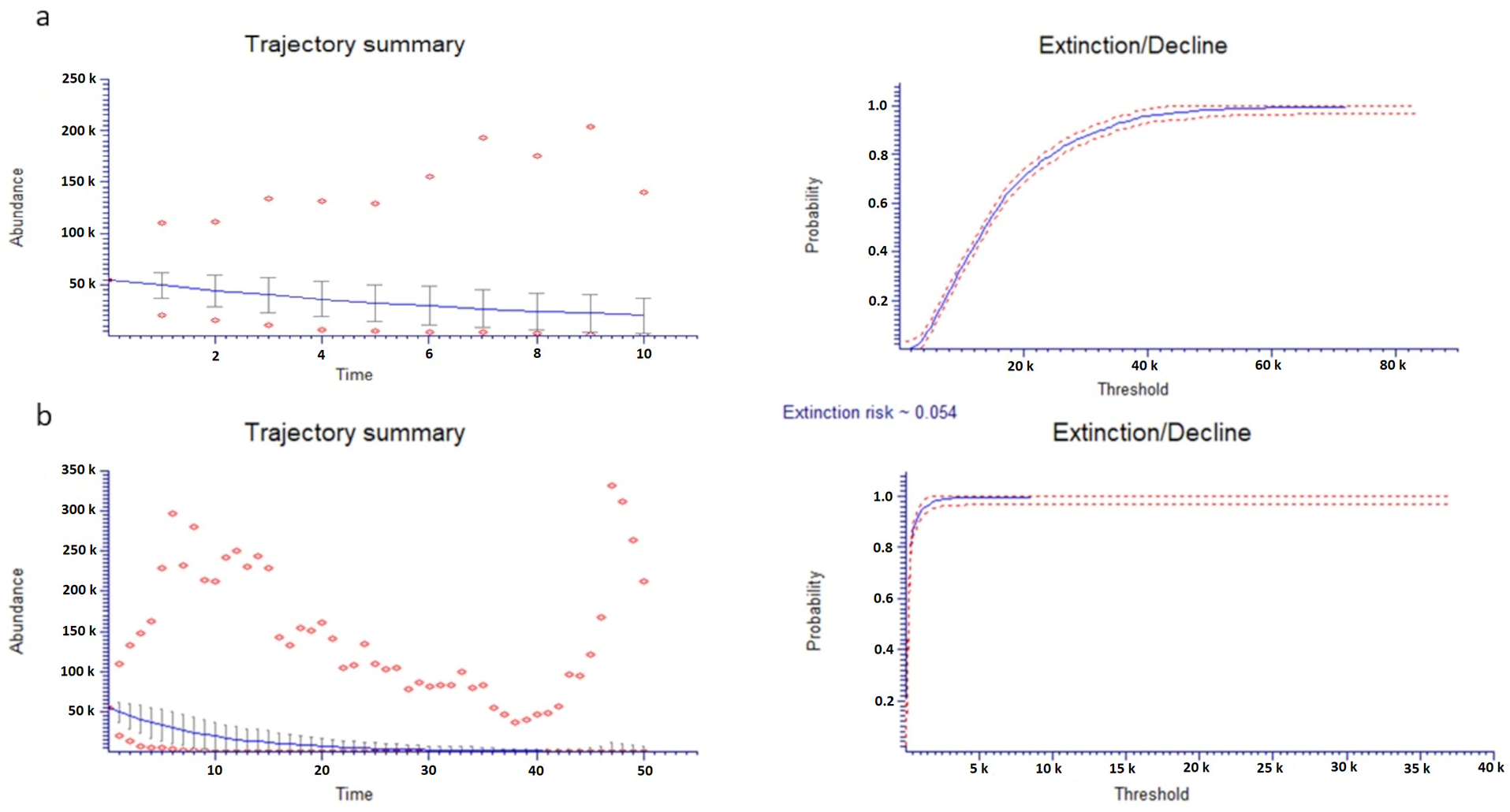
2.5. Population Viability Analysis (PVA)
2.6. Threats
2.7. Conservation Status Assessment
3. Discussion
4. Materials and Methods
4.1. Definitions
4.2. Spatial Distribution
4.3. Population Size
4.4. Reproductive Biology
4.5. Chlorophyll Fluorescence Measurements
4.6. Population Viability Analysis and Conservation Status Assessment
4.7. Threats
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABS/RC | absorbed photon flux per active reaction centre |
| Amax | maximum photosynthetic rate |
| DI0/RC | dissipated energy flux per active reaction centre |
| PItotal | potential for energy conservation from exciton to the reduction in PSI end acceptors |
| VK/VJ | measure of the relative amplitude of K band |
| φP0 | maximum quantum yield of primary PSII photochemistry |
| φR0 | quantum yield for reduction in end electron acceptors at the PSI acceptor side |
| EOO | Extent of Occurrence |
| AOO | Area of Occupancy |
| Local Extent | local extent of occurrence of each subpopulation, i.e., the minimum area polygon or polygons including all the plant individuals/colonies not separated by unsuitable habitat at each location |
| SP | subpopulation |
References
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.-H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent Anthropogenic Plant Extinctions Differ in Biodiversity Hotspots and Coldspots. Curr. Biol. 2019, 29, 2912–2918. [Google Scholar] [CrossRef]
- Pereira, H.M.; Leadley, P.W.; Proença, V.; Alkemade, R.; Scharlemann, J.P.W.; Fernandez-Manjarrés, J.F.; Araújo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for Global Biodiversity in the 21st Century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef] [Green Version]
- Brummitt, N.A.; Bachman, S.P.; Griffiths-Lee, J.; Lutz, M.; Moat, J.F.; Farjon, A.; Donaldson, J.S.; Hilton-Taylor, C.; Meagher, T.R.; Albuquerque, S.; et al. Green Plants in the Red: A Baseline Global Assessment for the IUCN Sampled Red List Index for Plants. PLoS ONE 2015, 10, e0135152. [Google Scholar] [CrossRef] [Green Version]
- Medail, F.; Quezel, P. Hot-Spots Analysis for Conservation of Plant Biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112. [Google Scholar] [CrossRef]
- Vogiatzakis, I.N.; Mannion, A.M.; Sarris, D. Mediterranean island biodiversity and climate change: The last 10,000 years and the future. Biodivers. Conserv. 2016, 25, 2597–2627. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Rosselló, J.A.; Cosín, R.; Bacchetta, G.; Brullo, S.; Mayol, M. Nuclear and chloroplast DNA variation in Cephalaria squamiflora (Dipsacaceae), a disjunct Mediterranean species. Taxon 2009, 58, 1242–1253. