
Morphological and Chemical Factors Related to Western Flower Thrips Resistance in the Ornamental Gladiolus


Abstract
:1. Introduction
2. Results
2.1. Differences in Resistance to Thrips
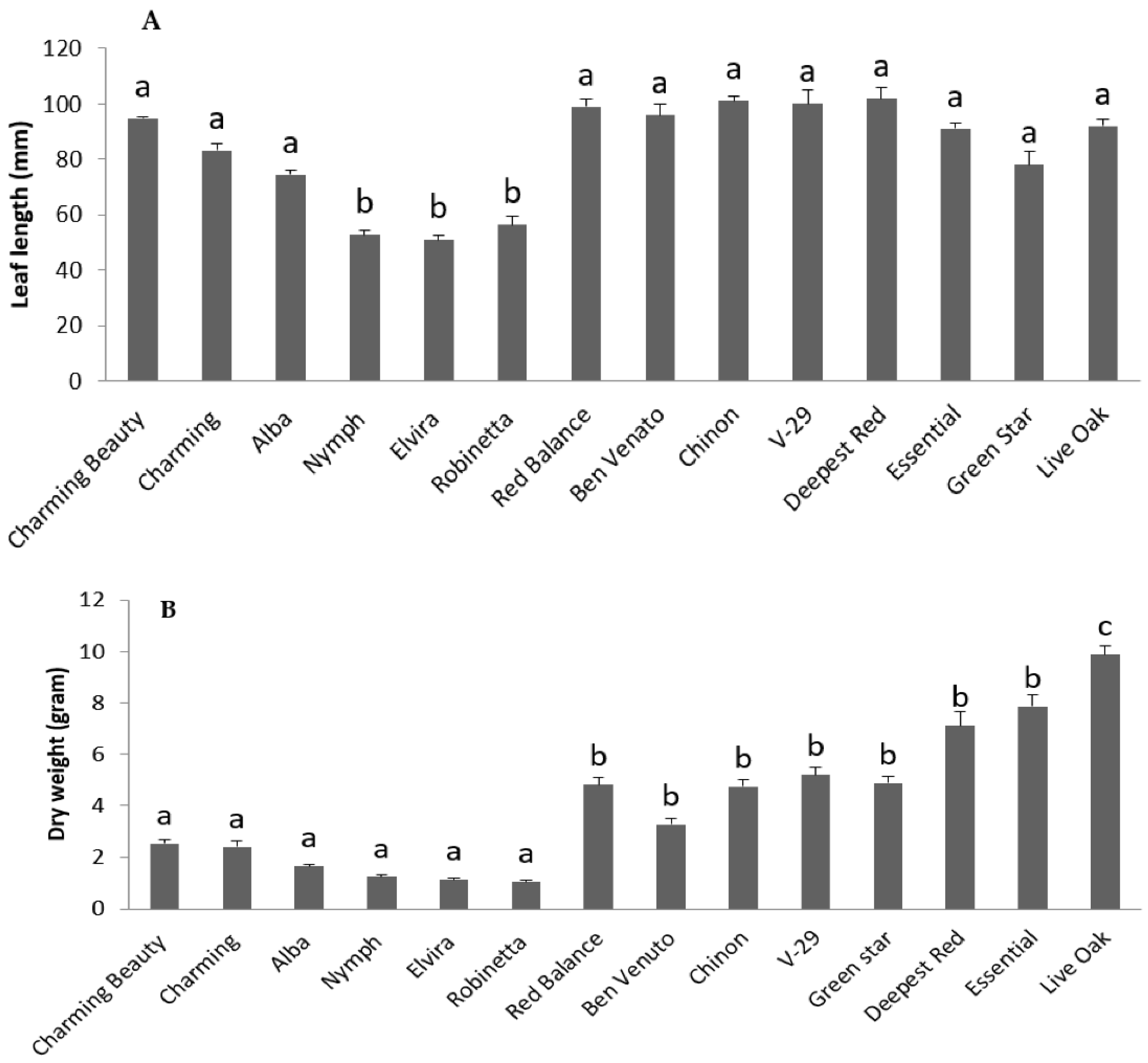
2.1.1. Differences in Morphological Traits
2.1.2. The Relationship between Thrips Damage and Morphological Characteristics
2.2. Chemical Study
2.2.1. Differences in the Effect of Six Leaf Extracts on Thrips Mortality
2.2.2. Metabolic Profiling
2.2.3. Signals Related to Resistance
2.2.4. Signals Related to Susceptibility
2.2.5. Correlations between Chemical and Morphological Characteristics Related to Thrips Resistance
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Resistance to Thrips
4.3. Morphological Measurements
4.4. In Vitro Thrips Bioassay
4.5. Metabolic Profiling
4.5.1. Extraction of Plant Materials for NMR Metabolomics
4.5.2. NMR Analysis
4.5.3. Data Processing
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar]
- Riddick, E.W.; Simmons, A.M. Do plant trichomes cause more harm than good to predatory insects? Pest Manag. Sci. 2014, 70, 1655–1665. [Google Scholar] [CrossRef]
- Duarte-Silva, A.G.; Carvalho-Silva, M.; Camara, P.E.A.S. Morphology and development of leaf papillae in the Pilotrichaceae. Acta Bot. Bras. 2013, 27, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Maluf, W.R.; Campos, G.A.; das Grasas Cardoso, M. Relationships between trichome types and spider mite (Tetranychus evansi) repellence in tomatoes with respect to foliar zingiberene contents. Euphytica 2001, 121, 73–80. [Google Scholar] [CrossRef]
- Underwood, W. The plant cell wall: A dynamic barrier against pathogen invasion. Front. Plant Sci. 2012, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prüm, B.; Bohn, H.F.; Seidel, R.; Rubach, S.; Speck, T. Plant surfaces with cuticular folds and their replicas: Influence of microstructuring and surface chemistry on the attachment of a leaf beetle. Acta Biomater. 2013, 9, 6360–6368. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Lucini, T.; Faria, M.V.; Rohde, C.; Resende, J.; de Oliveira, J.R.F. Acylsugar and the role of trichomes in tomato genotypes resistance to Tetranychus urticae. Arthropod. Plant Interact. 2015, 9, 45–53. [Google Scholar] [CrossRef]
- Mirnezhad, M.; Romero-González, R.R.; Leiss, K.A.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Metabolomic analysis of host plant resistance to thrips in wild and cultivated tomatoes. Phytochem. Anal. 2010, 21, 110–117. [Google Scholar] [CrossRef]
- Munien, P.; Naidoo, Y.; Naidoo, G. Micromorphology, histochemistry and ultrastructure of the foliar trichomes of Withania somnifera (L.) Dunal (Solanaceae). Planta 2015, 242, 1107–1122. [Google Scholar] [CrossRef]
