
Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropods in a Tomato Field Margin

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Sown Plants Establishment and Flowering
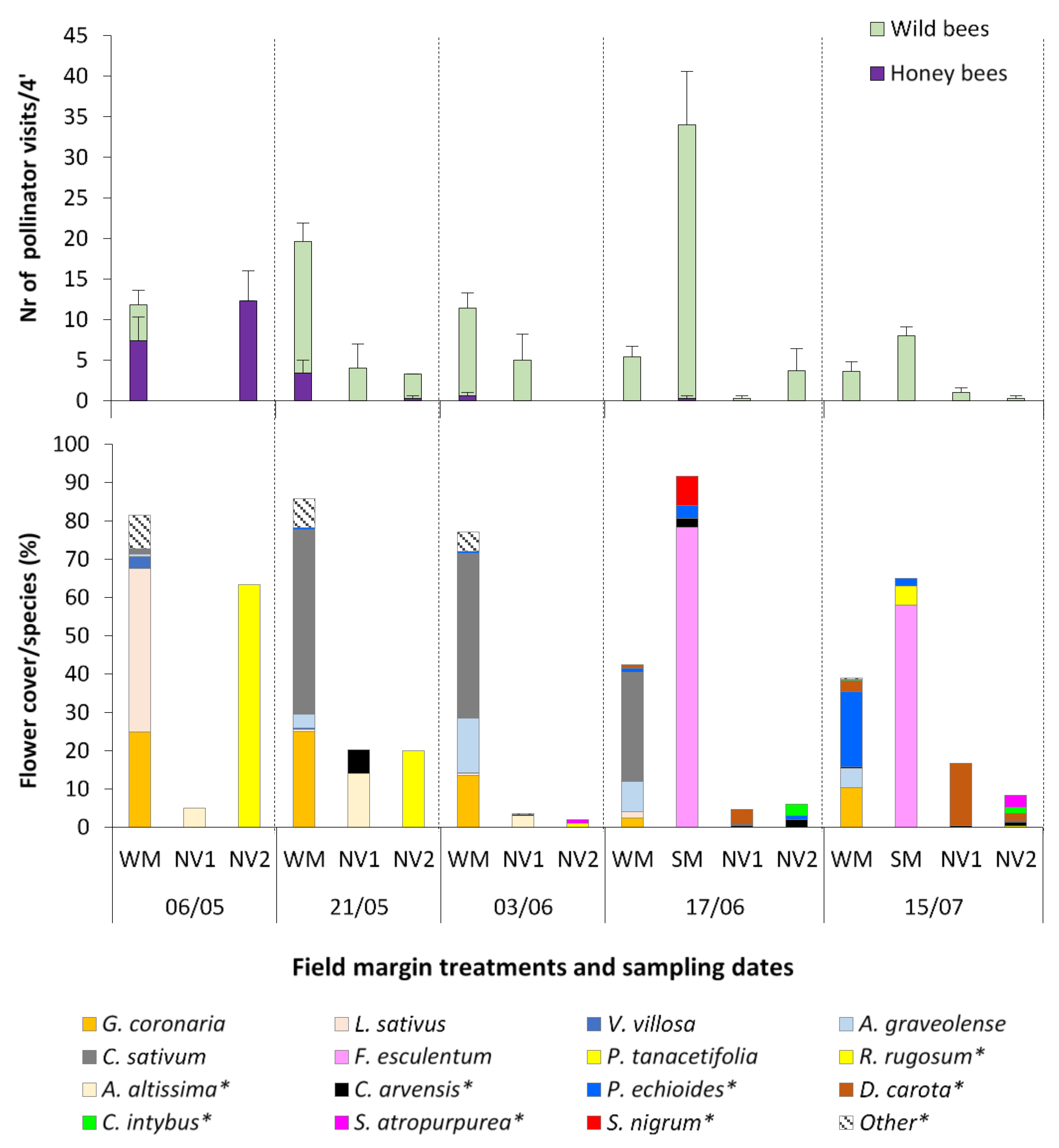
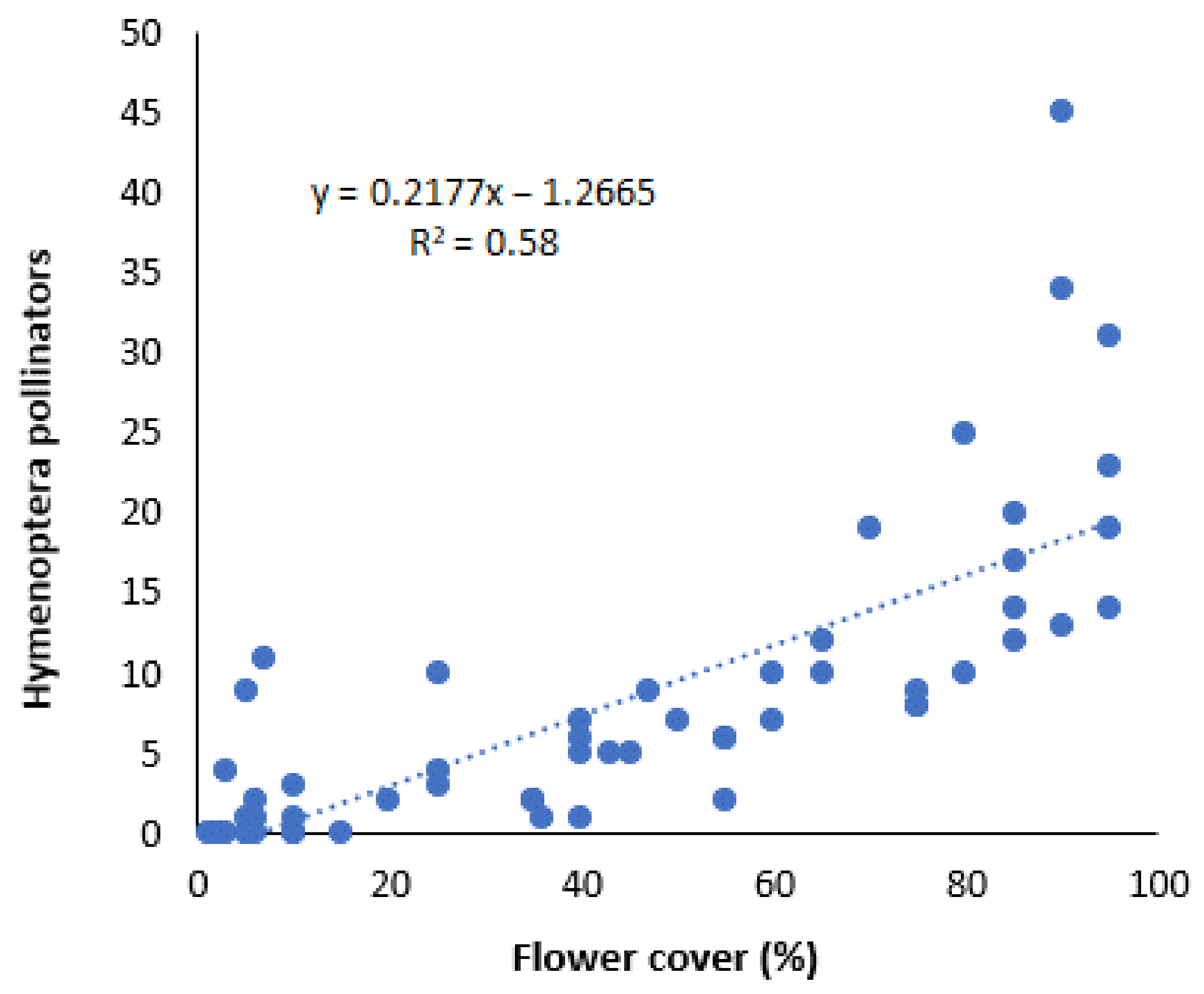
2.2. Effect of Wildflower Margins on Pollinator Abundance and Diversity
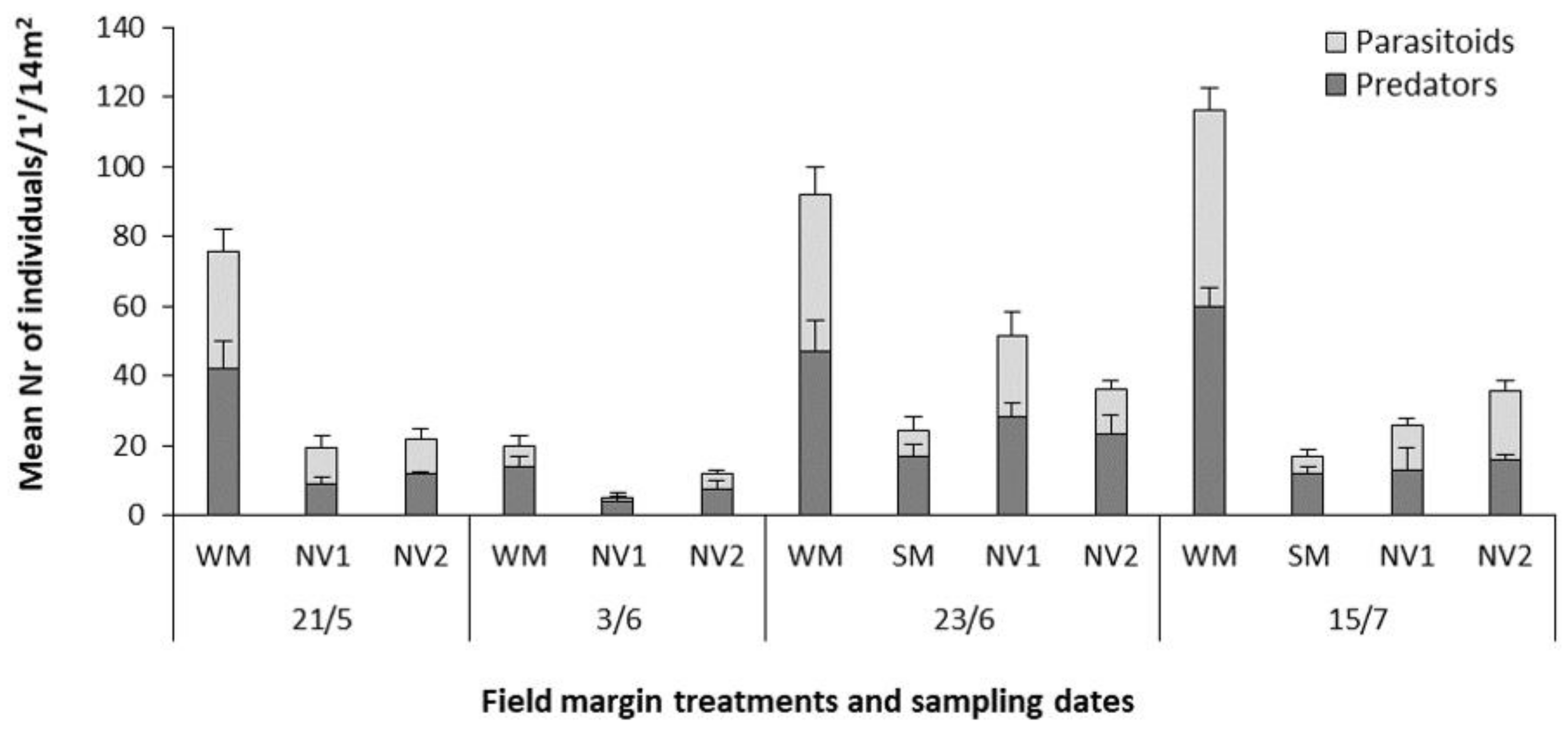
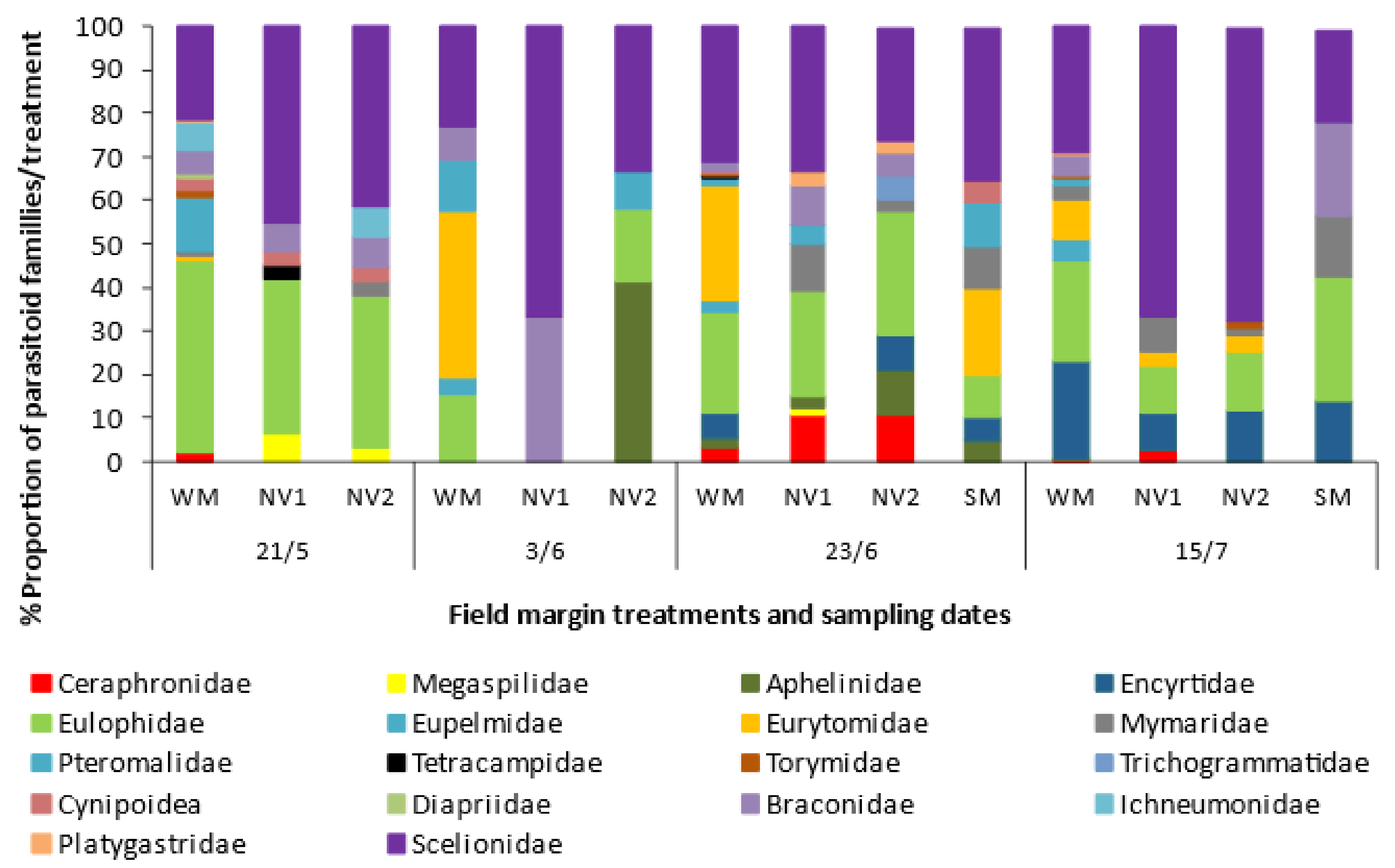
2.3. Effect of Wildflower Margins on Arthropod Abundance and Diversity
3. Discussion
4. Materials and Methods
4.1. Selection of Plant Species
4.2. Seed-Rate Calculation
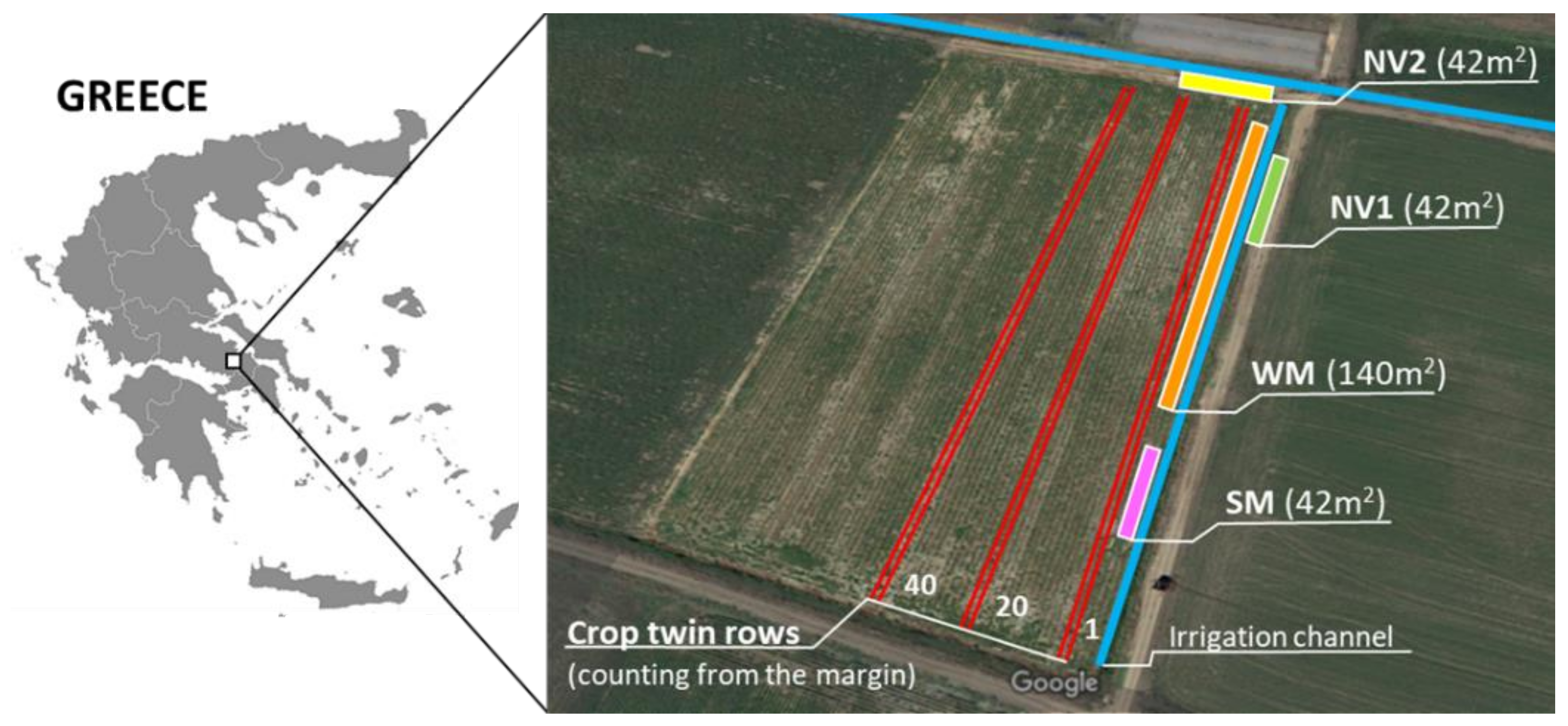
4.3. Experimentation Site
4.4. Measurements
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Naylor, R.; Ehrlich, P.R. Natural Pest Control Services and Agriculture. In Nature’s Services: Societal Dependence on Natural Ecosystems; Daily, G.C., Ed.; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Dale, V.H.; Polasky, S. Measures of the effects of agricultural practices on ecosystem services. Ecol. Econ. 2007, 64, 286–296. [Google Scholar] [CrossRef]
- German, R.N.; Thompson, C.E.; Benton, T.G. Relationships among multiple aspects of agriculture’s environmental impact and productivity: A meta-analysis to guide sustainable agriculture. Biol. Rev. Camb. Philos. Soc. 2017, 92, 716–738. [Google Scholar] [CrossRef] [PubMed]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the interactions between resource availability and the utilisation of semi-natural habitats by insect pollinators in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Brandt, K.; Glemnitz, M.; Schröder, B. The impact of crop parameters and surrounding habitats on different pollinator group abundance on agricultural fields. Agric. Ecosyst. Environ. 2017, 243, 55–66. [Google Scholar] [CrossRef]
