
Fertilizers and Fertilization Strategies Mitigating Soil Factors Constraining Efficiency of Nitrogen in Plant Production


Abstract
:1. Fertilizer Use Efficiency—A Real Farming Practice
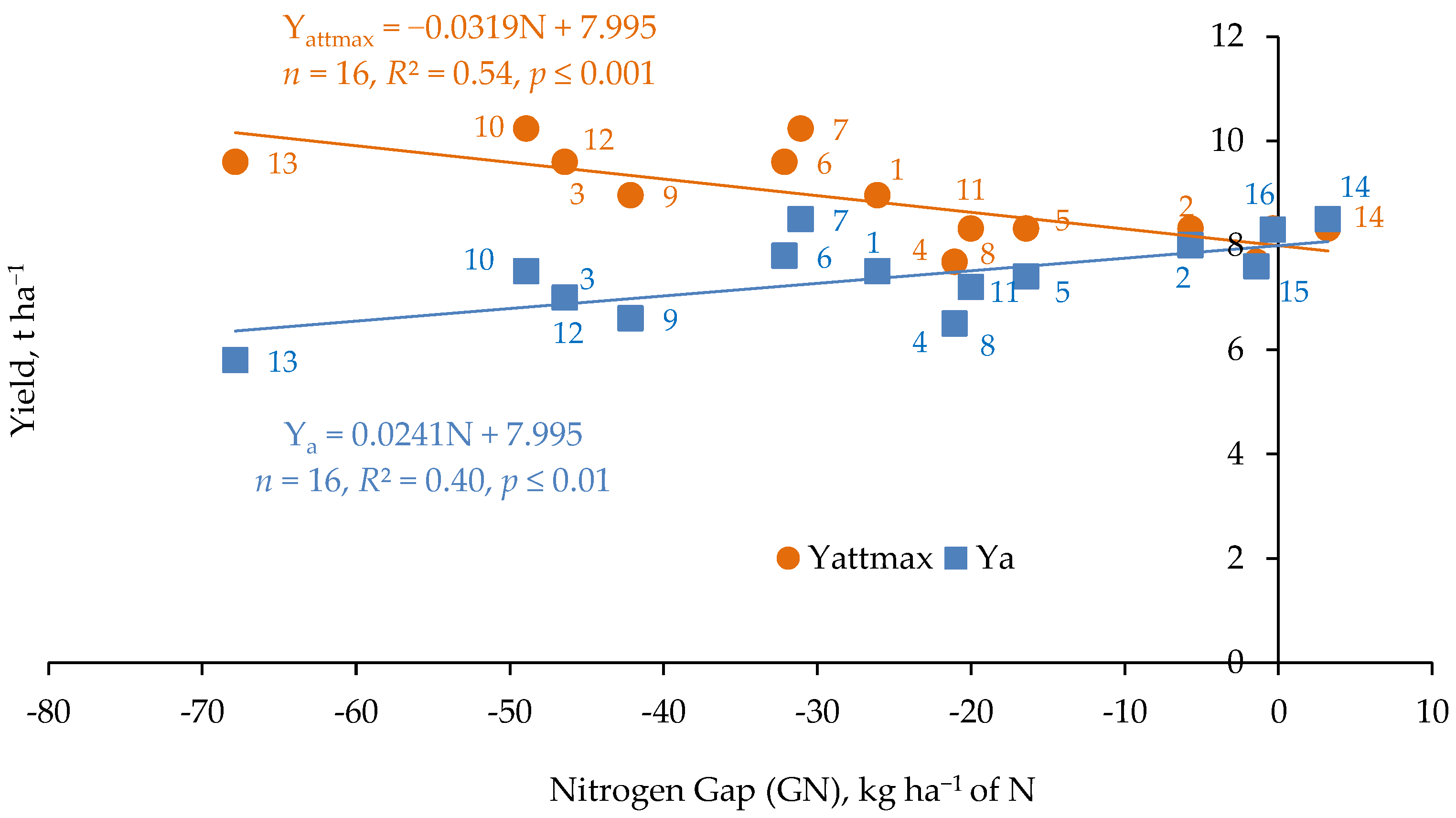
1.1. Nitrogen Gap and the Maximum Attainable Yield
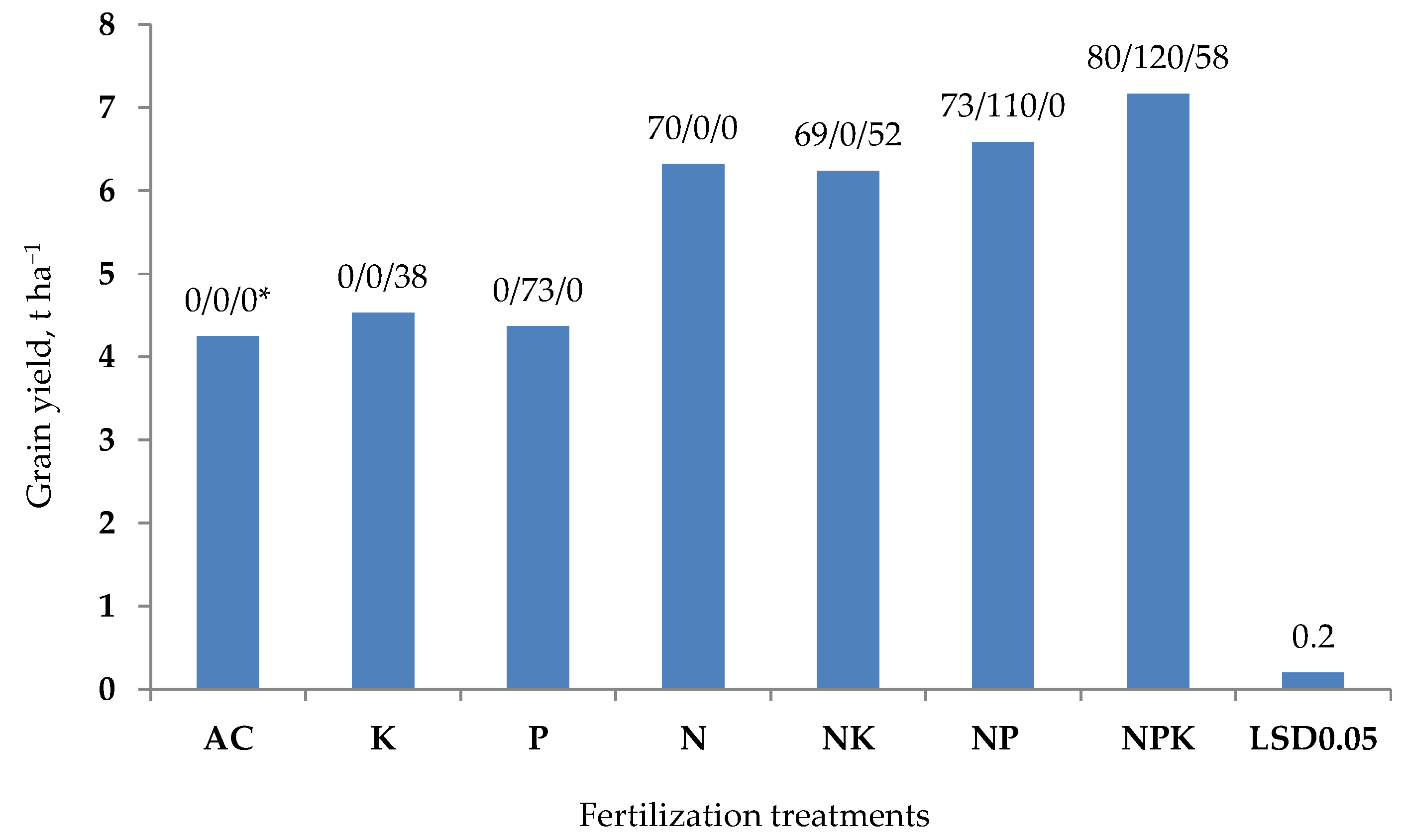
1.2. Fertilizer Use Efficiency—FUE
