
Growth and Photosynthetic Responses of Cowpea Genotypes under Waterlogging at the Reproductive Stage
 ,
,  , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Growth Responses of Cowpea Genotypes during and after Waterlogging
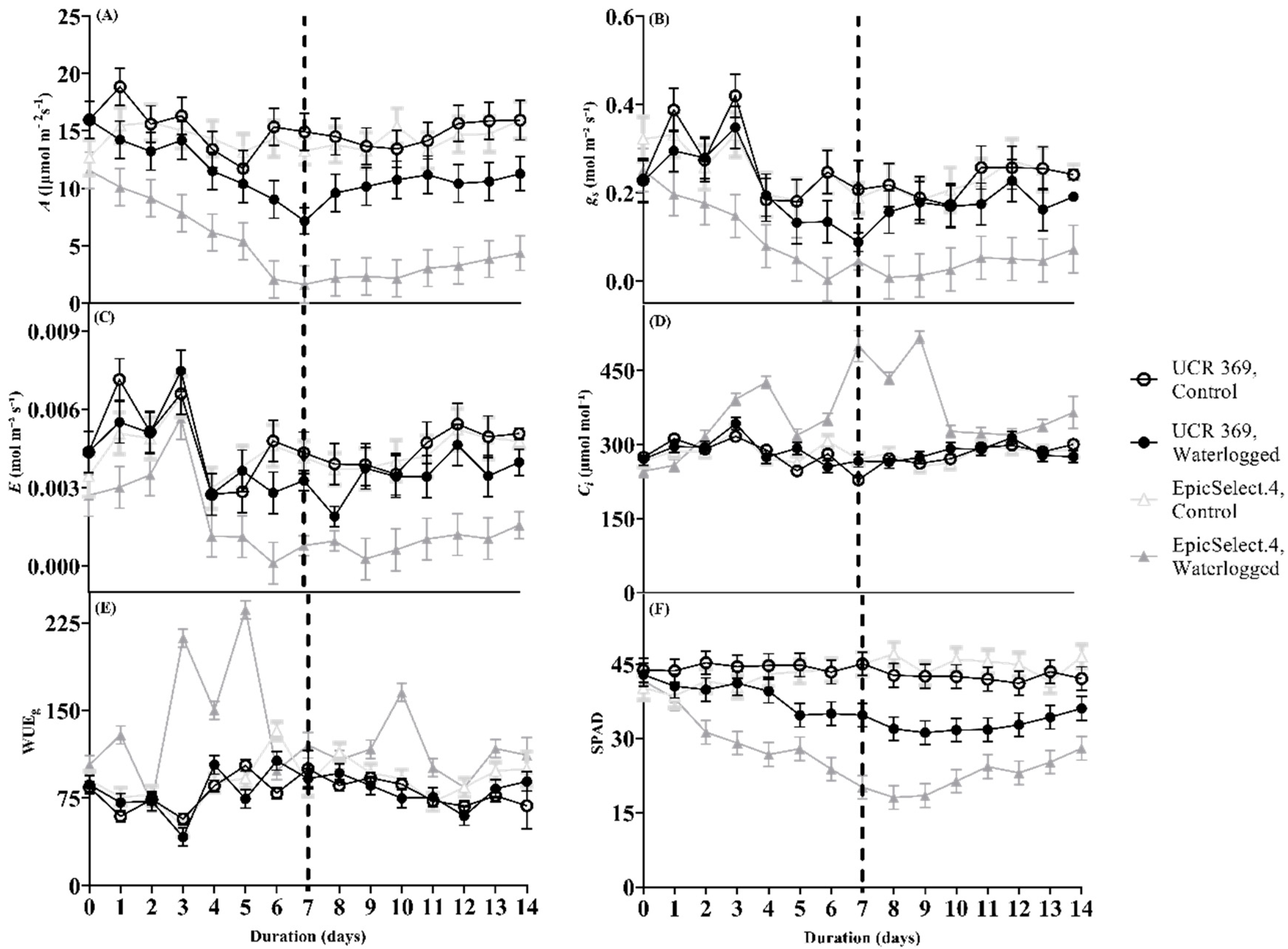
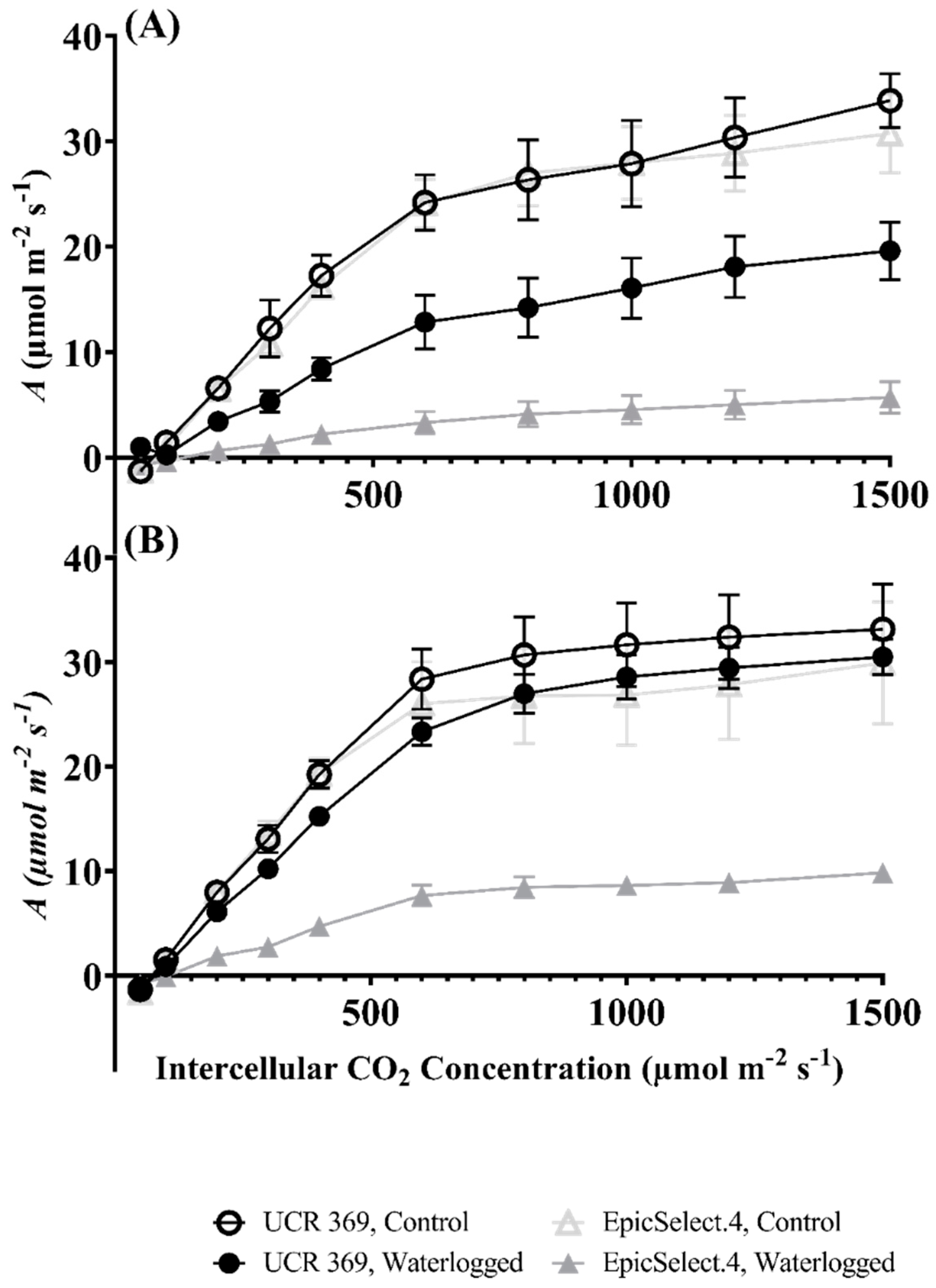
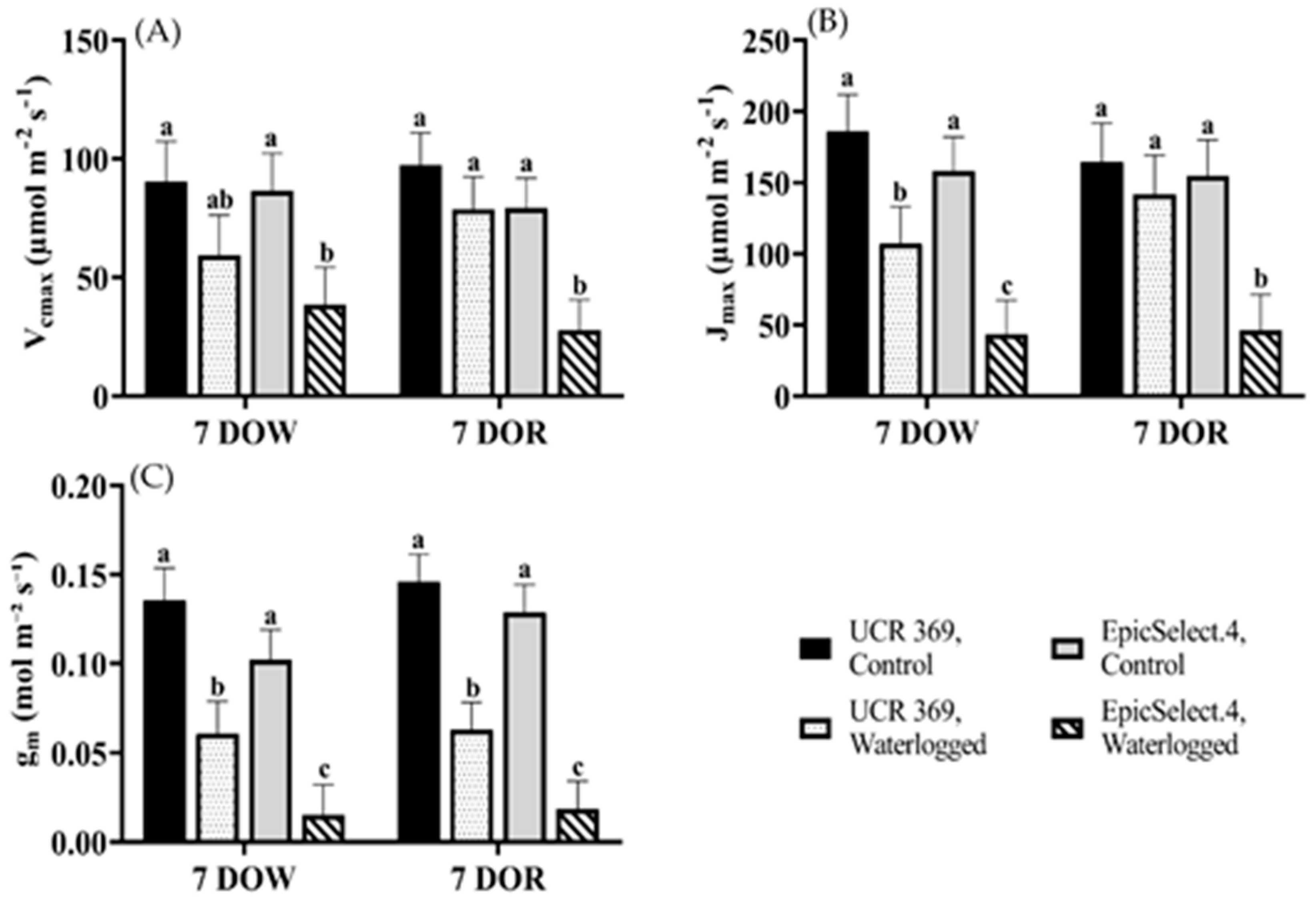