- Perovic, D.J.; Gamez-Virues, S.; Landis, D.A.; Wackers, F.; Gurr, G.M.; Wratten, S.D.; You, M.; Desneux, N. Managing biological control services through multi-trophic trait interactions: Review and guidelines for implementation at local and landscape scales. Biol. Rev. Camb. Philos. Soc. 2017, 93, 306–321. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, G.J.K.; Holland, J.M.; Bailey, A.; Thomas, M.B. Efficacy and economics of shelter habitats for conservation biological control. Biol. Control 2008, 45, 200–209. [Google Scholar] [CrossRef]
- Rollin, O.; Pérez-Méndez, N.; Bretagnolle, V.; Henry, M. Preserving habitat quality at local and landscape scales increases wild bee diversity in intensive farming systems. Agric. Ecosyst. Environ. 2019, 275, 73–80. [Google Scholar] [CrossRef]
- Rollin, O.; Benelli, G.; Benvenuti, S.; Decourtye, A.; Wratten, S.D.; Canale, A.; Desneux, N. Weed-insect pollinator networks as bio-indicators of ecological sustainability in agriculture. A review. Agron. Sustain. Dev. 2016, 36. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; West, T.M.; Kleijn, D. Impacts of an agri-environment field margin prescription on the flora and fauna of arable farmland in different landscapes. Agric. Ecosyst. Environ. 2006, 113, 36–44. [Google Scholar] [CrossRef]
- Page, G.; Bellotti, B. Farmers value on-farm ecosystem services as important, but what are the impediments to participation in PES schemes? Sci. Total Environ. 2015, 515–516, 12–19. [Google Scholar] [CrossRef]
- Balzan, M.V.; Bocci, G.; Moonen, A.-C. Landscape complexity and field margin vegetation diversity enhance natural enemies and reduce herbivory by Lepidoptera pests on tomato crop. BioControl 2016, 61, 141–154. [Google Scholar] [CrossRef]
- Fiedler, A.K.; Landis, D.A.; Wratten, S.D. Maximizing ecosystem services from conservation biological control: The role of habitat management. Biol. Control 2008, 45, 254–271. [Google Scholar] [CrossRef]
- Storkey, J.; Döring, T.; Baddeley, J.; Collins, R.; Roderick, S.; Jones, H.; Watson, C. Engineering a plant community to deliver multiple ecosystem services. Ecol. Appl. 2015, 25, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- European Commission, Eurostat, and Directorate General for Economic and Financial Affairs. In Statistical Factsheet, Greece; DG Agriculture and Rural Development, Farm Economics Unit: Brussels, Belgium, 2020.
