
How Leaf Vein and Stomata Traits Are Related with Photosynthetic Efficiency in Falanghina Grapevine in Different Pedoclimatic Conditions

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Environmental Data Characterization
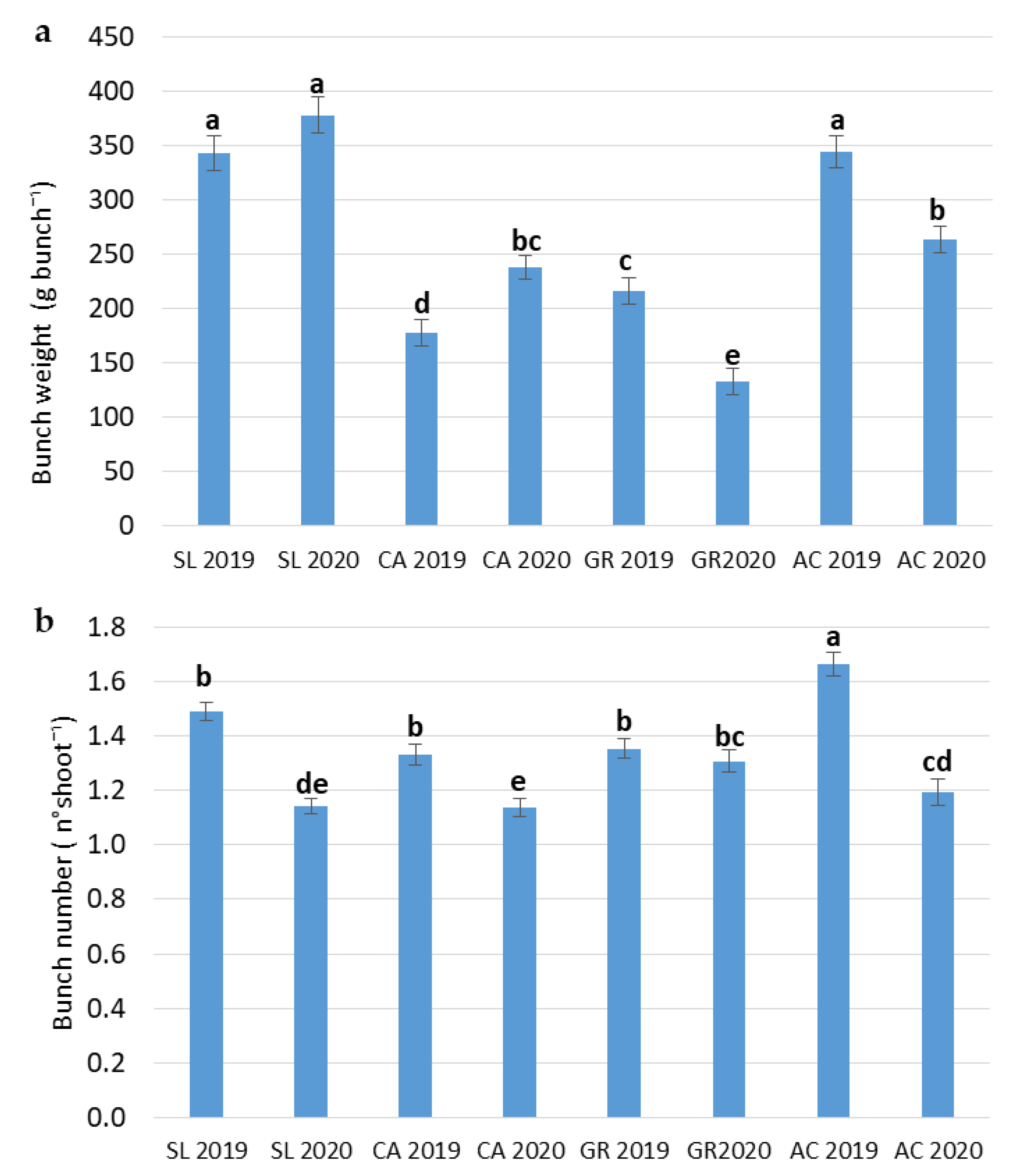
2.2. Growth and Production Parameters
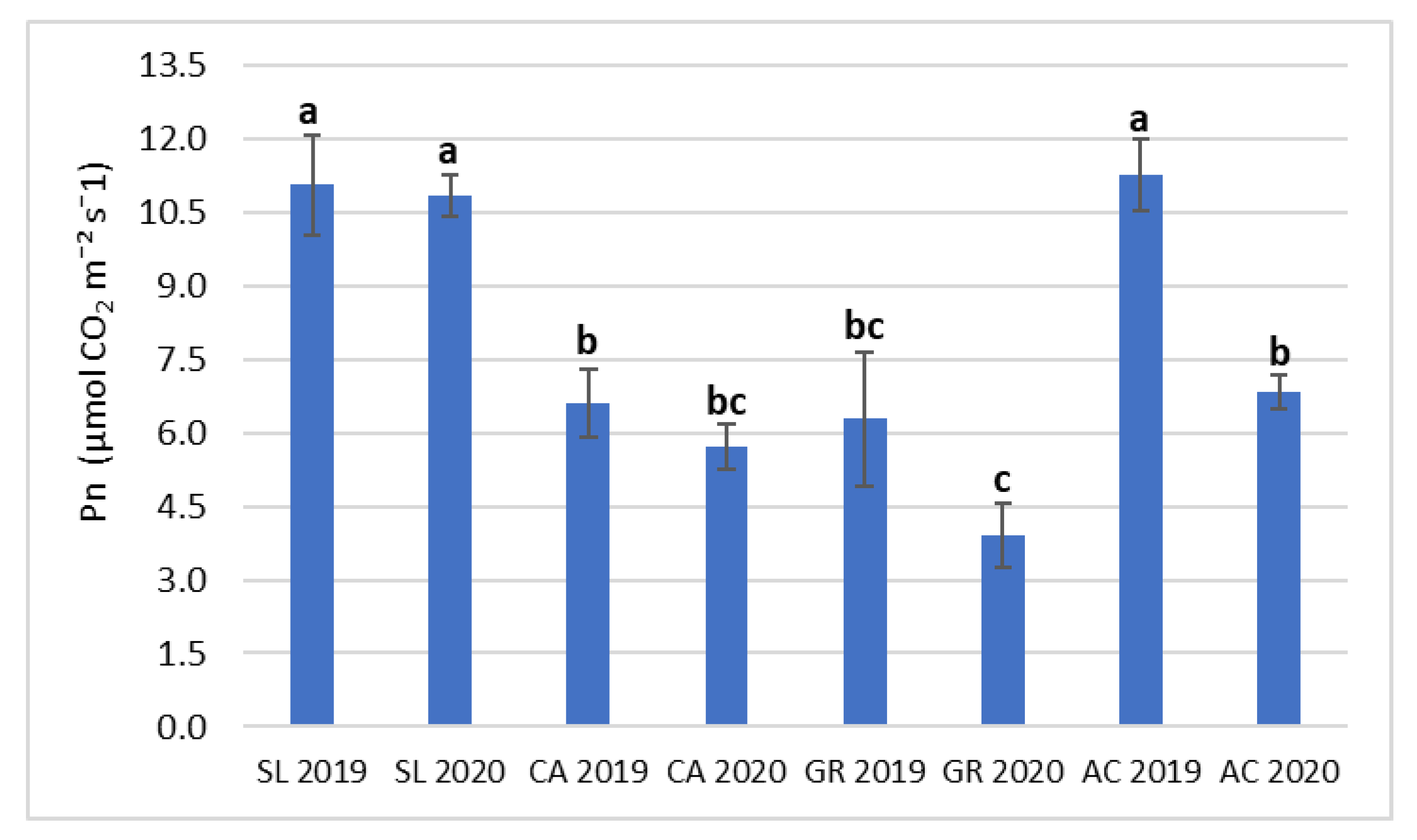
2.3. Gas-Exchange and Chlorophyll a Fluorescence
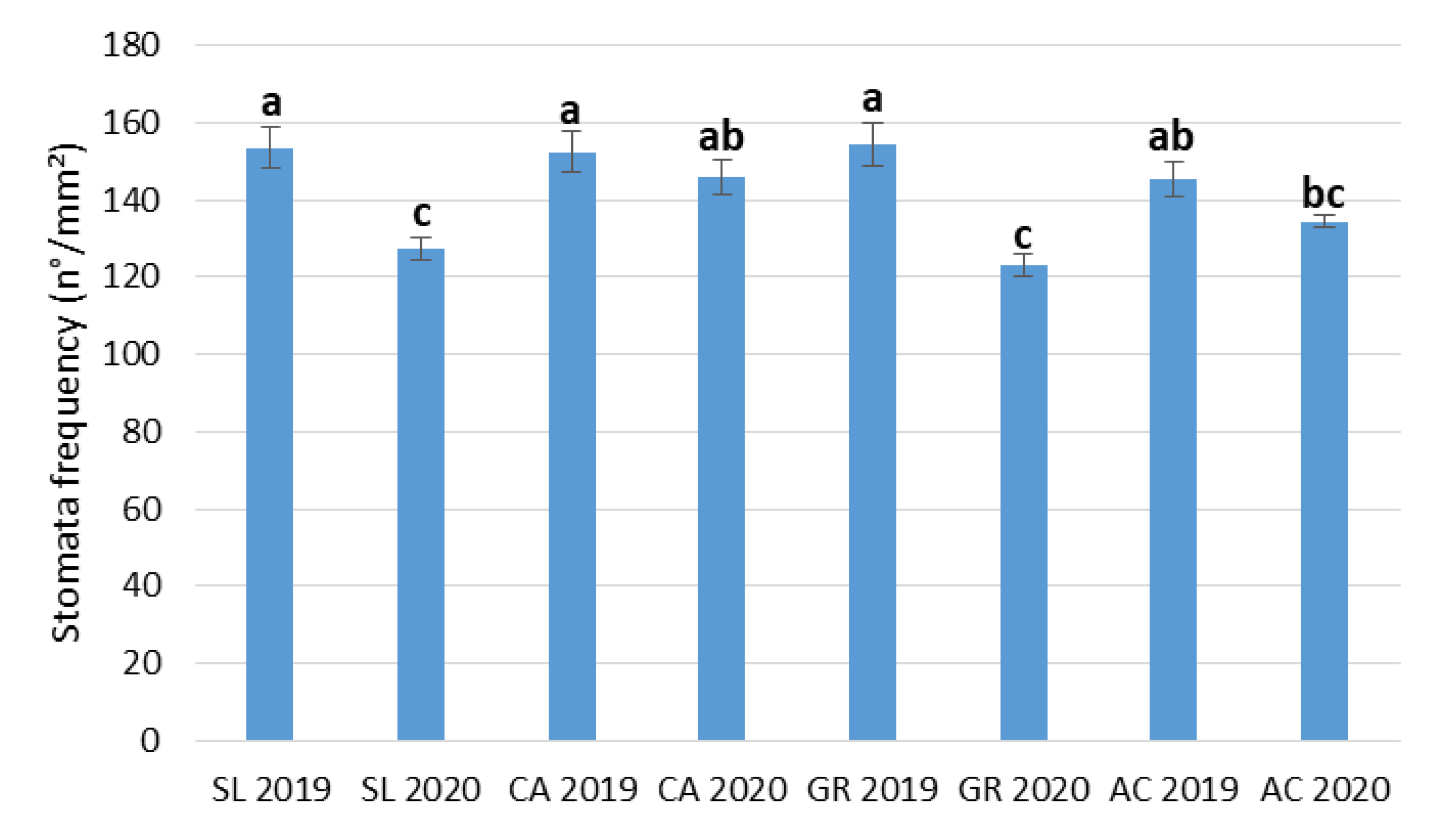
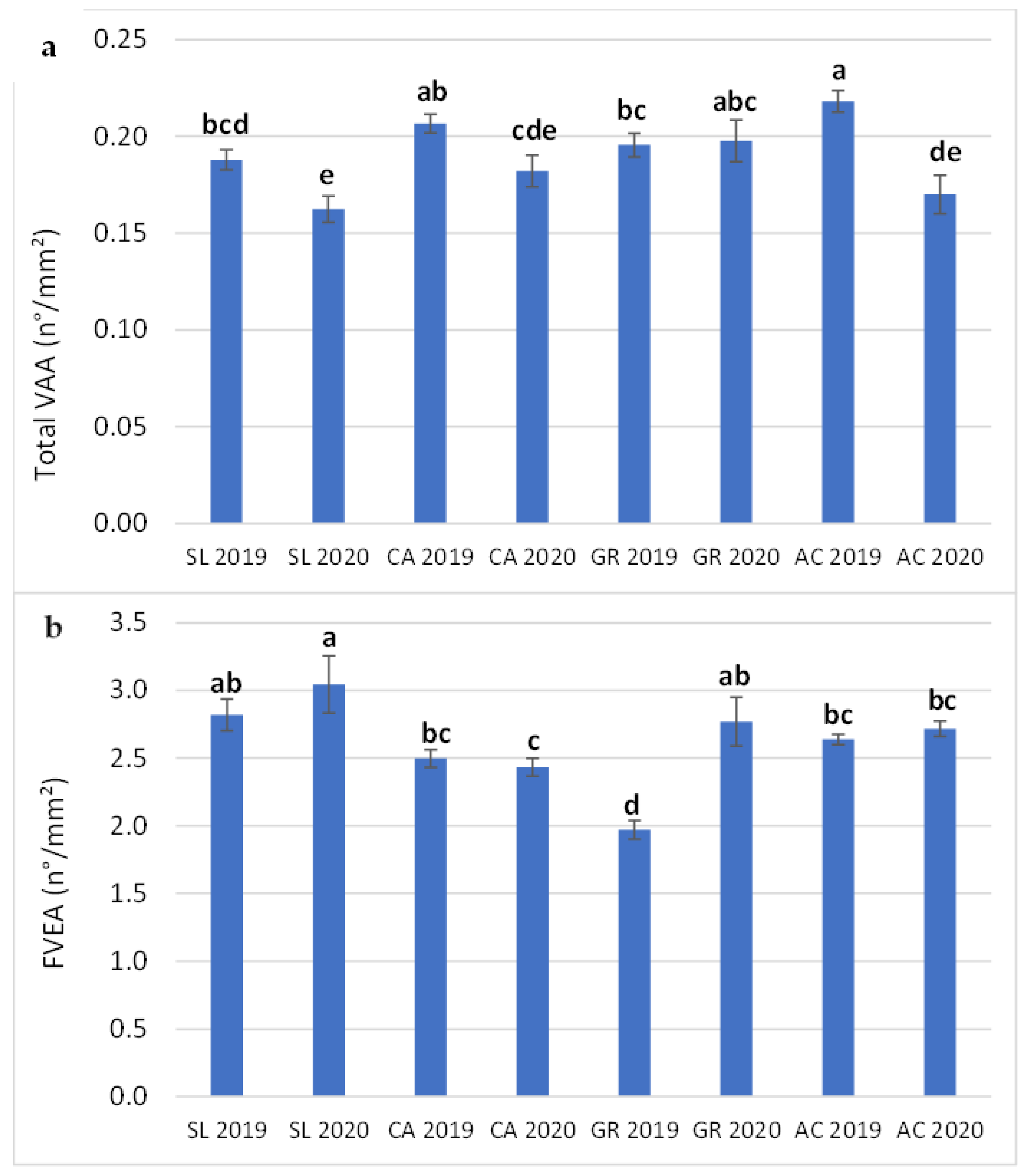
2.4. Stomata and Vein Traits
3. Discussion
4. Materials and Methods
4.1. Study Area and Vineyard Characteristics
4.2. Biometry and Yield
4.3. Gas-Exchange and Chlorophyll a Fluorescence Emission Measurements
4.4. Microscopy and Digital Image Analysis
- minor vein length per unit area (Minor VLA) = sum of vein lengths of third or higher orders of veins divided by the difference between the area imaged and the area occupied by the second-order veins (mm/mm2);
- major vein length per unit area (Major VLA) = sum of vein lengths of second-order veins divided by the area imaged (mm/mm2);
- minor vein area per unit area (Minor VAA) = sum of vein areas of third or higher orders of veins divided by the difference between the area imaged and the area occupied by the second-order veins (mm/mm2);
- major vein area per unit area (Major VAA) = sum of vein areas of second-order veins divided by the area imaged (mm/mm2);
- total vein length per area (Total VLA) = sum of vein lengths of all order veins divided by the area imaged (mm/mm2);
- total vein area per unit area (Total VAA) = sum of vein areas of all order veins divided by the area imaged (mm2/mm2);
- free vein endings per unit area (FVEA) = number of vein endings divided by the area imaged (n°/mm2).
4.5. Statistical Analysis of Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2021: Sixth Assessment Report; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2021. [Google Scholar]
- Webb, L.B.; Watterson, I.; Bhend, J.; Whetton, P.H.; Barlow, E.W.R. Global Climate Analogues for Winegrowing Regions in Future Periods: Projections of Temperature and Precipitation. Aust. J. Grape Wine Res. 2013, 19, 331–341. [Google Scholar] [CrossRef]
- Lionello, P.; Congedi, L.; Reale, M.; Scarascia, L.; Tanzarella, A. Sensitivity of Typical Mediterranean Crops to Past and Future Evolution of Seasonal Temperature and Precipitation in Apulia. Reg. Environ. Change 2014, 14, 2025–2038. [Google Scholar] [CrossRef]