- Wakte, K.V.; Nadaf, A.B.; Krishnan, S.; Thengane, R.J. Studies on lower epidermal papillae, the site of storage of basmati rice aroma compounds in Pandanus amaryllifolius Roxb. Curr. Sci. 2007, 93, 238–242. [Google Scholar]
- Ramalho-Santos, M.; Pissarra, J.; Veríssimo, P.; Pereira, S.; Salema, R.; Pires, E.; Faro, C.J. Cardosin A, an abundant aspartic proteinase, accumulates in protein storage vacuoles in the stigmatic papillae of Cynara cardunculus L. Planta 1997, 203, 204–212. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Shipp, J.L. Influence of plant species and plant growth stage on Frankliniella occidentalis pupation behaviour in greenhouse ornamentals. J. Appl. Entomol. 2008, 132, 86–88. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agric. For. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Lewis, T. Thrips as Crop Pests; Cab International: Wallingford, UK, 1997; ISBN 0851991785. [Google Scholar]
- Capinera, J.L. Encyclopedia of Entomology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004; ISBN 1402062427. [Google Scholar]
- Ogada, P.A.; Poehling, H.-M. Sex-specific influences of Frankliniella occidentalis (western flower thrips) in the transmission of Tomato spotted wilt virus (Tospovirus). J. Plant Dis. Prot. 2015, 122, 264–274. [Google Scholar] [CrossRef]
- Denmark, H.A.; Price, J.F. Gladiolus thrips, thrips simplex (Morison) (Insecta: Thysanoptera: Thripidae). DPI Entomol. Circ. 1998, 322, 1–3. [Google Scholar]
- Childers, C.C. Feeding and oviposition injuries to plants. In Invasive Species Compendium; CAB International: Wallingford, UK, 1997; pp. 505–537. [Google Scholar]
- Kirk, W.D.J. The pest and vector from the West: Frankliniella occidentalis BT—Thrips and Tospoviruses. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; Australian National Insect Collection: Canberra, Australia, 2002; pp. 33–42. [Google Scholar]
- Mouden, S.; Leiss, K.A. Host plant resistance to thrips (Thysanoptera: Thripidae)—Current state of art and future research avenues. Curr. Opin. Insect Sci. 2021, 45, 28–34. [Google Scholar] [CrossRef]
- Leiss, K.A.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. An overview of NMR-based metabolomics to identify secondary plant compounds involved in host plant resistance. Phytochem. Rev. 2011, 10, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Leiss, K.A.; Choi, Y.H.; Abdel-Farid, I.B.; Verpoorte, R.; Klinkhamer, P.G.L. NMR metabolomics of thrips (Frankliniella occidentalis) resistance in Senecio hybrids. J. Chem. Ecol. 2009, 35, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Identification of chlorogenic acid as a resistance factor for thrips in chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef] [Green Version]