2.2. Gas Exchange Responses of Cowpea Genotypes during and after Waterlogging
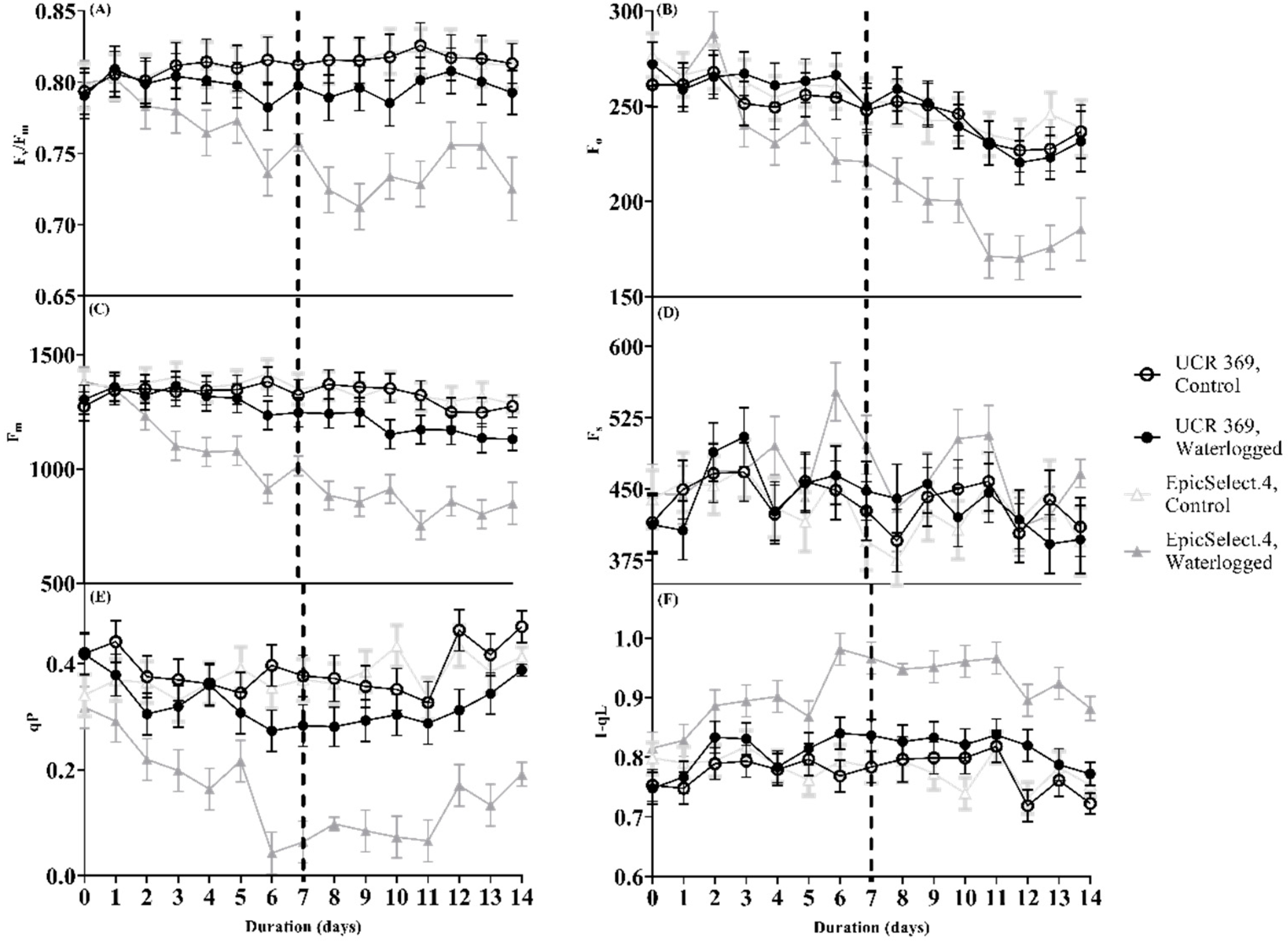
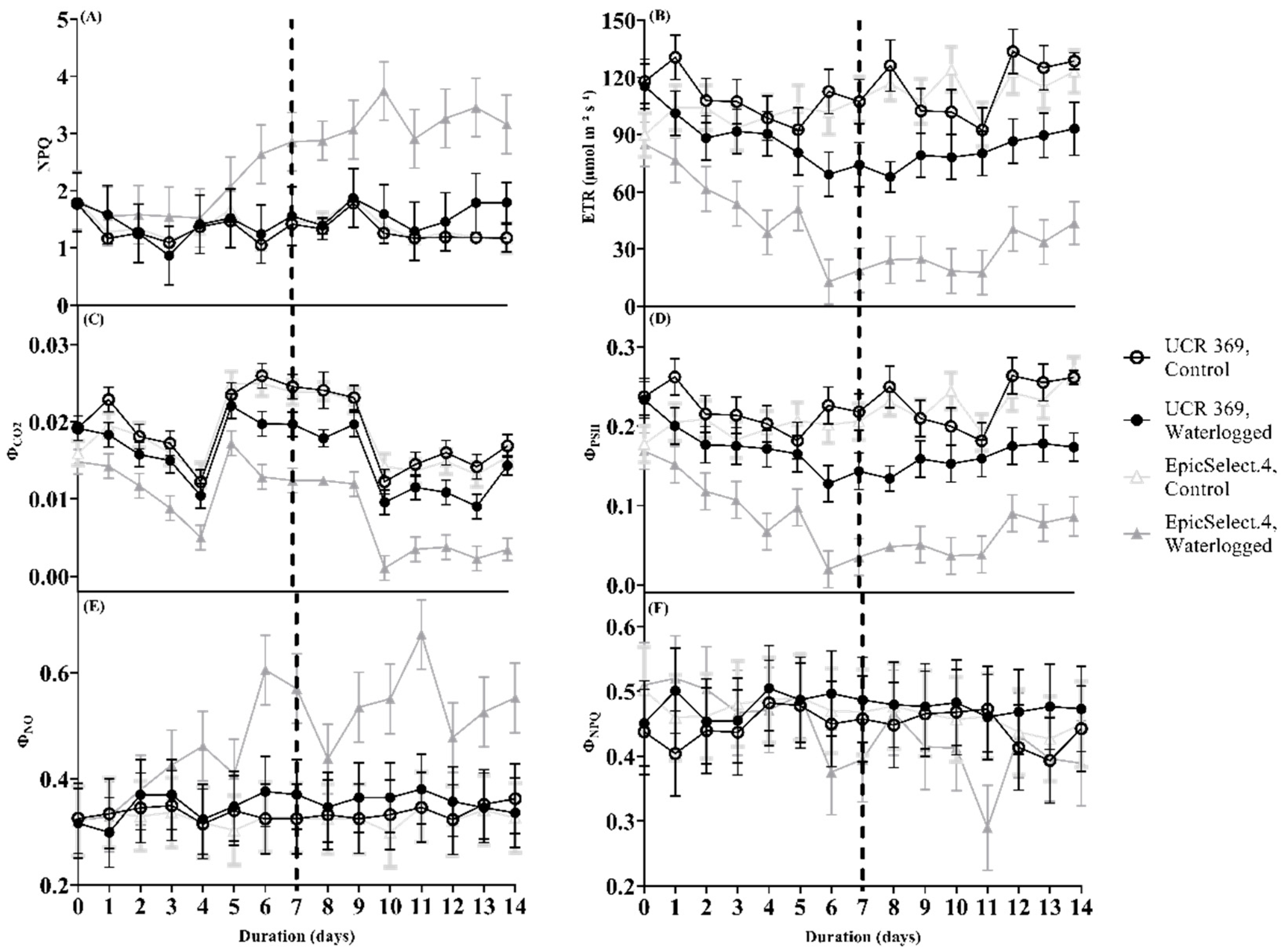
2.3. Photochemical Efficiency of Cowpea Genotypes during and after Waterlogging
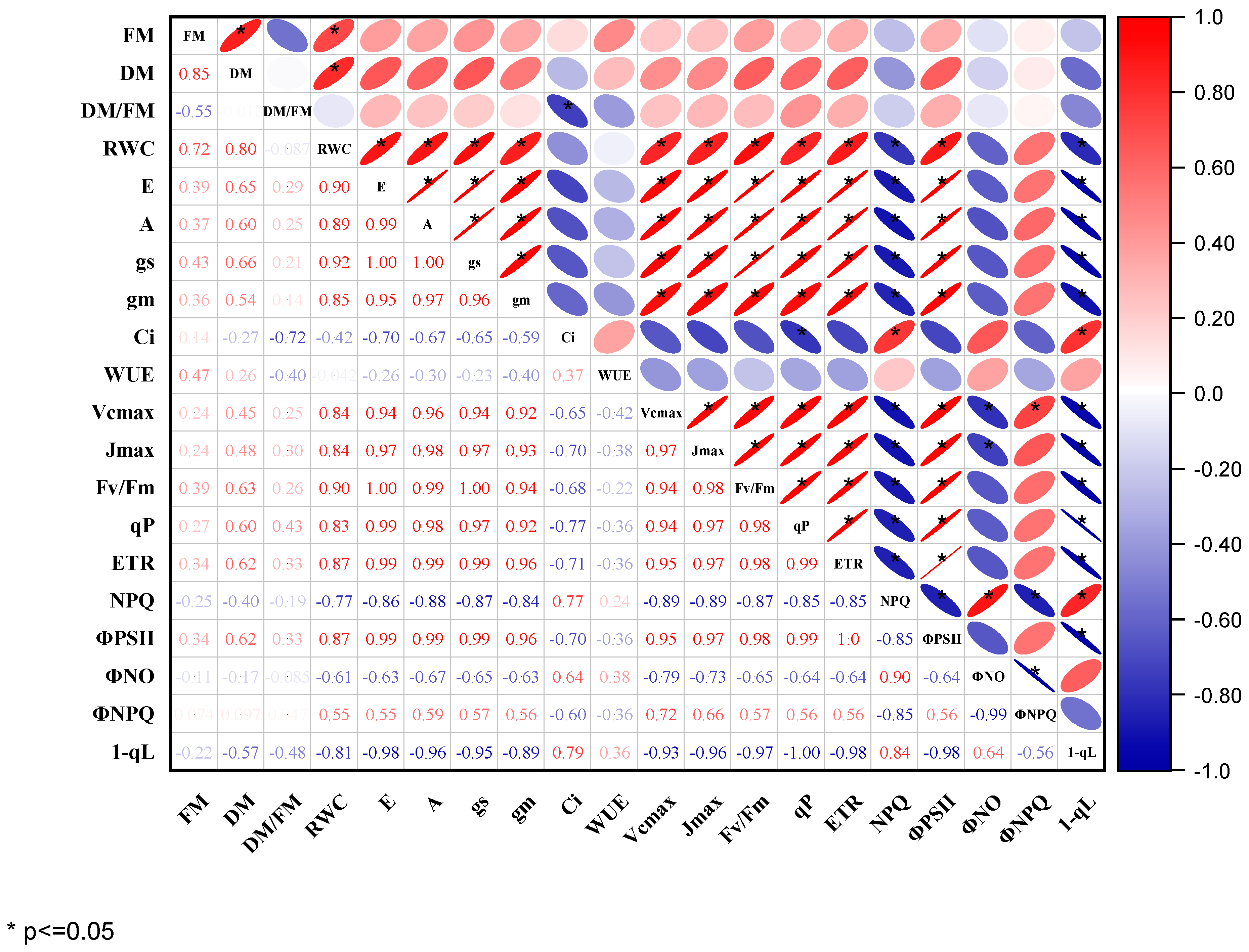
2.4. Relating Photochemical Efficiency in PSII with Leaf Gas Exchanges as Parameters for Waterlogging Tolerance
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Waterlogging Treatments
4.3. Gas Exchange Measurements
4.4. Chlorophyll Fluorescence Measurements
4.5. Plant Growth, Relative Water Content, and Harvest
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443. [Google Scholar] [CrossRef] [PubMed]
- Ploschuk, R.A.; Miralles, D.J.; Colmer, T.D.; Ploschuk, E.L.; Striker, G.G. Waterlogging of Winter Crops at Early and Late Stages: Impacts on Leaf Physiology, Growth and Yield. Front. Plant Sci. 2018, 9, 1863. [Google Scholar] [CrossRef] [PubMed]
- FAO. World Food and Agriculture—Statistical Yearbook 2021; Food Agriculture Organization of the United Nations: Rome, Italy, 2021; p. 368.