- Yang, C.; Menz, C.; Fraga, H.; Costafreda-Aumedes, S.; Leolini, L.; Ramos, M.C.; Molitor, D.; Leeuwen, C.V.; Santos, J.A. Assessing the Grapevine Crop Water Stress Indicator over the Flowering-Veraison Phase and the Potential Yield Lose Rate in Important European Wine Regions. Agric. Water Manag. 2022, 261, 107349. [Google Scholar] [CrossRef]
- Cogato, A.; Wu, L.; Jewan, S.Y.Y.; Meggio, F.; Marinello, F.; Sozzi, M.; Pagay, V. Evaluating the Spectral and Physiological Responses of Grapevines (Vitis Vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies. Agronomy 2021, 11, 1940. [Google Scholar] [CrossRef]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; Leeuwen, C.V. Influence of Vineyard Location and Vine Water Status on Fruit Maturation of Nonirrigated Cv. Agiorgitiko (Vitis Vinifera L.). Effects on Wine Phenolic and Aroma Components. J. Agric. Food Chem. 2006, 54, 5077–5086. [Google Scholar] [CrossRef]
- Pagay, V.; Zufferey, V.; Lakso, A.N. The Influence of Water Stress on Grapevine (Vitis vinifera L.) Shoots in a Cool, Humid Climate: Growth, Gas Exchange and Hydraulics. Funct. Plant Biol. 2016, 43, 827–837. [Google Scholar] [CrossRef]
- Micco, V.D.; Aronne, G. Morpho-Anatomical Traits for Plant Adaptation to Drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. ISBN 9783642326530. [Google Scholar]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Ferreira, H.F.; Correia, C.M. Changes in Growth, Gas Exchange, Xylem Hydraulic Properties and Water Use Efficiency of Three Olive Cultivars under Contrasting Water Availability Regimes. Environ. Exp. Bot. 2007, 60, 183–192. [Google Scholar] [CrossRef]
- Huang, L.; Li, Z.; Liu, Q.; Pu, G.; Zhang, Y.; Li, J. Research on the Adaptive Mechanism of Photosynthetic Apparatus under Salt Stress: New Directions to Increase Crop Yield in Saline Soils. Ann. Appl. Biol. 2019, 175, 1–17. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Martinez-Luscher, J.; Girardello, R.C.; Kostaki, E.; Oberholster, A.; Kurtural, S.K. Shifts in the Phenolic Composition and Aromatic Profiles of Cabernet Sauvignon (Vitis vinifera L.) Wines Are Driven by Different Irrigation Amounts in a Hot Climate. Food Chem. 2022, 371, 131163. [Google Scholar] [CrossRef]
- Bonfante, A.; Monaco, E.; Langella, G.; Mercogliano, P.; Bucchignani, E.; Manna, P.; Terribile, F. A Dynamic Viticultural Zoning to Explore the Resilience of Terroir Concept under Climate Change. Sci. Total Environ. 2018, 624, 294–308. [Google Scholar] [CrossRef]