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M.; Matthews, T.J.; Borregaard, M.K.; Triantis, K.A. Island biogeography: Taking the long view of nature’s laboratories. Science 2017, 357, eaam8326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliades, N.-G.H.; Andreou, M.; Laguna, E.; Kounnamas, C.; Georghiou, K.; Costantinou, C.; Kouzali, I.; Thanos, C.A.; Kadis, C. Integrated conservation of important plant taxa through the improvement of the original plant micro-reserve (PMR) approach: The intensive PMR monitoring case of Ophrys kotschyi. J. Environ. Manag. 2021, 280, 111731. [Google Scholar] [CrossRef] [PubMed]
- Valli, A.-T.; Kougioumoutzis, K.; Iliadou, E.; Panitsa, M.; Trigas, P. Determinants of alpha and beta vascular plant diversity in Mediterranean island systems: The Ionian islands, Greece. Nord. J. Bot. 2018, 37, e02156. [Google Scholar] [CrossRef] [Green Version]
- Valli, A.-T.; Koumandou, V.L.; Iatrou, G.; Andreou, M.; Papasotiropoulos, V.; Trigas, P. Conservation biology of threatened Mediterranean chasmophytes: The case of Asperula naufraga endemic to Zakynthos island (Ionian islands, Greece). PLoS ONE 2021, 16, e0246706. [Google Scholar] [CrossRef]
- Lyons, K.G.; Schwartz, M.W. Rare species loss alters ecosystem function—Invasion resistance. Ecol. Lett. 2001, 4, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Lyons, K.G.; Brigham, C.A.; Traut, B.; Schwartz, M.W. Rare Species and Ecosystem Functioning. Conserv. Biol. 2005, 19, 1019–1024. [Google Scholar] [CrossRef]
- Ferrer-Sánchez, Y.; Rodríguez-Estrella, R. How rare species conservation management can be strengthened with the use of ecological niche modelling: The case for endangered endemic Gundlach’s Hawk and Cuban Black-Hawk. Glob. Ecol. Conserv. 2016, 5, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Crain, B.J.; Sánchez-Cuervo, A.M.; White, J.W.; Steinberg, S.J. Conservation ecology of rare plants within complex local habitat networks. Oryx 2014, 49, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Lavergne, S.; Thompson, J.D.; Garnier, E.; Debussche, M. The biology and ecology of narrow endemic and widespread plants: A comparative study of trait variation in 20 congeneric pairs. Oikos 2004, 107, 505–518. [Google Scholar] [CrossRef]
- Gulías, J.; Flexas, J.; Mus, M.; Cifre, J.; Lefi, E.; Medrano, H. Relationship between Maximum Leaf Photosynthesis, Nitrogen Content and Specific Leaf Area in Balearic Endemic and Non-endemic Mediterranean Species. Ann. Bot. 2003, 92, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Catoni, R.; Gratani, L. Morphological and physiological adaptive traits of Mediterranean narrow endemic plants: The case of Centaurea gymnocarpa (Capraia Island, Italy). Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 174–183. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Adams, V.M.; Marsh, D.M.; Knox, J.S. Importance of the seed bank for population viability and population monitoring in a threatened wetland herb. Biol. Conserv. 2005, 124, 425–436. [Google Scholar] [CrossRef]