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground cover management with mixtures of flowering plants to enhance insect pollinators and natural enemies of pests in olive groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Potts, S.G.; Petanidou, T.; Roberts, S.; O’Toole, C.; Hulbert, A.; Willmer, P. Plant-pollinator biodiversity and pollination services in a complex Mediterranean landscape. Biol. Conserv. 2006, 129, 519–529. [Google Scholar] [CrossRef]
- Solomou, A.; Sfougaris, A. Comparing conventional and organic olive groves in central Greece: Plant and bird diversity and abundance. Renew. Agric. Food Syst. 2011, 26, 297–316. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Kremen, C. Wild bee species increase tomato production and respond differently to surrounding land use in Northern California. Biol. Conserv. 2006, 133, 81–87. [Google Scholar] [CrossRef]
- Teppner, H. Pollinators of tomato, Solanum lycopersicum (Solanaceae), in Central Europe. Phyton Ann. Rei Bot. 2005, 45, 217–235. [Google Scholar]
- Balzan, M.V.; Bocci, G.; Moonen, A.-C. Augmenting flower trait diversity in wildflower strips to optimise the conservation of arthropod functional groups for multiple agroecosystem services. J. Insect Conserv. 2014, 18, 713–728. [Google Scholar] [CrossRef]
- Balzan, M.V.; Moonen, A.-C. Field margin vegetation enhances biological control and crop damage suppression from multiple pests in organic tomato fields. Entomol. Exp. Appl. 2014, 150, 45–65. [Google Scholar] [CrossRef]
- Balzan, M.V. Flowering banker plants for the delivery of multiple agroecosystem services. Arthropod Plant Interact. 2017, 11, 743–754. [Google Scholar] [CrossRef]
- Kishinevsky, M.; Keasar, T.; Harari, A.R.; Chiel, E. A comparison of naturally growing vegetation vs. border-planted companion plants for sustaining parasitoids in pomegranate orchards. Agric. Ecosyst. Environ. 2017, 246, 117–123. [Google Scholar] [CrossRef]