- Cirillo, C.; Arena, C.; Rouphael, Y.; Caputo, R.; Amitrano, C.; Petracca, F.; Francesco, S.D.; Vitale, E.; Erbaggio, A.; Bonfante, A.; et al. Counteracting the Negative Effects of Copper Limitations Through the Biostimulatory Action of a Tropical Plant Extract in Grapevine Under Pedo-Climatic Constraints. Front. Environ. Sci. 2021, 9, 587550. [Google Scholar] [CrossRef]
- Micco, V.D.; Zalloni, E.; Battipaglia, G.; Erbaggio, A.; Scognamiglio, P.; Caputo, R.; Cirillo, C. Rootstock Effect on Tree-Ring Traits in Grapevine under a Climate Change Scenario. IAWA J. 2018, 39, 145–155. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Frole, K. Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 2006, 87, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Feild, T.S.; Jordan, G.J. Leaf Maximum Photosynthetic Rate and Venation Are Linked by Hydraulics. Plant Physiol. 2007, 144, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Rockwell, F.E.; Holbrook, N.M. Leaf Hydraulic Architecture and Stomatal Conductance: A Functional Perspective. Plant Physiol. 2017, 174, 1996–2007. [Google Scholar] [CrossRef] [Green Version]
- Franks, P.J.; Leitch, I.J.; Ruszala, E.M.; Hetherington, A.M.; Beerling, D.J. Physiological Framework for Adaptation of Stomata to CO2 from Glacial to Future Concentrations. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 537–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salleo, S.; Nardini, A.; Pitt, F.; Gullo, M.A.L. Xylem Cavitation and Hydraulic Control of Stomatal Conductance in Laurel (Laurus nobiris L.). Plant Cell. Environ. 2000, 23, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Poorter, H. Inherent Variation in Growth Rate Between Higher Plants: A Search for Physiological Causes and Ecological Consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Jordan, G.J. Water Supply and Demand Remain Balanced during Leaf Acclimation of Nothofagus Cunninghamii Trees. New Phytol. 2011, 192, 437–448. [Google Scholar] [CrossRef]
- Murphy, M.R.C.; Jordan, G.J.; Brodribb, T.J. Differential Leaf Expansion Can Enable Hydraulic Acclimation to Sun and Shade. Plant Cell Environ. 2012, 35, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Gerzon, E.; Biton, I.; Yaniv, Y.; Zemach, H.; Netzer, Y.; Schwartz, A.; Fait, A.; Ben-Ari, G. Grapevine Anatomy as a Possible Determinant of Isohydric or Anisohydric Behavior. Am. J. Enol. Vitic. 2015, 66, 340–347. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T. Variability among Species of Stomatal Control under Fluctuating Soil Water Status and Evaporative Demand: Modelling Isohydric and Anisohydric Behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Gago, P.; Conejero, G.; Martínez, M.C.; This, P.; Verdeil, J.L. Comparative Anatomy and Morphology of the Leaves of Grenache Noir and Syrah Grapevine Cultivars. S. Afr. J. Enol. Vitic. 2019, 40, 1–9. [Google Scholar] [CrossRef]