1.3. Factors Affecting Fertilizer Use Efficiency
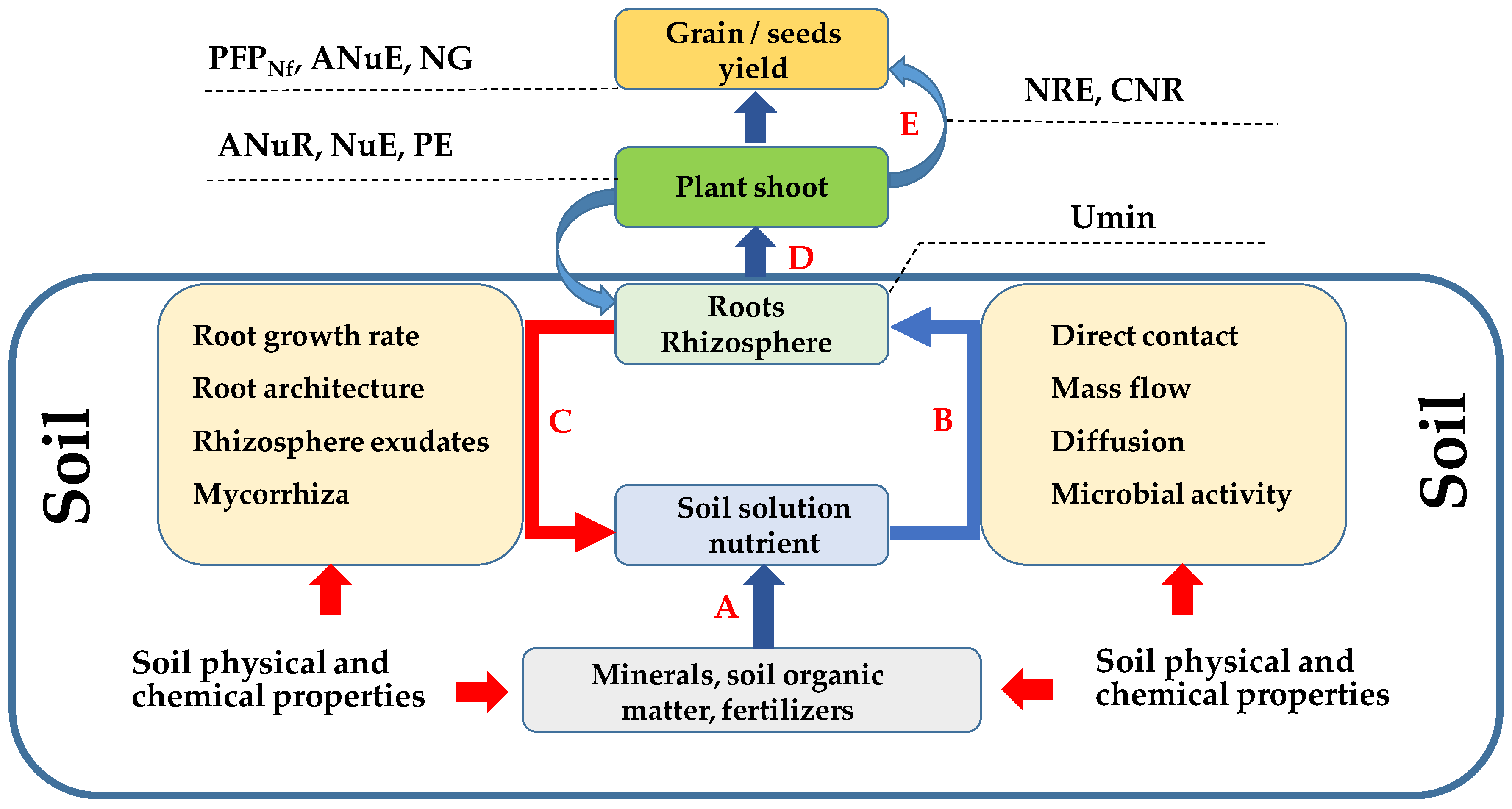
2. Factors Affecting Nutrient Uptake
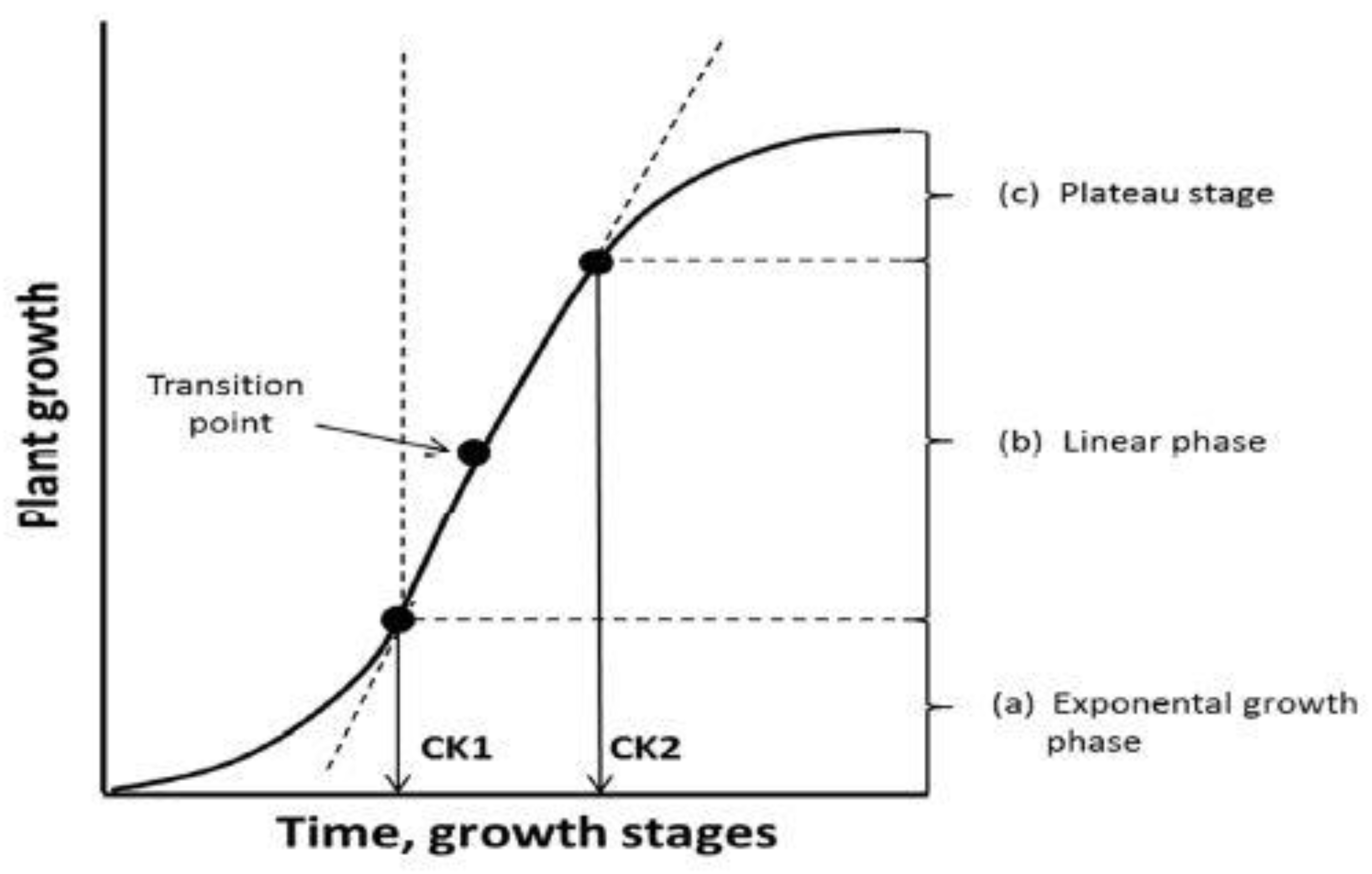
2.1. Plant Growth and Nutrient Requirement
- Exponential → Crop Foundation Period—CFP;
- Linear → Yield Formation Period—YFP;
- Quadratic or linear plateau → Yield Realization Period—YRP.