- Lehtila, K.; Syrjanen, K.; Leimu, R.; Garcia, M.B.; Ehrlen, J. Habitat change and demography of Primula veris: Identification of management targets. Conserv. Biol. 2006, 20, 833–843. [Google Scholar] [CrossRef]
- Massey, J.R.; Whitson, P.D. Species Biology, the Key to Plant Preservation. Rhodora 1980, 82, 97–103. [Google Scholar]
- Schemske, D.W.; Husband, B.C.; Ruckelshaus, M.H.; Goodwillie, C.; Parker, I.M.; Bishop, J.G. Evaluating Approaches to the Conservation of Rare and Endangered Plants. Ecology 1994, 75, 584–606. [Google Scholar] [CrossRef] [Green Version]
- Marsh, D.M.; Trenham, P.C. Current Trends in Plant and Animal Population Monitoring. Conserv. Biol. 2008, 22, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Yoccoz, N.G.; Nichols, J.D.; Boulinier, T. Monitoring of biological diversity in space and time. Trends Ecol. Evol. 2001, 16, 446–453. [Google Scholar] [CrossRef]
- Pino, J.; De Roa, E. Population biology of Kosteletzkya pentacarpos (Malvaceae) in the Llobregat delta (Catalonia, NE of Spain). Plant Ecol. 2006, 188, 1–16. [Google Scholar] [CrossRef]
- Davy, A.J.; Jefferies, R.L. Approaches to the Monitoring of Rare Plant Populations. In The Biological Aspects of Rare Plant Conservation; Synge, H., Ed.; John Wiley and Sons: New York, NY, USA, 1981; pp. 219–232. [Google Scholar]
- Pavlik, B.M. Perspectives, Tools, and Institutions for Conserving Rare Plants. Southwest. Nat. 1997, 42, 375–383. [Google Scholar]
- Donovan, T.M.; Welden, C.W. Conservation Biology and Landscape Ecology; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Pavlik, B.M. Demographic Monitoring and the Recovery of Endangered Plant Populations. In Restoration of Endangered Species; Cambridge University Press: Cambridge, UK, 1994; pp. 322–350. [Google Scholar]
- Yates, C.J.; Broadhurst, L.M. Assessing limitations on population growth in two critically endangered Acacia taxa. Biol. Conserv. 2002, 108, 13–26. [Google Scholar] [CrossRef]
- Tan, K.; Zielinski, J. “Micromeria browiczii” (Labiatae), an unusual new species from Zakinthos (Ionian Islands, Greece). Pol. Bot. J. 2001, 46, 31–34. [Google Scholar]
- Šerá, B.; Feráková, V.; Kocianová, E.; Vágenknecht, V.; Majzlan, O.; Dúbravcová, Z. Peucedanum arenarium subsp. arenarium—A critically endangered species of the Slovak flora (morphometry, distribution, biology). Biologia 2005, 60, 17–23. [Google Scholar]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria; Version 14; IUCN: Gland, Switzerland, 2019; Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 1 September 2020).