- Jones, H.G.; Sutherland, R.A. Stomatal control of xylem embolism. Plant Cell Environ. 1991, 14, 607–612. [Google Scholar] [CrossRef]
- MacMillan, P.; Teixeira, G.; Lopes, C.M.; Monteiro, A. The Role of Grapevine Leaf Morphoanatomical Traits in Determining Capacity for Coping with Abiotic Stresses: A Review. Ciência E Técnica Vitivinícola 2021, 36, 75–88. [Google Scholar] [CrossRef]
- Patakas, A.; Kofidis, G.; Bosabalidis, A.M. The relationships between CO2 transfer mesophyll resistance and photosynthetic efficiency in grapevine cultivars. Sci. Hortic. 2003, 97, 255–263. [Google Scholar] [CrossRef]
- Patakas, A.; Stavrakas, D.; Fisarakis, I. Relationship between CO2 assimilation and leaf anatomical characteristics of two grapevine cultivars. Agronomie 2003, 23, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Tomás, M.; Medrano, H.; Brugnoli, E.; Escalona, J.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of mesophyll conductance in grapevine cultivars under water stress conditions in relation to leaf anatomy and water use efficiency. Aust. J. Grape Wine Res. 2014, 20, 272–280. [Google Scholar] [CrossRef]
- Romero, P.; Navarro, J.M.; Ordaz, P.B. Towards a sustainable viticulture: The combination of deficit irrigation strategies and agroecological practices in Mediterranean vineyards. A review and update. Agric. Water Manag. 2022, 259, 107216. [Google Scholar] [CrossRef]
- Damiano, N.; Altieri, S.; Battipaglia, G.; Micco, V.D. Comparing Methods for the Analysis of δ13C in Falanghina Grape Must from Different Pedoclimatic Conditions. Horticulturae 2022, 8, 226. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Bienaimé, D.; Marmottant, P. Revealing Catastrophic Failure of Leaf Networks under Stress. Proc. Natl. Acad. Sci. USA 2016, 113, 4865–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creek, D.; Lamarque, L.J.; Torres-Ruiz, J.M.; Parise, C.; Burlett, R.; Tissue, D.T.; Delzon, S. Xylem Embolism in Leaves Does Not Occur with Open Stomata: Evidence from Direct Observations Using the Optical Visualization Technique. J. Exp. Bot. 2020, 71, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Dayer, S.; Herrera, J.C.