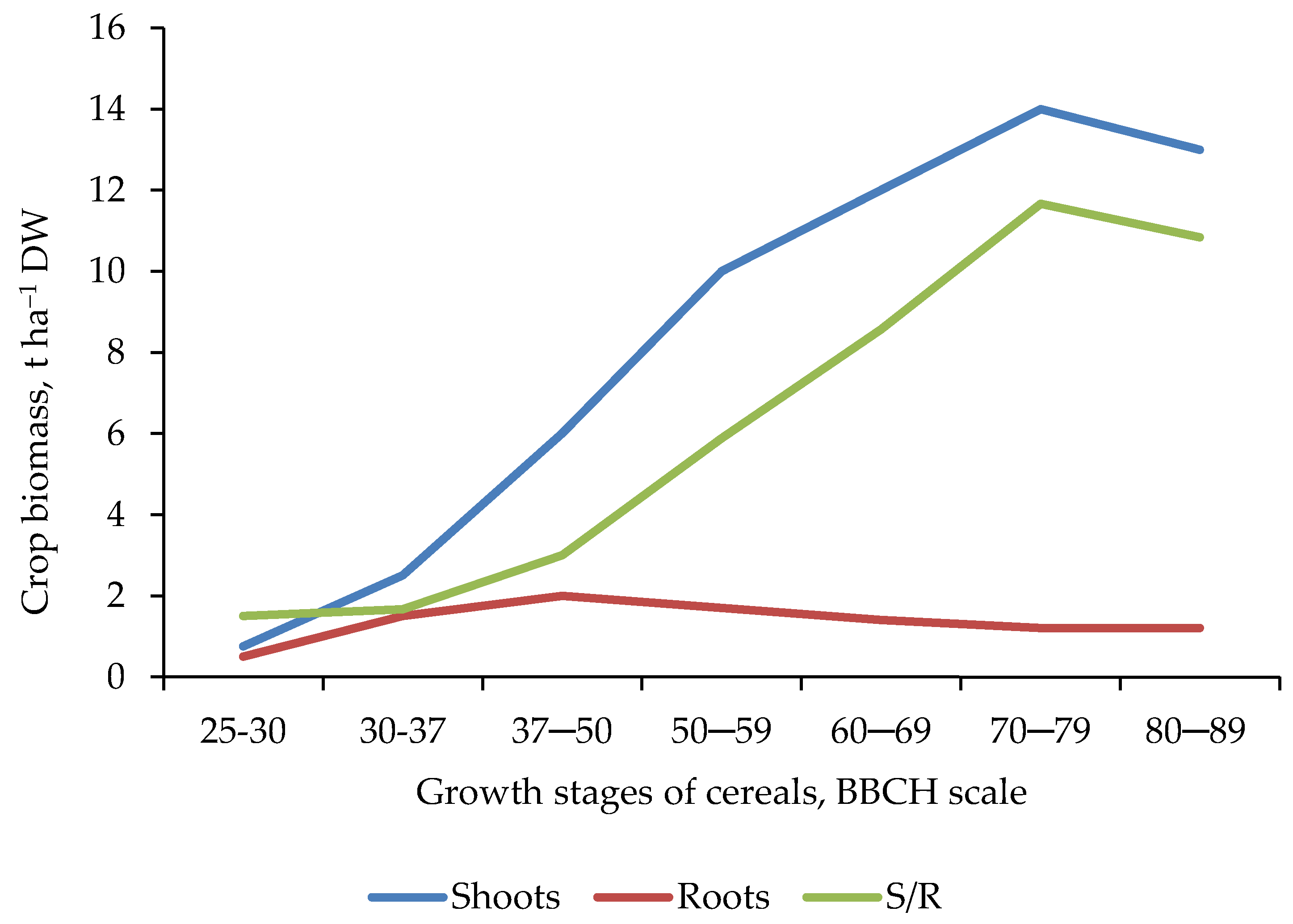
2.2. The Root System Architecture—RSA
- Movement of nutrients along the soil/plant continuum:
- transport of ions/molecules from the soil solution towards the root surface,
- ingrowth of the root into soil patches rich in available nutrients;
- Transport of nutrients adsorbed on the root surface through the plasma membrane into the cytoplasm;
- Direct utilization of the nutrient in the root or its transport via the xylem to active plant tissues.
- Anchorage of the plant in the soil;
- Water extraction from the soil to:
- stabilize the shoot temperature
- transport nutrients to the shoot;
- Nutrient uptake from the soil solution;
- Impact on rhizosphere processes through:
- release of organic compounds → a source of energy for microorganisms present in the rhizosphere
- release of protons or chelating agents → increase in nutrient availability
- deposition of carbon by dead roots → humus build-up;
- Symbiotic associations with bacteria or fungi;
- Storage organs, treated as main yield (sugar beets, cassava, sweet potato).