- Tienes, M.; Skogen, K.; Vitt, P.; Havens, K. Optimal Monitoring of Rare Plant Populations: Report for the USDA Forest Service; Chicago Botanic Garden: Glencoe, IL, USA, 2010. [Google Scholar]
- Heywood, V.H.; Iriondo, J.M. Plant conservation: Old problems, new perspectives. Biol. Conserv. 2003, 113, 321–335. [Google Scholar] [CrossRef]
- Kier, G.; Kreft, H.; Lee, T.M.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [Green Version]
- Weigelt, P.; Jetz, W.; Kreft, H. Bioclimatic and physical characterization of the world’s islands. Proc. Natl. Acad. Sci. USA 2013, 110, 15307–15312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, R.W.; Forseth, I.N.; Casper, B.B. Using rainout shelters to evaluate climate change effects on the demography of Cryptantha flava. J. Ecol. 2008, 96, 514–522. [Google Scholar] [CrossRef]
- Dahlgren, D.K.; Guttery, M.R.; Messmer, T.A.; Caudill, D.; Elmore, R.D.; Chi, R.; Koons, D.N. Evaluating vital rate contributions to greater sage-grouse population dynamics to inform conservation. Ecosphere 2016, 7, e01249. [Google Scholar] [CrossRef] [Green Version]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Collet, C.; Le Moguedec, G. Individual seedling mortality as a function of size, growth and competition in naturally regenerated beech seedlings. Forestry 2007, 80, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.; Hoff, W.D. Water Transport in Brick, Stone and Concrete; Taylor & Francis: Abingdon, UK, 2002. [Google Scholar]
- Moore, L.M.; Lauenroth, W.K. Differential effects of temperature and precipitation on early- vs. late-flowering species. Ecosphere 2017, 8, e01819. [Google Scholar] [CrossRef] [Green Version]
- Gordon, C.; Llorens, L.; Nielsen, T.; Tietema, A.; Beier, C.; Bruna, P.; Emmett, B.; Estiarte, M.; Gorissen, A. Nonintrusive Field Experiments Show Different Plant Responses to Warming and Drought Among Sites, Seasons, and Species in a North–South European Gradient. Ecosystems 2004, 7, 598–612. [Google Scholar] [CrossRef]
- Prieto, P.; Peñuelas, J.; Ogaya, R.; Estiarte, M. Precipitation-dependent Flowering of Globularia alypum and Erica multiflora in Mediterranean Shrubland Under Experimental Drought and Warming, and its Inter-annual Variability. Ann. Bot. 2008, 102, 275–285. [Google Scholar] [CrossRef] [PubMed]
- De Jong, T.J.; Roo, L.G.-D.; Klinkhamer, P.G.L. Is the threshold size for flowering in Cynoglossum officinale fixed or dependent on environment? New Phytol. 1998, 138, 489–496. [Google Scholar] [CrossRef]