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G.A. The Sequence and Thresholds of Leaf Hydraulic Traits Underlying Grapevine Varietal Differences in Drought Tolerance. J. Exp. Bot. 2020, 71, 4333–4344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, W.W.; Watson, A.M.; Mueh, K.E.; Amiard, V.; Turgeon, R.; Ebbert, V.; Logan, B.A.; Combs, A.F.; Demmig-Adams, B. Photosynthetic acclimation in the context of structural constraints to carbon export from leaves. Photosyn. Res. 2007, 94, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Sack, L.; Scoffoni, C. Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytol. 2013, 198, 983–1000. [Google Scholar] [CrossRef] [PubMed]
- Maximov, N.A. The physiological significance of the xeromorphic structure of plants. J. Ecol. 1931, 19, 279–282. [Google Scholar] [CrossRef]
- Grubb, P. A reassessment of the strategies of plants which cope with shortages of resources. Perspect. Plant Ecol. Evol. Syst. 1998, 1, 3–31. [Google Scholar] [CrossRef]
- Scoffoni, C.; Rawls, M.; McKown, A.; Cochard, H.; Sack, L. Decline of Leaf Hydraulic Conductance with Dehydration: Relationship to Leaf Size and Venation Architecture. Plant Physiol. 2011, 156, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Giorio, P.; Basile, A.; Sorrentino, G.; Albrizio, R. Physiological responses of Falanghina grapevines in soils with different water availability in Southern Italy. Acta Hortic. 2007, 754, 235–240. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poni, S.; Bernizzoni, F.; Civardi, S. Response of ‘Sangiovese’ grapevines to partial rootzone drying gas-exchange, growth and grape composition. Scientia Horticulturae 2007, 114, 96–103. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines. Anatomy and Physiology, 1st ed.; Academic Press: Burlington, MA, USA, 2010; ISBN 9780080890487. [Google Scholar]
- Salem-Fnayou, A.B.; Bouamama, B.; Ghorbel, A.; Mliki, A. Investigations on the leaf anatomy and ultrastructure of grapevine (Vitis vinifera) under heat stress. Microsc. Res. Tech. 2011, 74, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Sadras, V.O.; Montoro, A.; Moran, M.A.; Aphalo, P.J. Elevated temperature altered the reaction norms of stomatal conductance in field-grown grapevine. Agric. For. Meteorol. 2012, 165, 35–42. [Google Scholar] [CrossRef]
- Serra, I.; Strever, A.; Myburgh, P.; Schmeisser, M.; Deloire, P.A. Grapevine (Vitis vinifera L. ‘Pinotage’) leaf stomatal size and density as modulated by different rootstocks and scion water status. Acta Hort. 2017, 1157, 177–181. [Google Scholar] [CrossRef]
- Amitrano, C.; Arena, C.; Cirillo, V.; Pascale, S.D.; Micco, V.D. Leaf morpho-anatomical traits in Vigna radiata L. affect plant photosynthetic acclimation to changing vapor pressure deficit. Environ. Exp. Bot. 2021, 186, 104453. [Google Scholar] [CrossRef]