- Lavandero, B.; Wratten, S.D.; Didham, R.K.; Gurr, G. Increasing floral diversity for selective enhancement of biological control agents: A double-edged sward? Basic Appl. Ecol. 2006, 7, 236–243. [Google Scholar] [CrossRef]
- Parker, J.E.; Snyder, W.E.; Hamilton, G.C.; Rodriguez-Saona, C. Companion Planting and Insect Pest Control. In Weed and Pest Control—Conventional and New Challenges; Soloneski, S., Larramendy, M.L., Eds.; IntechOpen Limited: London, UK, 2013. [Google Scholar]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Brigham, C.A.; Hoeksema, J.D.; Lyons, K.G.; Mills, M.H.; van Mantgem, P.J. Linking biodiversity to ecosystem function: Implications for conservation ecology. Oecologia 2000, 122, 297–305. [Google Scholar] [CrossRef]
- Moonen, A.-C.; Bàrberi, P. Functional biodiversity: An agroecosystem approach. Agric. Ecosyst. Environ. 2008, 127, 7–21. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Escobar, S.; Galindo, V.; Gutiérrez, C.; López, S.D.; et al. Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Jedlicka, J.A.; Bothwell, S.G.; Moreno, C.R. Effects of natural enemy biodiversity on the suppression of arthropod herbivores in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 573–592. [Google Scholar] [CrossRef]
- Wäckers, F.L. Suitability of (extra-) floral nectar, pollen and honeydew as insect food sources. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Wäckers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 17–74. [Google Scholar]
- Wäckers, F.L.; van Rijn, P.C.J. Pick and mix: Selecting flowering plants to meet the requirements of target biological control insects. In Biodiversity and Insect Pests; Gurr, G.M., Wratten, S.D., Snyder, W.E., Read, D.M.Y., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 139–165. [Google Scholar]
- Campbell, J.W.; Irvin, A.; Irvin, H.; Stanley-Stahr, C.; Ellis, J.D. Insect visitors to flowering buckwheat, Fagopyrum esculentum (Polygonales: Polygonaceae), in North-Central Florida. Fla. Entomol. 2016, 99, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Haaland, C.; Naisbit, R.E.; Bersier, L.-F. Sown wildflower strips for insect conservation: A review. Insect Conserv. Divers. 2011, 4, 60–80. [Google Scholar] [CrossRef]