- BrysRichard, R.; Shefferson, R.P.; Jacquemyn, H. Impact of herbivory on flowering behaviour and life history trade-offs in a polycarpic herb: A 10-year experiment. Oecologia 2010, 166, 293–303. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Gouvra, E.; Grammatikopoulos, G. Diurnal and seasonal trends of water relations in five co-occurring chasmophytic species. Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 237–248. [Google Scholar] [CrossRef]
- Chondrogiannis, C.; Grammatikopoulos, G. Photosynthesis in developing leaf of juveniles and adults of three Mediterranean species with different growth forms. Photosynth. Res. 2016, 130, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.H. Physiological Aspects of Ex Situ Seed Conservation. In Conservation Biology; Kapoor-Vijay, P., White, J., Eds.; The Commonwealth Secretariat: London, UK, 1992; pp. 171–177. [Google Scholar]
- Maunder, M.; Guerrant, E.O.; Havens, K.; Dixon, K.W. Realizing the Full Potential of Ex Situ Contributions to Global Plant Conservation. In Ex Situ Plant Conservation: Supporting Species Survival in the Wild; Guerrant, E.O., Havens-Young, K., Maunder, M., Eds.; Society for Ecological Restoration International: Washington, DC, USA; Center for Plant Conservation: Escondido, CA, USA; Island Press: Washington, DC, USA, 2004; pp. 389–418. [Google Scholar]
- IUCN. Threats Classification Scheme; Version 3.2; IUCN: Gland, Switzerland, 2012; Available online: https://www.iucnredlist.org/technicaldocuments/classification-schemes/threatsclassification-scheme (accessed on 20 May 2020).
- Andreou, M.; Delipetrou, P.; Kadis, C.; Tsiamis, G.; Bourtzis, K.; Georghiou, K. An integrated approach for the conservation of threatened plants: The case of Arabis kennedyae (Brassicaceae). Acta Oecologica 2011, 37, 239–248. [Google Scholar] [CrossRef]
- Andreou, M.; Kadis, C.; Delipetrou, P.; Georghiou, K. Conservation biology of Chionodoxa lochiae and Scilla morrisii (Asparagaceae): Two priority bulbous plant species of the European Union in Cyprus. Glob. Ecol. Conserv. 2015, 3, 511–525. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.H.; Pardini, E.A.; Schutzenhofer, M.R.; Chung, Y.A.; Seidler, K.J.; Knight, T.M. Greater sexual reproduction contributes to differences in demography of invasive plants and their noninvasive relatives. Ecology 2013, 94, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- Stirbet, A. Govindjee on the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Rexstad, E.; Akçakaya, H.R.; Burgman, M.A.; Ginzburg, L.R. Applied Population Ecology: Principles and Computer Exercises Using RAMAS EcoLab. J. Wildl. Manag. 