- Vitale, L.; Arena, C.; Santo, A.V.D. Seasonal changes in photosynthetic activity and photochemical efficiency of the Mediterranean shrub Phillyrea angustifolia L. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2012, 146, 443–450. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Santo, A.V.D. Photosynthesis and photoprotective strategies in Laurus nobilis L. and Quercus ilex L. under summer drought and winter cold. Plant Biosyst. 2008, 142, 472–479. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Santo, A.V.D. Paraheliotropism in Robinia pseudoacacia L.: An efficient strategy to optimise photosynthetic performance under natural environmental conditions. Plant Biol. 2008, 10, 194–201. [Google Scholar] [CrossRef]
- Vitale, L.; Magliulo, V.; Arena, C. Morphological and physiological modifications of Cistus salvifolius L. winter leaves to rise of winter temperatures. Plant Biosyst. 2014, 148, 1093–1101. [Google Scholar] [CrossRef]
- Baas, P.; Beeckman, H.; Čufar, K.; Micco, V.D. Functional Traits in Wood Anatomy. IAWA J. 2016, 37, 124–126. [Google Scholar] [CrossRef]
- Beeckman, H. Wood anatomy and trait-based ecology. IAWA J. 2016, 37, 127–151. [Google Scholar] [CrossRef]
- Bonfante, A.; Basile, A.; Bouma, J. Exploring the Effect of Varying Soil Organic Matter Contents on Current and Future Moisture Supply Capacities of Six Italian Soils. Geoderma 2020, 361, 114079. [Google Scholar] [CrossRef]
- Terribile, F.; Gennaro, A.D.; Mascellis, R.D. Carta dei suoli della Valle Telesina (1:50,000). In Progetto UOT Relazione Finale Convenzione CNR-ISPAIM-Regione Campania Assessorato Alla Agricoltura; Raccolta di 10 Carte Pedologiche della Regione Campania; Stampa System Cart: Rome, Italy, 1996. [Google Scholar]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Forlani, M.; Basile, B. Influence of intensity of post-veraison defoliation or shoot trimming on vine physiology, yield components, berry and wine composition in Aglianico grapevines. Aust. J. Grape Wine Res. 2017, 23, 226–239. [Google Scholar] [CrossRef]
- Buttaro, D.; Rouphael, Y.; Rivera, C.M.; Colla, G.; Gonnella, M. Simple and accurate allometric model for leaf area estimation in Vitis vinifera L. genotypes. Photosynthetica 2015, 53, 342–348. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Et Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosyn. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Micco, V.D.; Arena, C.; Vitale, L.; Aronne, G.; Santo, A.V.D. Anatomy and photochemical behaviour of Mediterranean Cistus incanus winter leaves under natural outdoor and warmer indoor conditions. Botany 2011, 89, 677–688. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Shoot Leaf Area | Single Leaf Area | Shoot Basal Diameter | Average Bunch Weight | Bunch Number | |