- Bac-Molenaar, J.A.; Mol, S.; Verlaan, M.G.; van Elven, J.; Kim, H.K.; Klinkhamer, P.G.L.; Leiss, K.A.; Vrieling, K. Trichome independent resistance against western flower thrips in tomato. Plant Cell Physiol. 2019, 60, 1011–1024. [Google Scholar] [CrossRef] [Green Version]
- Leiss, K.A.; Cristofori, G.; van Steenis, R.; Verpoorte, R.; Klinkhamer, P.G.L. An eco-metabolomic study of host plant resistance to Western flower thrips in cultivated, biofortified and wild carrots. Phytochemistry 2013, 93, 63–70. [Google Scholar] [CrossRef]
- Maharijaya, A.; Vosman, B.; Verstappen, F.; Steenhuis-Broers, G.; Mumm, R.; Purwito, A.; Visser, R.G.F.; Voorrips, R.E. Resistance factors in pepper inhibit larval development of thrips (F rankliniella occidentalis). Entomol. Exp. Appl. 2012, 145, 62–71. [Google Scholar] [CrossRef]
- Macel, M.; Visschers, I.G.S.; Peters, J.L.; Kappers, I.F.; de Vos, R.C.H.; van Dam, N.M. Metabolomics of thrips resistance in pepper (Capsicum spp.) reveals monomer and dimer acyclic diterpene glycosides as potential chemical defenses. J. Chem. Ecol. 2019, 45, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Shelton, A.M. Evaluation of onion cultivars for resistance to onion thrips (Thysanoptera: Thripidae) and Iris yellow spot virus. J. Econ. Entomol. 2010, 103, 925–937. [Google Scholar] [CrossRef]
- Benschop, M.; Kamenetsky, R.; Le Nard, M.; Okubo, H.; De Hertogh, A. The global flower bulb industry: Production, utilization, research. Hortic. Rev. 2010, 36, 1. [Google Scholar]
- Bailey, N.J.C.; Wang, Y.; Sampson, J.; Davis, W.; Whitcombe, I.; Hylands, P.J.; Croft, S.L.; Holmes, E. Prediction of anti-plasmodial activity of Artemisia annua extracts: Application of 1H NMR spectroscopy and chemometrics. J. Pharm. Biomed. Anal. 2004, 35, 117–126. [Google Scholar] [CrossRef]
- de Jager, C.M.; Butot, R.P.T.; Klinkhamer, P.G.L.; de Jong, T.J.; Wolff, K.; van der Meijden, E. Genetic variation in chrysanthemum for resistance to Frankliniella occidentalis. Entomol. Exp. Appl. 1995, 77, 277–287. [Google Scholar] [CrossRef]
- Kos, S.P.; Klinkhamer, P.G.L.; Leiss, K.A. Cross-resistance of chrysanthemum to western flower thrips, celery leafminer, and two-spotted spider mite. Entomol. Exp. Appl. 2014, 151, 198–208. [Google Scholar] [CrossRef]
- Gaum, W.G.; Giliomee, J.H.; Pringle, K.L. Resistance of some rose cultivars to the western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Bull. Entomol. Res. 1994, 84, 487–492. [Google Scholar] [CrossRef]