- NASA Record-Setting Precipitation Leaves U.S. Soils Soggy. Available online: https://earthobservatory.nasa.gov/images/145101/record-setting-precipitation-leaves-us-soils-soggy (accessed on 23 February 2022).
- IPCC. Summary for Policymakers. In Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC); Cambridge University Press: Cambridge, UK, 2022; p. 40. [Google Scholar]
- Easterling, D.R.; Kunkel, K.E.; Arnold, J.R.; Knutson, T.; LeGrande, A.N.; Leung, L.R.; Vose, R.S.; Waliser, D.E.; Wehner, M.F. Precipitation Change in the United States. In Climate Science Special Report: Fourth National Climate Assessment, Volume I; Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., Maycock, T.K., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2017; pp. 207–230. [Google Scholar]
- Olorunwa, O.J.; Adhikari, B.; Shi, A.; Barickman, T.C. Screening of Cowpea (Vigna unguiculata (L.) Walp.) Genotypes for Waterlogging Tolerance Using Morpho-Physiological Traits at Early Growth Stage. Plant Sci. 2022, 315, 111136. [Google Scholar] [CrossRef] [PubMed]
- Timsina, J.; Garrity, D.P.; Pandey, R.K. The Interaction of Cowpea Maturity with Degree of Waterlogging in the Post-Rice Environment. Field Crops Res. 1994, 39, 39–48. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Lee, S.C.; Brinton, E. Waterproofing Crops: Effective Flooding Survival Strategies. Plant Physiol. 2012, 160, 1698. [Google Scholar] [CrossRef]
- Ponnamperuma, F.N. The Chemistry of Submerged Soils. In Advances in Agronomy; Brady, N.C., Ed.; Academic Press: Cambridge, MA, USA, 1972; Volume 24, pp. 29–96. ISBN 0065-2113. [Google Scholar]
- Nishiuchi, S.; Yamauchi, T.; Takahashi, H.; Kotula, L.; Nakazono, M. Mechanisms for Coping with Submergence and Waterlogging in Rice. Rice 2012, 5, 2. [Google Scholar] [CrossRef]
- Kaur, G.; Zurweller, B.A.; Nelson, K.A.; Motavalli, P.P.; Dudenhoeffer, C.J. Soil Waterlogging and Nitrogen Fertilizer Management Effects on Corn and Soybean Yields. Agron. J. 2017, 109, 97–106. [Google Scholar] [CrossRef]
- Gibbs, J.; Greenway, H. Review: Mechanisms of Anoxia Tolerance in Plants. I. Growth, Survival and Anaerobic Catabolism. Funct. Plant Biol. 2003, 30, 1–47. [Google Scholar] [CrossRef]
- Araki, H.; Hamada, A.; Hossain, M.A.; Takahashi, T. Waterlogging at Jointing and/or after Anthesis in Wheat Induces Early Leaf Senescence and Impairs Grain Filling. Field Crops Res. 2012, 137, 27–36. [Google Scholar] [CrossRef]
- Arduini, I.; Baldanzi, M.; Pampana, S. Reduced Growth and Nitrogen Uptake During Waterlogging at Tillering Permanently Affect Yield Components in Late Sown Oats. Front. Plant Sci. 2019, 10, 1087. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Al Mahmud, J.; Nahar, K.; Anee, T.I.; Inafuku, M.; Oku, H.; Fujita, M. Responses, Adaptation, and ROS Metabolism in Plants Exposed to Waterlogging Stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 257–281. ISBN 978-981-10-5254-5. [Google Scholar]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of Waterlogging Tolerance in Plants: Research Progress and Prospects. Front. Plant Sci. 2021, 11, 2319. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Pal, M.; Joshi, R.; Sairam, R.K. Yield, Growth and Physiological Responses of Mung Bean [Vigna radiata (L.) Wilczek] Genotypes to Waterlogging at Vegetative Stage. Physiol. Mol. Biol. Plants 2013, 19, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; da-Silva, C.J.; Cocco, K.L.T.; Pomagualli, D.; de Oliveira, F.K.; da Silva, J.V.L.; de Oliveira, A.C.B.; do Amarante, L. Waterlogging Tolerance of Five Soybean Genotypes through Different Physiological and Biochemical Mechanisms. Environ. Exp. Bot. 2020, 172, 103975. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flood Adaptive Traits and Processes: An Overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Kreuzwieser, J.; Rennenberg, H. Molecular and Physiological Responses of Trees to Waterlogging Stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef]
- Black, K.; Davis, P.; Mc Grath, J.; Doherty, P.; Osborne, B. Interactive Effects of Irradiance and Water Availability on the Photosynthetic Performance of Picea Sitchensis Seedlings: Implications for Seedling Establishment under Different Management Practices. Ann. For. Sci. 2005, 62, 413–422. [Google Scholar] [CrossRef]
- Zhou, S.; Medlyn, B.; Sabaté, S.; Sperlich, D.; Prentice, I.C.; Whitehead, D. Short-Term Water Stress Impacts on Stomatal, Mesophyll and Biochemical Limitations to Photosynthesis Differ Consistently among Tree Species from Contrasting Climates. Tree Physiol. 2014, 34, 1035–1046. [Google Scholar] [CrossRef]
- Pompeiano, A.; Huarancca Reyes, T.; Moles, T.M.; Guglielminetti, L.; Scartazza, A. Photosynthetic and Growth Responses of Arundo donax L. Plantlets Under Different Oxygen Deficiency Stresses and Reoxygenation. Front. Plant Sci. 2019, 10, 408. [Google Scholar] [CrossRef]