2000, 64, 1093. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | IUCN SP | Colony | Altitude (m) | Longitude | Latitude | Aspect (°) | Slope (°) | Substrate | Ao (cm) | Local Extent (km2) | AOO (km2) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Marathias | Mb1 | M1 | 104–112 | 20.851298° | 37.667198° | 22.5–67.5 (NE) | 45–90 | Rock | 1.5 | 0.0001257 | 8 |
| M2 | 21–316 | 67.5–112.5 (E) | |||||||||
| Faros Keriou | Mb2 | F1 | 141–188 | 20.807577° | 37.656210° | 247–337.5 (W. NW) | 45–90 | Rock | 1.5 | 0.008016 | 4 |
| F2 | |||||||||||
| F3 | |||||||||||
| Ethniki | Mb3 | 154–159 | 20.812783° | 37.669883° | 202.5–247.5 (SW) | 80 | Rock | 1.5 | 0.000027 | 4 | |
| Plakaki (l.c.) | Mb4 | Pl1 | 81–220 | 20.777832° | 37.689664° | 247.5–292.5 (W. S) | 45–110 | Rock | 1.5 | 0.102 | 4 |
| Pl2 | 202.5–247.5 (SW) | ||||||||||
| Psaris | Mb5 | 183–227 | 20.769923° | 37.698439° | 247.5–337.5 (NW. W) | 20–50 | Rock | 1.5 | 0.01119 | 4 | |
| Gravel | 7.9 | ||||||||||
| Agalas | Mb6 | 272–287 | 20.774257° | 37.707252° | 202.5–247.5 (SW) | 45–90 | Wall | 0.7 | 0.00218 | 4 | |
| Rock | 1.5 | ||||||||||
| Korakonisi | Mb7 | 32–205 | 20.739322° | 37.722931° | 22.5–67.5 (NE) | 20–45 | Rock | 1.5 | 0.01295 | 4 | |
| Limnionas | Mb8 | 21–178 | 20.703129° | 37.746664° | 157.5–247.5 (SW. S) | 30–90 | Rock | 1.5 | 0.02941 | 12 | |
| Kampi | Mb9 | K1 | 166–182 | 20.681802° | 37.780336° | 337.5–360 (N) | 10–50 | Rock | 1.5 | 0.002303 | 4 |
| K2 | 158–167 | 292.5–337.5 (NW) | |||||||||
| K3 | 163 | 292.5–337.5 (NW) | |||||||||
| K4 | 164 | 247.5–292.5 (W) | |||||||||
| Maries | Mb10 | 343–416 | 20.678940° | 37.819131° | 202.5–292.5 (SW. W) | 45–90 | Rock | 1.5 | 0.000653 | 4 | |
| Porto Vromi | Mb11 | 13–228 | 20.631936° | 37.829057° | 112.5–292.5 (SE. S. SW. W) | 45–90 | Rock | 1.5 | 0.01549 | 4 | |
| Anafonitria | Mb12 | 310–317 | 20.644937° | 37.844176° | 202.5–247.5 (SW) | 90 | Wall | 0.7 | 0.0000255 | 4 | |
| Aghios Georgios | Mb13 | 309–325 | 20.635926° | 37.859460° | 205.5–247.5 (SW) | 10–90 | Gravel | 7.9 | 0.000402 | 4 | |
| Navagio | Mb14 | 200–229 | 20.625690° | 37.862401° | 247.5–292.5 (W) | 45 | Rock | 1.5 | 0.010378 | 4 | |
| Gravel | 7.9 | ||||||||||
| Xigia | Mb15 | 10–38 | 20.731328° | 37.879880° | 67.5–112.5 (E) | 30–90 | Rock | 1.5 | 0.006244 | 8 |
| SP | Colony | 2014 | 2015 | 2016 | 2017 | 2018 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SP Size | L. extent (m2) | Plants/m2 | SP size | L. extent (m2) | Plants/m2 | SP Size | L. extent (m2) | Plants/m2 | SP Size | L. extent (m2) | Plants/m2 | SP Size | L. extent (m2) | Plants/m2 | ||