- Scott Brown, A.S.; Simmonds, M.S.J. Leaf morphology of hosts and nonhosts of the thrips Heliothrips haemorrhoidalis (Bouche). Bot. J. Linn. Soc. 2006, 152, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Boughton, A.J.; Hoover, K.; Felton, G.W. Methyl jasmonate application induces increased densities of glandular trichomes on tomato, lycopersicon esculentum. J. Chem. Ecol. 2005, 31, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Yadwad, A.; Sridevi, O.; Salimath, P.M. Leaf trichomes-based resistance in chilli (Capsicum annuum) to thrips and mites. Indian J. Agric. Sci. 2008, 78, 518–521. [Google Scholar]
- Ben-Mahmoud, S.; Anderson, T.; Chappell, T.M.; Smeda, J.R.; Mutschler, M.A.; Kennedy, G.G.; De Jong, D.M.; Ullman, D.E. A thrips vector of tomato spotted wilt virus responds to tomato acylsugar chemical diversity with reduced oviposition and virus inoculation. Sci. Rep. 2019, 9, 17157. [Google Scholar] [CrossRef]
- Haralampidis, K.; Trojanowska, M.; Osbourn, A.E. Biosynthesis of triterpenoid saponins in plants. In History and Trends in Bioprocessing and Biotransformation; Springer: Berlin/Heidelberg, Germany, 2002; pp. 31–49. [Google Scholar]
- Capitani, D.; Sobolev, A.P.; Tomassini, A.; Sciubba, F.; De Salvador, F.R.; Mannina, L.; Delfini, M. Peach fruit: Metabolic comparative analysis of two varieties with different resistances to insect attacks by NMR spectroscopy. J. Agric. Food Chem. 2012, 61, 1718–1726. [Google Scholar] [CrossRef]
- Dhillon, M.K.; Kumar, S. Amino acid profiling of Sorghum bicolor vis-à-vis Chilo partellus (Swinhoe) for biochemical interactions and plant resistance. Arthropod. Plant. Interact. 2017, 11, 537–550. [Google Scholar] [CrossRef]
- Bernards, M.A. Plant natural products: A primer. Can. J. Zool. 2010, 88, 601–614. [Google Scholar] [CrossRef]
- Ferracini, C.; Curir, P.; Dolci, M.; Lanzotti, V.; Alma, A. Aesculus pavia foliar saponins: Defensive role against the leafminer Cameraria ohridella. Pest Manag. Sci. 2010, 66, 767–772. [Google Scholar] [CrossRef]
- Kuzina, V.; Ekstrom, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of defense compounds in barbarea vulgaris against the herbivore phyllotreta nemorum by an ecometabolomic approach. Plant Physiol. 2009, 151, 1977–1990. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.K.; Nagao, T.; Okabe, H.; Shinoda, T. Resistance in the plant, barbarea vulgaris, and counter-adaptations in flea beetles mediated by saponins. J. Chem. Ecol. 2010, 36, 277–285. [Google Scholar] [CrossRef]
- Badenes-Pérez, F.R.; López-Pérez, J.A. Resistance and susceptibility to powdery mildew, root-knot nematode, and western flower thrips in two types of winter cress (Brassicaceae). Crop Prot. 2018, 110, 41–47. [Google Scholar] [CrossRef]
- Applebaum, S.W.; Marco, S.; Birk, Y. Saponins as possible factors of resistance of legume seeds to the attack of insects. J. Agric. Food Chem. 1969, 17, 618–622. [Google Scholar] [CrossRef]