- Bansal, R.; Srivastava, J.P. Effect of Waterlogging on Photosynthetic and Biochemical Parameters in Pigeonpea. Russ. J. Plant Physiol. 2015, 62, 322–327. [Google Scholar] [CrossRef]
- Olorunwa, O.J.; Adhikari, B.; Brazel, S.; Popescu, S.; Popescu, G.; Barickman, T.C. Short Waterlogging Events Differently Affect Morphology and Photosynthesis of Two Cucumber (Cucumis sativus L.) Cultivars. Front. Plant Sci. 2022, 13, 2530. [Google Scholar] [CrossRef]
- Sairam, R.K.; Kumutha, D.; Ezhilmathi, K.; Chinnusamy, V.; Meena, R.C. Waterlogging Induced Oxidative Stress and Antioxidant Enzyme Activities in Pigeon Pea. Biol. Plant. 2009, 53, 493–504. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll Fluorescence, Photoinhibition and Abiotic Stress: Does It Make Any Difference the Fact to Be a C3 or C4 Species? Front.Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Smethurst, C.F.; Shabala, S. Screening Methods for Waterlogging Tolerance in Lucerne: Comparative Analysis of Waterlogging Effects on Chlorophyll Fluorescence, Photosynthesis, Biomass and Chlorophyll Content. Funct. Plant Biol. 2003, 30, 335. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in Photosynthesis and Some Antioxidant Enzymatic Activities of Mungbean Subjected to Waterlogging. Plant Sci. 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Velasco, N.F.; Ligarreto, G.A.; Díaz, H.R.; Fonseca, L.P.M. Photosynthetic Responses and Tolerance to Root-Zone Hypoxia Stress of Five Bean Cultivars (Phaseolus vulgaris L.). S. Afr. J. Bot. 2019, 123, 200–207. [Google Scholar] [CrossRef]
- Minchin, F.R.; Summerfield, R.J.; Eaglesham, A.R.J.; Stewart, K.A. Effects of Short-Term Waterlogging on Growth and Yield of Cowpea (Vigna unguiculata). J. Agric. Sci. 1978, 90, 355–366. [Google Scholar] [CrossRef]
- Umaharan, P.; Ariyanayagam, R.P.; HAQUE, S.Q. Effect of Short-Term Waterlogging Applied at Various Growth Phases on Growth, Development and Yield in Vigna Unguiculata. J. Agric. Sci. 1997, 128, 189–198. [Google Scholar] [CrossRef]
- Ploschuk, R.A.; Miralles, D.J.; Striker, G.G. A Quantitative Review of Soybean Responses to Waterlogging: Agronomical, Morpho-Physiological and Anatomical Traits of Tolerance. Plant Soil 2022, 475, 237–252. [Google Scholar] [CrossRef]
- Hong, T.D.; Minchin, F.R.; Summerfield, R.J. Recovery of Nodulated Cowpea Plants (Vigna unguiculata (L.) Walp.) From Waterlogging During Vegetative Growth. Plant Soil 1977, 48, 661–672. [Google Scholar] [CrossRef]
- Barik, J.; Panda, D.; Mohanty, S.K.; Lenka, S.K. Leaf Photosynthesis and Antioxidant Response in Selected Traditional Rice Landraces of Jeypore Tract of Odisha, India to Submergence. Physiol. Mol. Biol. Plants 2019, 25, 847–863. [Google Scholar] [CrossRef]
- Peng, J.; Feng, Y.; Wang, X.; Li, J.; Xu, G.; Phonenasay, S.; Luo, Q.; Han, Z.; Lu, W. Effects of Nitrogen Application Rate on the Photosynthetic Pigment, Leaf Fluorescence Characteristics, and Yield of Indica Hybrid Rice and Their Interrelations. Sci. Rep. 2021, 11, 7485. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Murchie, E.H. Assessing the Photoprotective Effectiveness of Non-Photochemical Chlorophyll Fluorescence Quenching: A New Approach. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QA Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and Waterlogging Stress in Plants: A Review Highlighting Research Opportunities and Understudied Aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Kyu, K.L.; Malik, A.I.; Colmer, T.D.; Siddique, K.H.M.; Erskine, W. Response of Mungbean (Cvs. Celera II-AU and Jade-AU) and Blackgram (Cv. Onyx-AU) to Transient Waterlogging. Front. Plant Sci. 2021, 12, 709102. [Google Scholar] [CrossRef]
- Bacanamwo, M.; Purcell, L.C. Soybean Dry Matter and N Accumulation Responses to Flooding Stress, N Sources and Hypoxia. J. Exp. Bot. 1999, 50, 689–696. [Google Scholar] [CrossRef]
- Soltani, A.; MafiMoghaddam, S.; Oladzad-Abbasabadi, A.; Walter, K.; Kearns, P.J.; Vasquez-Guzman, J.; Mamidi, S.; Lee, R.; Shade, A.L.; Jacobs, J.L.; et al. Genetic Analysis of Flooding Tolerance in an Andean Diversity Panel of Dry Bean (Phaseolus vulgaris L.). Front. Plant Sci. 2018, 9, 767. [Google Scholar] [CrossRef]
- Malik, A.I.; Ailewe, T.I.; Erskine, W. Tolerance of Three Grain Legume Species to Transient Waterlogging. AoB PLANTS 2015, 7, 1–11. [Google Scholar] [CrossRef]