- Primary root (PR) length, which determines the depth of a plant rooting;
- Root branching patterns, which are represented by a number of characteristics, among others (i) number of lateral roots (LR), number of adventitious roots (AR), (ii) growth angle of LR and AR in relation to the primary root (s), (iii) root diameter, (iv) root length density (RLD);
- Root hairs (RH), including length, diameter, number per root unit length or area.
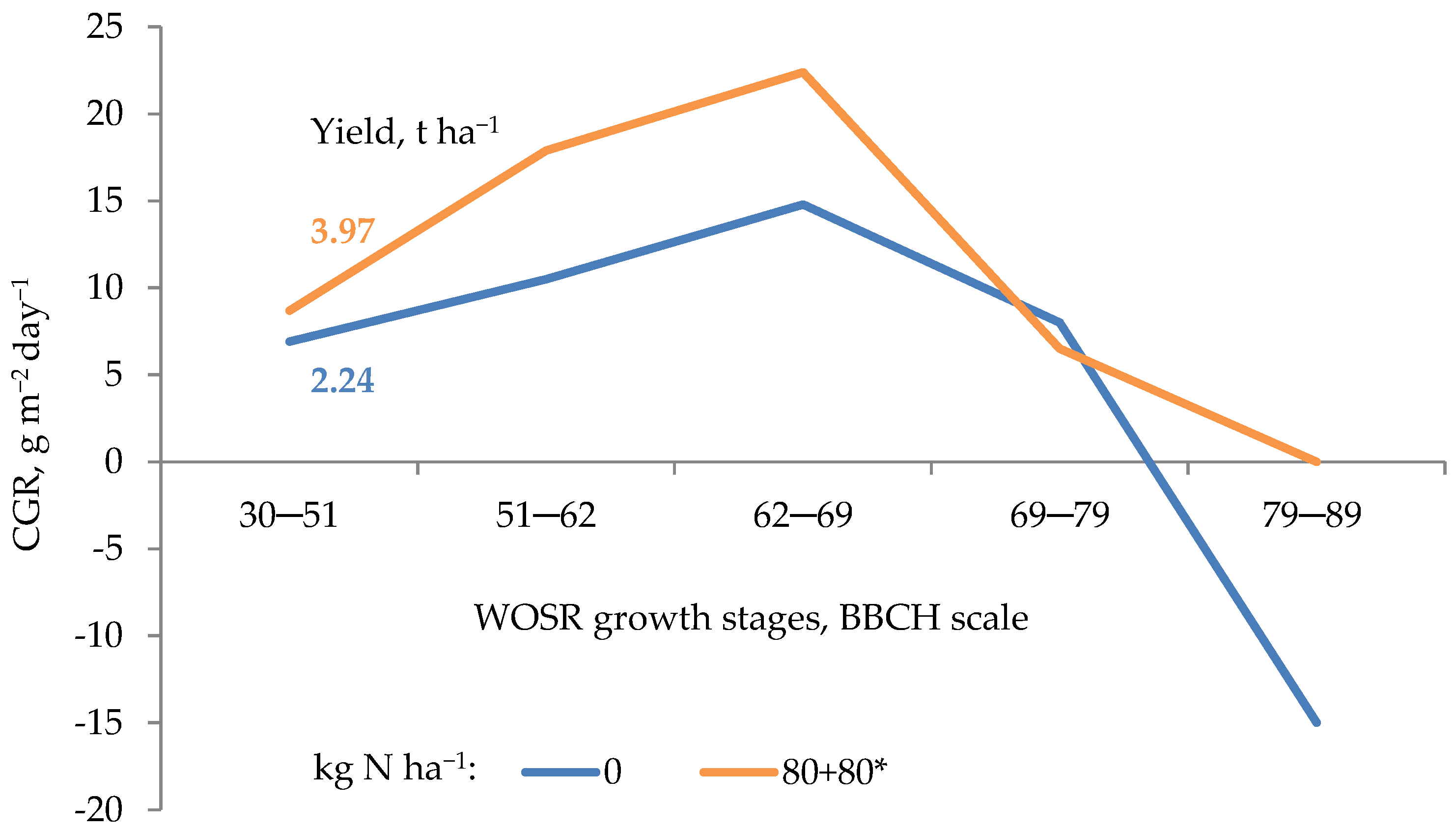