- Asmussen, C.B. Pollination biology of the Sea Pea, Lathyrus japonicus: Floral characters and activity and flight patterns of bumblebees. Flora 1993, 188, 227–237. [Google Scholar] [CrossRef]
- Sasaki, H.; Wagatsuma, T. Bumblebees (Apidae: Hymenoptera) are the main pollinators of common buckwheat, Fagopyrum esculentum, in Hokkaido, Japan. Appl. Entomol. Zool. 2007, 42, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Usman, M. Incidence of different insect visitors and their relative abundance associated with coriander (Coriandrum sativum) in district Charsadda. Pure Appl. Biol. 2018, 7, 539–546. [Google Scholar] [CrossRef]
- Montalva, J.; Dudley, L.; Arroyo, M.K.; Retamales, H.; Abrahamovich, A.H. Geographic distribution and associated flora of native and introduced bumble bees (Bombus spp.) in Chile. J. Apic. Res. 2015, 50, 11–21. [Google Scholar] [CrossRef]
- Bareke, T.; Addi, A. Effect of honeybee pollination on seed and fruit yield of agricultural crops in Ethiopia. Moj Ecol. Environ. Sci. 2019, 4, 205–209. [Google Scholar] [CrossRef]
- Bendifallah, L.; Louadi, K.; Doumandji, S. Bee fauna potential visitors of coriander flowers Coriandrum sativum L. (Apiaceae) in the Mitidja area (Algeria). J. Apic. Sci. 2013, 57, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Thapa, R. Honeybees and other insect pollinators of cultivated plants: A review. J. Inst. Agric. Anim. Sci. 2006, 27, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Denys, C.; Tscharntke, T. Plant-insect communities and predator-prey ratios in field margin strips, adjacent crop fields, and fallows. Oecologia 2002, 130, 315–324. [Google Scholar] [CrossRef]
- Carreck, N.L.; Williams, I.H.; Oakley, J.N. Enhancing farmland for insect pollinators using flower mixtures. Asp. Appl. Biol. 1999, 54, 101–108. [Google Scholar]
- Petanidou, T. Introducing plants for bee-keeping at any cost?—Assessment of Phacelia tanacetifolia as nectar source plant under xeric Mediterranean conditions. Plant Syst. Evol. 2016, 238, 155–168. [Google Scholar] [CrossRef]