| Mb1 | M1 | 4 | 28.1 | 0.14 | 2 | 28.1 | 0.07 | 0 | 28.1 | 0 | 2 | 28.1 | 0.07 | 12 | 28.1 | 0.43 |
| M2 | 1 | 97.5 | 0.01 | 11 | 97.5 | 0.113 | 8 | 97.5 | 0.08 | 4 | 97.5 | 0.04 | 81 | 97.5 | 0.83 | |
| Mb2 | F1 | 19 | 3790 | 0.003 | 33 | 3790 | 0.009 | 27 | 3790 | 0.007 | 52 | 3790 | 0.014 | 20 | 250 | 0.08 |
| F2 | 2 | 727 | 0.003 | 3 | 727 | 0.004 | 14 | 727 | 0.02 | 7 | 727 | 0.01 | 4 | 727 | 0.006 | |
| F3 | 4 | 9192 | 0.0004 | 6 | 9192 | 0.0007 | 35 | 9192 | 0.004 | 11 | 9192 | 0.0012 | 37 | 9192 | 0.004 | |
| Mb3 | 13 | 27 | 0.48 | 1 | 27 | 0.037 | 11 | 27 | 0.4 | 11 | 27 | 0.4 | 27 | 27 | 1 | |
| Mb4 | Pl1 | 124 | 92,653 | 0.0013 | 32 | 92,652 | 0.0003 | 81 | 92,652 | 0.0009 | 41 | 92,652 | 0.0004 | 99 | 926,52 | 0.0001 |
| Pl2 | 43 | 9411 | 0.005 | 17 | 9411 | 0.002 | 92 | 9411 | 0.01 | 152 | 9411 | 0.02 | 43 | 120 | 0.36 | |
| Mb5 | 3245 | 11,192 | 0.3 | 1354 | 11,192 | 0.12 | 3562 | 11,192 | 0.32 | 1260 | 11,192 | 0.112 | 1632 | 11,192 | 0.146 | |
| Mb6 | 61 | 2180 | 0.028 | 63 | 2180 | 0.028 | 86 | 2180 | 0.039 | 87 | 2180 | 0.04 | 276 | 2180 | 0.13 | |
| Mb7 | 1425 | 12,950 | 0.11 | 1166 | 12,950 | 0.09 | 1036 | 12,950 | 0.08 | 259 | 12,950 | 0.019 | 907 | 12,950 | 0.07 | |
| Mb8 | 782 | 29,412 | 0.026 | 382 | 29,412 | 0.012 | 2147 | 29,412 | 0.07 | 441 | 29,412 | 0.015 | 3824 | 29,412 | 0.13 | |
| Mb9 | K1 | 59 | 1546 | 0.04 | 26 | 1546 | 0.017 | 61 | 1546 | 0.04 | 25 | 1546 | 0.016 | 48 | 1546 | 0.03 |
| K2 | 15 | 17.5 | 0.86 | 10 | 17.5 | 0.57 | 14 | 17.5 | 0.8 | 19 | 17.5 | 1.09 | 14 | 17.5 | 0.8 | |
| K3 | 14 | 475.7 | 0.03 | 18 | 475.7 | 0.038 | 25 | 475.7 | 0.05 | 20 | 475.7 | 0.04 | 34 | 475.7 | 0.07 | |
| K4 | 6 | 90.8 | 0.07 | 0 | 90.8 | 0 | 1 | 90.8 | 0.01 | 0 | 90.8 | 0 | 5 | 90.8 | 0.06 | |
| Mb10 | 164 | 653 | 0.3 | 336 | 653 | 0.5 | 372 | 653 | 0.56 | 568 | 653 | 0.87 | 347 | 653 | 0.53 | |
| Mb11 | 926 | 15,490 | 0.06 | 1431 | 15,490 | 0.09 | 1345 | 15,490 | 0.086 | 168 | 15,490 | 0.01 | 168 | 15,490 | 0.01 | |
| Mb12 | 86 | 25.5 | 3.4 | 185 | 25.5 | 7.25 | 184 | 25.5 | 7.2 | 239 | 25.5 | 9.37 | 274 | 25.5 | 10.6 | |
| Mb13 | 23 | 402 | 0.06 | 12 | 402 | 0.029 | 29 | 402 | 0.07 | 5 | 402 | 0.01 | 41 | 402 | 0.1 | |
| Mb14 | 115 | 10,378 | 0.01 | 65 | 10,378 | 0.006 | 88 | 10,378 | 0.008 | 58 | 10,378 | 0.0055 | 54 | 10,378 | 0.0052 | |
| Mb15 | 1374 | 6244 | 0.22 | 499 | 6244 | 0.08 | 312 | 6244 | 0.05 | 300 | 6244 | 0.048 | 687 | 6244 | 0.11 | |
| TOTAL | 8547 | 22,2831 | 5641 | 222,831 | 9523 | 22,2831 | 3725 | 222,831 | 8579 | 210,000 | ||||||