|---|---|---|---|---|---|
| (cm2 shoot−1) | (cm2) | (mm) | (g bunch−1) | (n° shoot−1) | |
| Field (F) | |||||
| SL | 4142 ± 250 a | 192 ± 10.4 a | 8.79 ± 0.16 ab | 360 ± 11.7 a | 1.3 ± 0.04 b |
| CA | 3416 ± 171 b | 135 ± 3.24 c | 7.89 ± 0.13 c | 208 ± 9.27 c | 1.2 ± 0.03 c |
| GR | 4194 ± 228 a | 151 ± 4.22 b | 9.02 ± 0.16 a | 175 ± 10.7 d | 1.3 ± 0.03 b |
| AC | 3320 ± 156 b | 156 ± 5.21 b | 8.49 ± 0.17 b | 304 ± 11.6 b | 1.4 ± 0.05 a |
| Year (Y) | |||||
| 2019 | 4308 ± 169 a | 168 ± 5.75 a | 9.13 ± 0.10 a | 270 ± 10.8 a | 1.5 ± 0.02 a |
| 2020 | 3228 ± 107 b | 149 ± 3.42 b | 7.97 ± 0.11 b | 253 ± 11.7 a | 1.2 ± 0.02 b |
| Significance | |||||
| Field (F) | *** | *** | *** | *** | *** |
| Year (Y) | *** | ** | *** | NS | *** |
| F x Y | NS | NS | NS | *** | *** |
| Pn | gs | Ci | E | inWUE | ETR | ΦPSII | Fv/Fm | |
|---|---|---|---|---|---|---|---|---|
| (µmol CO2 m−2s−1) | (mmol H2O m−2 s−1) | (µmol CO2 mol−1) | (mol H2O m−2 s−1) | (µmol CO2/ mol H2O) | ||||
| Field (F) | ||||||||
| SL | 10.9 ± 0.66 a | 183.5 ± 19.1 a | 267.6 ± 12.5 a | 5.00 ± 0.31 a | 2.55 ± 0.26 a | 172.1 ± 5.30 a | 0.309 ± 0.007 a | 0.784 ± 0.003 a |
| CA | 6.20 ± 0.49 c | 103.6 ± 12.1 b | 228.3 ± 16.0 a | 3.37 ± 0.34 b | 1.97 ± 0.20 a | 137.3 ± 3.40 b | 0.250 ± 0.006 b | 0.752 ± 0.003 b |
| GR | 5.15 ± 0.78 c | 66.7 ± 10.9 b | 237.9 ± 17.8 a | 3.33 ± 0.37 b | 1.83 ± 0.32 a | 135.1 ± 6.19 b | 0.242 ± 0.010 b | 0.754 ± 0.004 b |
| AC | 9.31 ± 0.61 b | 159.4 ± 15.5 a | 225.2 ± 15.3 a | 5.31 ± 0.24 a | 1.98 ± 0.17 a | 181.2 ± 4.81 a | 0.325 ± 0.008 a | 0.788 ± 0.003 a |
| Year (Y) | ||||||||
| 2019 | 8.77 ± 0.58 a | 131.5 ± 14.0 a | 214.6 ± 13.6 b | 4.60 ± 0.27 a | 2.26 ± 0.21 a | 153.8 ± 5.14 a | 0.279 ± 0.008 a | 0.772 ± 0.004 a |
| 2020 | 6.78 ± 0.45 b | 120.4 ± 8.8 a | 267.4 ± 4.10 a | 4.00 ± 0.22 a | 1.74 ± 0.09 b | 159.3 ± 3.98 a | 0.284 ± 0.008 a | 0.766 ± 0.002 a |
| Significance | ||||||||
| Field (F) | *** | *** | NS | *** | NS | *** | *** | *** |
| Year (Y) | ** | NS | *** | NS | * | NS | NS | NS |
| F x Y | * | NS | NS | NS | NS | NS | NS | NS |
| Stomata Length | Stomata Width | Stomata Frequency | |
|---|---|---|---|
| (µm) | (µm) | (n/mm2) | |
| Field (F) | |||
| SL | 33.2 ± 0.43 a | 19.2 ± 0.31 a | 140.3 ± 3.74 a |
| CA | 29.6 ± 0.34 b | 16.9 ± 0.23 b | 149.2 ± 3.54 a |