- Skinner, K.; Smith, L.; Rice, P. Using noxious weed lists to prioritize targets for developing weed management strategies. Weed Sci. 2000, 48, 640–644. [Google Scholar] [CrossRef]
- Simmons, M.T. Bullying the bullies: The selective control of an exotic, invasive annual (Rapistrum rugosum) by oversowing with a competitive native species (Gaillardia pulchella). Restor. Ecol. 2005, 13, 609–615. [Google Scholar] [CrossRef]
- Connop, S.; Hill, T.; Steer, J.; Shaw, P. The role of dietary breadth in national bumblebee (Bombus) declines: Simple correlation? Biol. Conserv. 2010, 143, 2739–2746. [Google Scholar] [CrossRef]
- Bennett, A.; Gratton, C. Floral diversity increases beneficial arthropod richness and decreases variability in arthropod community composition. Ecol. Appl. 2013, 23, 86–95. [Google Scholar] [CrossRef]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Wäckers, F.L. Effects of flower attractiveness and nectar availability in field margins on biological control by parasitoids. Biol. Control 2008, 46, 400–408. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Jervis, M.A. Does floral nectar improve biological control by parasitoids. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Wäckers, F.L., Bruin, J., van Rijn, P.C.J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 267–304. [Google Scholar]
- Lu, Z.X.; Zhu, P.Y.; Gurr, G.M.; Zheng, X.S.; Read, D.M.; Heong, K.L.; Yang, Y.J.; Xu, H.X. Mechanisms for flowering plants to benefit arthropod natural enemies of insect pests: Prospects for enhanced use in agriculture. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Samin, N.; Asgari, S. A study on the fauna of Scelionid wasps (Hymenoptera: Platygastroidea: Scelionidae) in the Isfahan province, Iran. Arch. Biol. Sci. 2012, 64, 1073–1077. [Google Scholar] [CrossRef]
- Jervis, M.A.; Kidd, N.A.C.; Fitton, M.G.; Huddleston, T.; Dawah, H.A. Flower-visiting by hymenopteran parasitoids. J. Nat. Hist. 1993, 27, 67–105. [Google Scholar] [CrossRef]