- Vitor, S.C.; Sodek, L. Products of Anaerobic Metabolism in Waterlogged Roots of Soybean Are Exported in the Xylem. Plant Sci. 2019, 284, 82–90. [Google Scholar] [CrossRef]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R.C. Waterlogging Induced Oxidative Stress and Antioxidant Activity in Pigeonpea Genotypes. Biol. Plant. 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Kramer, P.J.; Jackson, W.T. Causes of Injury to Flooded Tobacco Plants. Plant Physiol. 1954, 29, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Brazel, S.R.; Barickman, T.C.; Sams, C.E. Short-Term Waterlogging of Kale (Brassica oleracea L. Var. Acephala) Plants Causes a Decrease in Carotenoids and Chlorophylls While Increasing Nutritionally Important Glucosinolates. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS), Leuven, Belgium, 3 December 2021; pp. 175–180. [Google Scholar]
- Else, M.A.; Janowiak, F.; Atkinson, C.J.; Jackson, M.B. Root Signals and Stomatal Closure in Relation to Photosynthesis, Chlorophyll a Fluorescence and Adventitious Rooting of Flooded Tomato Plants. Ann. Bot. 2009, 103, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Chen, L.; Wang, X.; Cao, J.; Li, X.; Xu, X.; Xia, Q.; Chen, T.; Zhang, L. Effect of Waterlogging Stress on Dry Matter Accumulation, Photosynthesis Characteristics, Yield, and Yield Components in Three Different Ecotypes of Peanut (Arachis hypogaea L.). Agronomy 2020, 10, 1244. [Google Scholar] [CrossRef]
- Islam, M.R.; Hamid, A.; Karim, M.A.; Haque, M.M.; Khaliq, Q.A.; Ahmed, J.U. Gas Exchanges and Yield Responses of Mungbean (Vigna radiata L. Wilczek) Genotypes Differing in Flooding Tolerance. Acta Physiol. Plant. 2008, 30, 697–707. [Google Scholar] [CrossRef]
- Meena, K.C.; Rao, S.; Gontia, A.S.; Upadhyaya, A.; Rao, S.K. Physiological Performance of Pigeon Pea [Cajanus cajan (L.) Millsp.] Genotypes after Recovery from Waterlogging. Ind. J. Plant Physiol. 2015, 20, 162–166. [Google Scholar] [CrossRef]
- Rao, L.; Li, S.; Cui, X. Leaf Morphology and Chlorophyll Fluorescence Characteristics of Mulberry Seedlings under Waterlogging Stress. Sci. Rep. 2021, 11, 13379. [Google Scholar] [CrossRef]
- Zhu, M.; Li, F.H.; Shi, Z.S. Morphological and Photosynthetic Response of Waxy Corn Inbred Line to Waterlogging. Photosynthetica 2016, 54, 636–640. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence and Photosynthesis: The Basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII Functionality of Drought-Resistant and Drought-Sensitive Weeping Lovegrass Plants. Environ.Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- Li, G.; Wan, S.; Zhou, J.; Yang, Z.; Qin, P. Leaf Chlorophyll Fluorescence, Hyperspectral Reflectance, Pigments Content, Malondialdehyde and Proline Accumulation Responses of Castor Bean (Ricinus communis L.) Seedlings to Salt Stress Levels. Ind. Crops Prod. 2010, 31, 13–19. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, J.; Wang, Y.; Yang, Q.; Chen, T.; Chen, Y.; Chi, D.; Xia, G.; Siddique, K.H.M.; Wang, T. Photosynthesis, Chlorophyll Fluorescence, and Yield of Peanut in Response to Biochar Application. Front. Plant Sci. 2021, 12, 650432. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Li, M.-S.; Chen, K.-T.; Lin, Y.-H.; Ko, S.-S. Photosynthetic and Morphological Responses of Sacha Inchi (Plukenetia volubilis L.) to Waterlogging Stress. Plants 2022, 11, 249. [Google Scholar] [CrossRef]
- Zhang, R.D.; Zhou, Y.F.; Yue, Z.X.; Chen, X.F.; Cao, X.; Xu, X.X.; Xing, Y.F.; Jiang, B.; Ai, X.Y.; Huang, R.D. Changes in Photosynthesis, Chloroplast Ultrastructure, and Antioxidant Metabolism in Leaves of Sorghum under Waterlogging Stress. Photosynthetica 2019, 57, 1076–1083. [Google Scholar] [CrossRef]
- Ma, Y.-Z.; Holt, N.E.; Li, X.-P.; Niyogi, K.K.; Fleming, G.R. Evidence for Direct Carotenoid Involvement in the Regulation of Photosynthetic Light Harvesting. Proc. Natl. Acad. Sci. USA 2003, 100, 4377–4382. [Google Scholar] [CrossRef]
- Martin, B.; Ruiz-Torres, N.A. Effects of Water-Deficit Stress on Photosynthesis, Its Components and Component Limitations, and on Water Use Efficiency in Wheat (Triticum aestivum L.). Plant Physiol. 1992, 100, 733. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting Photosynthetic Carbon Dioxide Response Curves for C3 Leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef]
- Olorunwa, O.J.; Shi, A.; Barickman, T.C. Varying Drought Stress Induces Morpho-Physiological Changes in Cowpea (Vigna unguiculata (L.) Genotypes Inoculated with Bradyrhizobium Japonicum. Plant Stress 2021, 2, 100033. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Singsaas, E.L.; Pimentel, C.; Portis Jr, A.R.; Long, S.P. Improved Temperature Response Functions for Models of Rubisco-Limited Photosynthesis. Plant Cell Environ. 2001, 24, 253–259. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta (BBA) Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Barrs, H.; Weatherley, P. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FM (g/plant) | DM (g/plant) | DM/FM (%) | RWC (%) | ||
|---|---|---|---|---|---|
| UCR 369 | |||||
| HARVEST 1 (3 DOW) | Control | 92.68 ± 4.63 b | 16.59 ± 1.16 h | 17.79 ± 0.56 b | 83.30 ± 1.13 f |