| Year | Seedlings (%) | Non-Reproductive Individuals (%) | Mature Individuals (%) |
|---|---|---|---|
| 2014 | 12.01 | 72.29 | 15.69 |
| 2015 | 6.46 | 78.69 | 14.86 |
| 2016 | 12.35 | 61.06 | 26.88 |
| 2017 | 12.18 | 79.18 | 8.64 |
| 2018 | 6.43 | 67.61 | 25.94 |
| 2014 | n | 2015 | n | 2016 | n | 2017 | n | 2018 | n | |
|---|---|---|---|---|---|---|---|---|---|---|
| Stems/individuals ± SE | 28.1 ± 5 a,* | 60 | 21.24 ± 5.5 a | 60 | 12.2 ± 1.8 b | 115 | 12.38 ± 1.4 a | 60 | 14.7 ± 1.9 a | 60 |
| Flowering or fruiting stems per individual (F) ± SE | 5.43 ± 0.7 a | 60 | 7.89 ± 1.49 a | 60 | 6.38 ± 1.58 a | 115 | 5.03 ± 0.57 a | 60 | 6.58 ± 0.97 a | 60 |
| Mean number of flowers per stem ± SE | 7.18 ± 0.99 a | 60 | 3.48 ± 0.36 b | 60 | 5.38 ± 0.55 a | 115 | 7.75 ± 0.69 c | 60 | 4.88 ± 0.55 a | 60 |
| Mean number of flowers per individual ± SE | 29.25 ± 6.3 a | 60 | 22.9 ± 3.7 a | 60 | 36.4 ± 10.09 a | 115 | 43.5 ± 8.08 a | 60 | 42.55 ± 9.6 a | 60 |
| Mean number of fruits (nutlets) per flower | 1.78 ± 0.17 a | 120 | 1.93 ± 0.08 a | 120 | 2.22 ± 0.14 a | 230 | 2.095 ± 0.13 a | 120 | 2.13 ± 0.19 a | 120 |
| Mean number of fruits (nutlets) per stem (S) ± SE | 50.5 ± 6.8 a | 120 | 38.09 ± 5 a | 120 | 48.03 ± 6.5 a | 230 | 48.95 ± 9.2 a | 120 | 40.89 ± 4.06 a | 120 |
| Fecundity (mean number of fruits per individual) (S × F) | 274.2 | 300.5 | 306.4 | 246.2 | 269.06 | |||||
| Seed rain (seeds/m2) | 0.065 | 0.028 | 0.086 | 0.04 | 0,047 | |||||
| Survival of juveniles (%) (Sj) | 64.5 | 45 | 35 | 70 | ||||||
| Relative Reproductive Success (%) | 44.5 ± 4.35 a | 48.26 ± 2.1 a | 55.34 ± 3.5 b | 52.4 ± 3.2 a | 53.6 ± 3.88 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valli, A.-T.; Chondrogiannis, C.; Grammatikopoulos, G.; Iatrou, G.; Trigas, P. Conservation of Micromeria browiczii (Lamiaceae), Endemic to Zakynthos Island (Ionian Islands, Greece). Plants 2021, 10, 778. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040778
Valli A-T, Chondrogiannis C, Grammatikopoulos G, Iatrou G, Trigas P. Conservation of Micromeria browiczii (Lamiaceae), Endemic to Zakynthos Island (Ionian Islands, Greece). Plants. 2021; 10(4):778. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040778
Chicago/Turabian StyleValli, Anna-Thalassini, Christos Chondrogiannis, George Grammatikopoulos, Gregoris Iatrou, and Panayiotis Trigas. 2021. "Conservation of Micromeria browiczii (Lamiaceae), Endemic to Zakynthos Island (Ionian Islands, Greece)" Plants 10, no. 4: 778. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040778