- Yefremova, Z. Order Hymenoptera, family Eulophidae. Arthropod Fauna Uae 2008, 1, 345–360. [Google Scholar]
- Schäfer, L.; Herz, A. Suitability of European Trichogramma species as biocontrol agents against the tomato leaf miner Tuta absoluta. Insects 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Babendreier, D.; Wäckers, F.L.; Shanower, T.G. Habitat and plant specificity of Trichogramma egg parasitoids—Underlying mechanisms and implications. Basic Appl. Ecol. 2005, 6, 215–236. [Google Scholar] [CrossRef]
- Bissinger, B.; Orr, D.B.; Linker, H.M. Effects of floral resources on fitness of the parasitoids Trichogramma exiguum (Hymenoptera: Trichogrammatidae) and Cotesia congregata (Hymenoptera: Braconidae). Biol. Control 2008, 47, 180–186. [Google Scholar] [CrossRef]
- Corbett, A.; Plant, R.E. Role of movement in the response of natural enemies to agroecosystem diversification: A theoretical evaluation. Environ. Entomol. 1993, 22, 519–531. [Google Scholar] [CrossRef]
- Silva, R.; Fernandes, D.; Pedroso, E.; Cividanes, F.; Perioto, N. First report of Aphelinus semiflavus Howard, 1908 (Hymenoptera: Aphelinidae) in Brazil. Braz. J. Biol. 2011, 71, 217–218. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.; Shekhawat, G.S. Anethum graveolens: An Indian traditional medicinal herb and spice. Pharm. Rev. 2010, 4, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Balzan, M.V.; Wäckers, F.L. Flowers to selectively enhance the fitness of a host-feeding parasitoid: Adult feeding by Tuta absoluta and its parasitoid Necremnus artynes. Biol. Control 2013, 67, 21–31. [Google Scholar] [CrossRef]
- Tutin, T.G.E.A. (Ed.) Flora Europaea 1-5; Cambridge University Press: Cambridge, UK, 1964–1980.
- Michener, C.D. The Bees of the World; Johns Hopkins University Press: Baltimore, MA, USA, 2007. [Google Scholar]
- Collins, A.G. Key to the Genera of British Bees. 2012. Available online: https://www.bwars.com/sites/www.bwars.com/files/diary_downloads/Britain%27s_Bees_Chapter_4_Keys_to_Genera.pdf (accessed on 20 July 2020).