| GR | 27.2 ± 0.55 c | 15.5 ± 0.36 c | 138.6 ± 4.04 a |
| AC | 24.8 ± 0.38 d | 14.3 ± 0.21 d | 139.9 ± 2.50 a |
| Year (Y) | |||
| 2019 | 28.8 ± 0.37 a | 16.1 ± 0.22 b | 151.4 ± 2.56 a |
| 2020 | 28.6 ± 0.36 a | 16.8 ± 0.24 a | 132.6 ± 1.88 b |
| Significance | |||
| Field (F) | *** | *** | NS |
| Year (Y) | NS | * | *** |
| F x Y | NS | NS | * |
| Minor VLA | Major VLA | Total VLA | Minor VAA | Major VAA | Total VAA | FVEA | |
|---|---|---|---|---|---|---|---|
| (mm/mm2) | (mm/mm2) | (mm/mm2) | (mm2/mm2) | (mm2/mm2) | (mm2/mm2) | (n/mm2) | |
| Field (F) | |||||||
| SL | 2.31 ± 0.06 a | 0.729 ± 0.036 a | 2.89 ± 0.06 c | 0.118 ± 0.004 c | 0.065 ± 0.004 a | 0.175 ± 0.005 b | 2.93 ± 0.11 a |
| CA | 2.59 ± 0.01 a | 0.865 ± 0.049 a | 3.27 ± 0.08 a | 0.132 ± 0.004 b | 0.072 ± 0.004 a | 0.194 ± 0.005 a | 2.47 ± 0.05 c |
| GR | 2.47 ± 0.09 a | 0.751 ± 0.054 a | 3.04 ± 0.07 bc | 0.134 ± 0.006 b | 0.072 ± 0.004 a | 0.197 ± 0.005 a | 2.37 ± 0.10 c |
| AC | 2.58 ± 0.14 a | 0.817 ± 0.042 a | 3.22 ± 0.12 ab | 0.145 ± 0.004 a | 0.069 ± 0.003 a | 0.194 ± 0.007 a | 2.68 ± 0.03 b |
| Year (Y) | |||||||
| 2019 | 2.45 ± 0.06 a | 0.799 ± 0.028 a | 3.07 ± 0.05 a | 0.137 ± 0.003 a | 0.075 ± 0.002 a | 0.202 ± 0.003 a | 2.48 ± 0.04 b |
| 2020 | 2.52 ± 0.09 a | 0.781 ± 0.039 a | 3.14 ± 0.07 a | 0.127 ± 0.004 b | 0.064 ± 0.003 b | 0.178 ± 0.004 b | 2.74 ± 0.06 a |
| Significance | |||||||
| Field (F) | NS | NS | * | ** | NS | ** | *** |
| Year (Y) | NS | NS | NS | * | * | *** | ** |
| F x Y | NS | NS | NS | NS | NS | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damiano, N.; Arena, C.; Bonfante, A.; Caputo, R.; Erbaggio, A.; Cirillo, C.; De Micco, V. How Leaf Vein and Stomata Traits Are Related with Photosynthetic Efficiency in Falanghina Grapevine in Different Pedoclimatic Conditions. Plants 2022, 11, 1507. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11111507
Damiano N, Arena C, Bonfante A, Caputo R, Erbaggio A, Cirillo C, De Micco V. How Leaf Vein and Stomata Traits Are Related with Photosynthetic Efficiency in Falanghina Grapevine in Different Pedoclimatic Conditions. Plants. 2022; 11(11):1507. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11111507
Chicago/Turabian StyleDamiano, Nicola, Carmen Arena, Antonello Bonfante, Rosanna Caputo, Arturo Erbaggio, Chiara Cirillo, and Veronica De Micco. 2022. "How Leaf Vein and Stomata Traits Are Related with Photosynthetic Efficiency in Falanghina Grapevine in Different Pedoclimatic Conditions" Plants 11, no. 11: 1507. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11111507