| Waterlogging | 89.60 ± 5.42 b | 17.93 ± 0.92 gh | 20.50 ± 0.36 a | 84.27 ± 3.18 ef | |
| EpicSelect.4 | |||||
| Control | 148.45 ± 6.33 a | 24.07 ± 1.22 bc | 16.20 ± 0.39 c | 96.19 ± 0.60 a | |
| Waterlogging | 137.66 ± 7.31 a | 21.84 ± 1.24 cde | 15.86 ± 0.24 c | 80.09 ± 1.60 g | |
| HARVEST 2 (7 DOW) | UCR 369 | ||||
| Control | 119.22 ± 5.17 b | 21.28 ± 0.81 def | 18.07 ± 0.72 b | 86.55 ± 0.46 cd | |
| Waterlogging | 120.02 ± 4.21 b | 24.43 ± 0.55 bc | 20.22 ± 0.35 a | 77.91 ± 0.84 gh | |
| EpicSelect.4 | |||||
| Control | 170.07 ± 5.90 a | 24.63 ± 1.19 b | 14.40 ± 0.34 c | 93.83 ± 1.33 b | |
| Waterlogging | 107.60 ± 10.60 c | 13.42 ± 1.55 i | 12.10 ± 0.42 d | 68.95 ± 1.76 i | |
| HARVEST 3 (3 DOR) | UCR 369 | ||||
| Control | 135.42 ± 8.10 b | 22.43± 1.30 bcd | 16.33 ± 0.33 b | 87.62 ± 0.92 cd | |
| Waterlogging | 98.82 ± 3.76 c | 19.40 ± 0.58 fg | 19.76 ± 0.32 a | 78.29 ± 0.99 g | |
| EpicSelect.4 | |||||
| Control | 176.74 ± 8.27 a | 22.94 ± 1.10 bcd | 12.98 ± 0.09 c | 96.06 ± 0.80 a | |
| Waterlogging | 76.94 ± 2.53 d | 10.12 ± 0.44 j | 13.16 ± 0.42 c | 79.40 ± 0.35 g | |
| HARVEST 4 (7 DOR) | UCR 369 | ||||
| Control | 119.39 ± 7.10 b | 21.85 ± 1.14 cde | 18.43 ± 0.31 b | 85.96 ± 1.04 cde | |
| Waterlogging | 93.65 ± 3.04 c | 19.75 ± 0.64 ef | 21.12 ± 0.32 a | 81.57 ± 1.05 fg | |
| EpicSelect.4 | |||||
| Control | 182.83 ± 3.22 a | 27.79 ± 0.64 a | 15.21 ± 0.28 d | 96.47 ± 0.57 a | |
| Waterlogging | 115.82 ± 5.58 b | 19.86 ± 0.69 efg | 17.39 ± 0.40 c | 76.09 ± 0.73 h | |
| Treatment | *** | ** | *** | *** | |
| Genotype | *** | NS | *** | NS | |
| Harvest | *** | *** | *** | *** | |
| Treatment * Genotype | *** | ** | NS | *** | |
| Treatment * Harvest | *** | *** | NS | * | |
| Genotype * Harvest | *** | *** | *** | *** | |
| Treatment * Genotype * Harvest | NS | * | NS | *** | |
| Studied Parameters | Source of Variation | ||||||
|---|---|---|---|---|---|---|---|
| Genotype (G) | Treatment (T) | Duration (D) | G × T | G × D | T × D | G × T × D | |
| CCI (SPAD) | 111.76 *** | 878.91 *** | 10.36 *** | 108.32 *** | 1.82 * | 14.56 *** | 3.07 *** |
| A (μmol m−2 s−1) | 110.44 *** | 335.44 *** | 9.06 *** | 72.78 *** | 0.67 NS | 4.53 *** | 1.39 NS |
| gs (mol m−2 s−1) | 51.08 *** | 66.43 *** | 14.06 *** | 34.51 *** | 0.59 NS | 3.04 ** | 0.72 NS |
| E(mol m−2 s−1) | 66.64 *** | 94.43 *** | 17.9 *** | 45.59 *** | 0.72 NS | 3.22 *** | 0.85 NS |
| Ci(μmol m−1 ) | 7.40 ** | 11.83 ** | 2.87 ** | 8.49 ** | 2.22 * | 2.11 * | 2.26 * |
| WUE | 16.13 *** | 8.00 ** | 1.32 NS | 8.21 ** | 0.89 NS | 0.88NS | 1.69 NS |
| Fv/Fm | 40.14 *** | 120.79 *** | 1.43 NS | 40.34 *** | 1.48 NS | 3.85 *** | 1.34 NS |
| Fm | 69.74 *** | 273.48 *** | 13.44 *** | 111.26 *** | 3.44 *** | 8.19 *** | 2.20 * |
| Fo | 28.44 *** | 43.42 *** | 26.82 *** | 58.71 *** | 2.97 *** | 3.58 *** | 2.44 ** |
| Fs | 4.06 * | 3.85 * | 4.03 *** | 5.54 * | 1.3 NS | 1.32NS | 1.07 NS |
| F′m | 6.99 ** | 36.04 *** | 3.24 *** | 0.64 NS | 1.09 NS | 2.16 * | 0.76 NS |
| F′o | 21.29 *** | 19.4 *** | 4.65 *** | 13.08 *** | 1.68 NS | 1.17 NS | 0.90 NS |
| F′v/F′m | 0.56 NS | 37.77 *** | 3.92 *** | 0.42NS | 1.7 NS | 2.18 * | 0.97 NS |
| ΦPSII | 108.87 *** | 316.93 *** | 6.19 *** | 68.53 *** | 0.78 NS | 5.49 *** | 2.23 * |
| ΦCO2 | 109.72 *** | 332.55 *** | 68.34 *** | 71.87 *** | 0.66 NS | 4.45 *** | 1.38NS |
| ETR | 108.87 *** | 316.93 *** | 6.19 *** | 68.53 *** | 0.78 NS | 5.49 *** | 2.23 * |
| NPQ | 5.33 * | 7.33 ** | 1.45 NS | 4.36 * | 0.8 NS | 1.06 NS | 0.82 NS |
| qP | 114.35 *** | 285.72 *** | 7.28 *** | 84.05 *** | 1.19 NS | 5.15 *** | 2.65 ** |
| qN | 2.34 NS | 31.39 *** | 16.08 *** | 0.06 | 1.38 NS | 2.33 ** | 0.95 NS |
| 1-qL | 86.12 *** | 205.19 *** | 7.05 *** | 69.8 *** | 1.16 NS | 4.34 *** | 2.09 * |
| ΦNO | 7.41 ** | 9.36 ** | 1.01 NS | 7.87 ** | 0.83 NS | 1.00 NS | 0.84 NS |
| ΦNPQ | 5.52 * | 5.91 * | 0.94 NS | 6.3 * | 0.82 NS | 0.88 NS | 0.77 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olorunwa, O.J.; Adhikari, B.; Brazel, S.; Shi, A.; Popescu, S.C.; Popescu, G.V.; Barickman, T.C. Growth and Photosynthetic Responses of Cowpea Genotypes under Waterlogging at the Reproductive Stage. Plants 2022, 11, 2315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172315
Olorunwa OJ, Adhikari B, Brazel S, Shi A, Popescu SC, Popescu GV, Barickman TC. Growth and Photosynthetic Responses of Cowpea Genotypes under Waterlogging at the Reproductive Stage. Plants. 2022; 11(17):2315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172315
Chicago/Turabian StyleOlorunwa, Omolayo J., Bikash Adhikari, Skyler Brazel, Ainong Shi, Sorina C. Popescu, George V. Popescu, and T. Casey Barickman. 2022. "Growth and Photosynthetic Responses of Cowpea Genotypes under Waterlogging at the Reproductive Stage" Plants 11, no. 17: 2315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172315