- Graham, M.W.R.D.V. The Pteromalidae of north-western Europe (Hymenoptera: Chalcidoidea). Bull. Nat. Hist. Mus. (Entomol.) Suppl. 1969, 16, 1–908. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Margin | Shannon’s Diversity Index (H) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total Arthropods | Hymenoptera Parasitoids | |||||||||||
| WT | Mean 21/5–15/7 | Mean 17/6 &15/7 | WT | Mean WT/Margin (21/5–15/7) | Mean WST/Margin (17/6 & 15/7) | |||||||
| WST | WST | |||||||||||
| 21/5 | 3/6 | 23/6 | 15/7 | 21/5 | 3/6 | 23/6 | 15/7 | |||||
| WM | 1.96 ± 0.06 | 1.91 ± 0.04 | 2.57 ± 0.08 | 2.60 ± 0.07 | 2.26 ± 0.08 A | 2.58 ±0.05 A | 1.77 ± 0.18 | 0.59 ± 0.34 | 1.35 ± 0.21 | 1.67 ± 0.09 | 1.34 ± 0.15 A | 1.51 ± 0.12 |
| NV1 | 1.51 ± 0.25 | 1.66 ± 0.29 | 2.46 ± 0.03 | 1.96 ± 0.16 | 1.90 ± 0.14 B | 2.21 ±0.13 B | 0.94 ± 0.23 | 0.23 ± 0.23 | 1.70 ± 0.22 | 0.84 ±0.26 | 0.93 ± 0.19 B | 1.28 ± 0.24 |
| NV2 | 1.80 ± 0.03 | 1.97 ± 0.18 | 2.40 ± 0.11 | 1.98 ± 0.11 | 2.04 ± 0.08 B | 2.19 ±0.12 B | 1.04 ± 0.04 | 0.66 ± 0.38 | 1.62 ± 0.14 | 0.89 ± 0.21 | 1.05 ± 0.15 AB | 1.26 ± 0.20 |
| SM | 2.21 ± 0.26 | 2.16 ± 0.20 | 2.19 ±0.15 B | 16.7 ± 3.4 | 12.0 ± 2.1 | 0.89 ± 0.30 | ||||||
| Mean WT/date | 1.79 ± 0.08 c | 1.86 ± 0.09 c | 2.49 ± 0.05 a | 2.25 ± 0.11 b | 1.34 ± 0.15 a | 0.51 ±0.19 b | 1.52 ± 0.12 a | 1.23 ± 0.15 a | ||||
| Mean WST/date | 2.43 ±0.07 b | 2.23 ±0.09 a | 0.99 ± 0.53 | 0.80 ± 0.41 | ||||||||
| WT 2-way ANOVA | Mixture: F2,32 = 9.94, p = 0.0004; Date: F3,32 = 21.33, p < 0.0001; Mixture × Date: F6,32 = 1.99, p = 0.0966 | Mixture: F2,32 = 3.53, p = 0.0413; Date: F3,32 = 10.16, p < 0.0001; Mixture × Date: F6,32 = 2.12, p = 0.0788 | ||||||||||
| WST 2-way ANOVA | Mixture: F3,20 = 5.57, p = 0.0060; Date: F1,20 = 6.55, p = 0.0187; Mixture × Date: F6,20 = 2.13, p = 0.1288 | Mixture: F3,20 = 1.99, p = 0.1475; Date: F1,20 = 3.71, p = 0.0686; Mixture × Date: F6,20 = 2.46, p = 0.0928 | ||||||||||
| Seed Weight (%) | Seeds/Species (%) | ||||
|---|---|---|---|---|---|
| Family | Species | WM | SM | WM | SM |
| Apiaceae | Anethum graveolens | 1.97 | 33 | ||
| Coriandrum sativum | 4 | 7 | |||
| Petroselinum crispum * | 17 | 54 | |||
| Asteraceae | Glebionis coronaria | 7 | |||
| Matricaria chamomilla* | 0.03 | 7 | |||
| Tagetes patula* | 5 | 31 | |||
| Fabaceae | Lathyrus sativus | 80 | 13 | ||
| Vicia villosa | 2 | 1 | |||
| Polygonaceae | Fagopyrum esculentum | 68 | 11 | ||
| Boraginaceae | Phacelia tanacetifolia | 15 | 36 | ||
| 6-May | 21-May | 3-June | 17-June | 23-June | 15-July | |
|---|---|---|---|---|---|---|
| WM | F, P | F, P, B | F, P, B | F, P | B | F, P, B |
| SM | F, P | B | F, P, B | |||
| NV1 | F, P | F, P, B | F, P, B | F, P | B | F, P, B |
| NV2 | F, P | F, P, B | F, P, B | F, P | B | F, P, B |
| T | F, P | F, P |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kati, V.; Karamaouna, F.; Economou, L.; Mylona, P.V.; Samara, M.; Mitroiu, M.-D.; Barda, M.; Edwards, M.; Liberopoulou, S. Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropods in a Tomato Field Margin. Plants 2021, 10, 1003. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051003
Kati V, Karamaouna F, Economou L, Mylona PV, Samara M, Mitroiu M-D, Barda M, Edwards M, Liberopoulou S. Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropods in a Tomato Field Margin. Plants. 2021; 10(5):1003. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051003
Chicago/Turabian StyleKati, Vaya, Filitsa Karamaouna, Leonidas Economou, Photini V. Mylona, Maria Samara, Mircea-Dan Mitroiu, Myrto Barda, Mike Edwards, and Sofia Liberopoulou. 2021. "Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropods in a Tomato Field Margin" Plants 10, no. 5: 1003. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10051003