- Ishaaya, I. Nutritional and allelochemic insect-plant interactions relating to digestion and food intake: Some examples. In Insect-Plant Interaction; Springer: Berlin/Heidelberg, Germany, 1986; pp. 191–223. [Google Scholar]
- Shany, S.; Birk, Y.; Gestetner, B.; Bondi, A. Preparation, characterisation and some properties of saponins from lucerne tops and roots. J. Sci. Food Agric. 1970, 21, 131–135. [Google Scholar] [CrossRef]
- De Geyter, E.; Lambert, E.; Geelen, D.; Smagghe, G. Novel advances with plant saponins as natural insecticides to control pest insects. Pest Technol. 2007, 1, 96–105. [Google Scholar]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epidermal Cell Length (µm) | Mesophyll Length (µm) | Density of Papillae (per 2100 µm2) | Leaf Length (cm) | Dry Mass (g) | |
|---|---|---|---|---|---|
| Ln silver damage | r = 0.596 p = 0.024 | r = 0.603 p = 0.022 | r = −0.628 p = 0.016 | r = 0.320 p = 0.264 | r = −0.222 p = 0.445 |
| Epidermal cell length | r = 0.931 p = 0.000 | r = −0.873 p = 0.000 | r = 0.704 p = 0.005 | r = 0.310 p = 0.281 | |
| Mesophyll length | r = −0.909 p = 0.000 | r = 0.777 p = 0.001 | r = 0.315 p = 0.273 | ||
| Density of papillae | r = −0.669 p = 0.009 | r = −0.389 p = 0.170 | |||
| Leaf length | r = 0.441 p = 0.114 |
| Valine | Sucrose | Glucose | EGC | Gallic Acid | |
|---|---|---|---|---|---|
| Ln damage | r = −0.100 p = 0.734 | r = 0.083 p = 0.779 | r = 0.265 p = 0.360 | r = 0.404 p = 0.152 | r = 0.313 p = 0.276 |
| Papillae | r = −0.034 p = 0.907 | r = −0.117 p = 0.692 | r = −0.442 p = 0.114 | r = −0.275 p = 0.342 | r = −0.019 p = 0.950 |
| Valine | r = 0.345 p = 0.227 | r = 0.096 p = 0. 743 | r = −0.594 * p = 0. 025 | r = −0.403 p = 0.153 | |
| Sucrose | r = −0.123 p = 0.676 | r = −0.576 * p = 0.031 | r = −0.058 p = 0.845 | ||
| Glucose | r = −0.047 p = 0.874 | r = 0.139 p = 0.636 | |||
| EGC | r = 0.074 p = 0.801 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahyuni, D.S.C.; Choi, Y.H.; Leiss, K.A.; Klinkhamer, P.G.L. Morphological and Chemical Factors Related to Western Flower Thrips Resistance in the Ornamental Gladiolus. Plants 2021, 10, 1384. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071384
Wahyuni DSC, Choi YH, Leiss KA, Klinkhamer PGL. Morphological and Chemical Factors Related to Western Flower Thrips Resistance in the Ornamental Gladiolus. Plants. 2021; 10(7):1384. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071384
Chicago/Turabian StyleWahyuni, Dinar S. C., Young Hae Choi, Kirsten A. Leiss, and Peter G. L. Klinkhamer. 2021. "Morphological and Chemical Factors Related to Western Flower Thrips Resistance in the Ornamental Gladiolus" Plants 10, no. 7: 1384. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071384