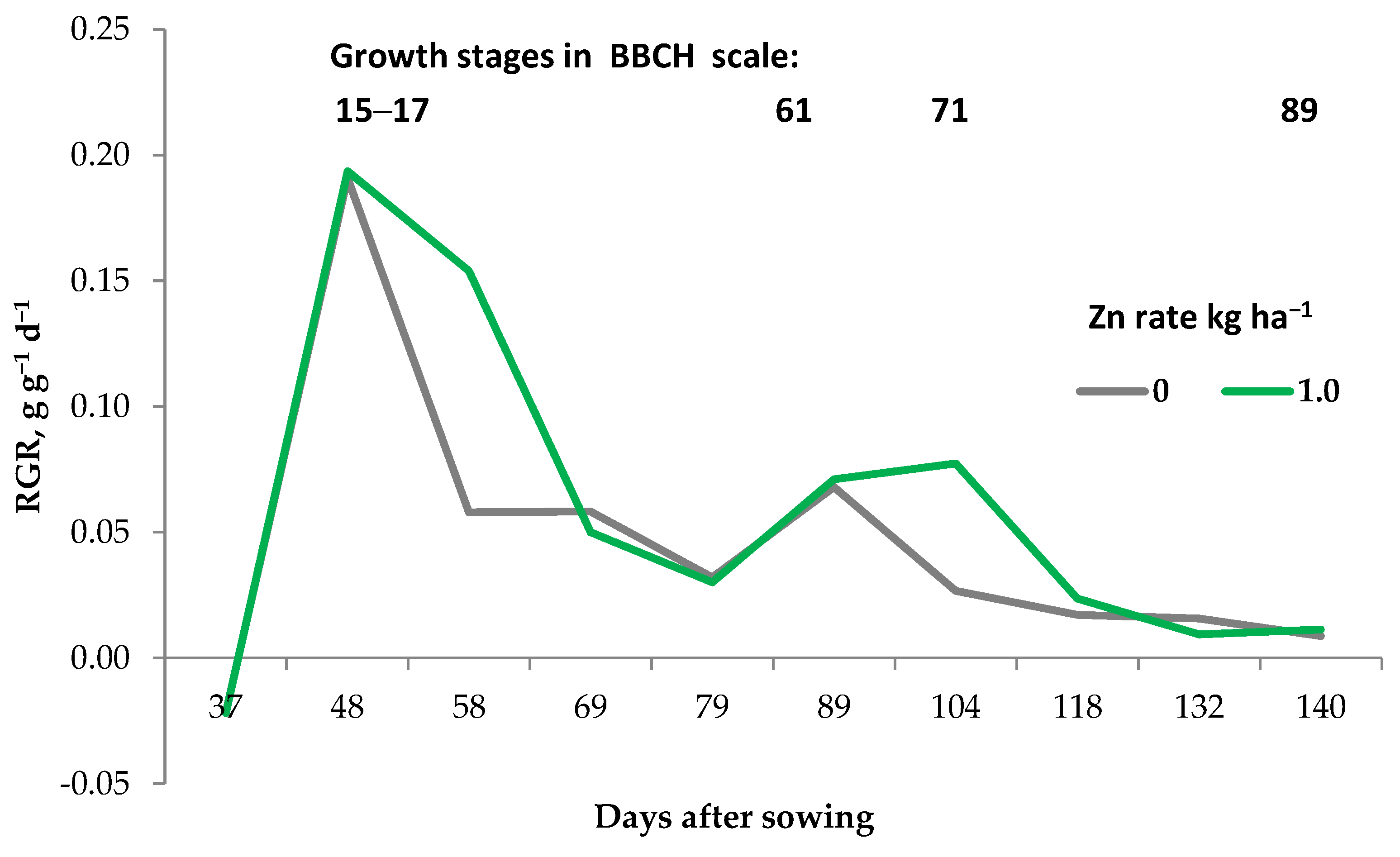
2.3. Root System Growth during the Growing Season
- (a)
- metabolic requirements of the above-ground parts of a plant are higher than the rate of nutrient supply to the plant from the soil solution;
- (b)
- the effective diffusion of a nutrient is sufficiently high;
- (c)
- the time of the plant root interaction with the soil in a particular soil zone is long enough.
3. Soil Factors Affecting FUE
3.1. Soil Texture
3.2. Water Content
3.3. Soil Compaction
3.4. Soil Temperature
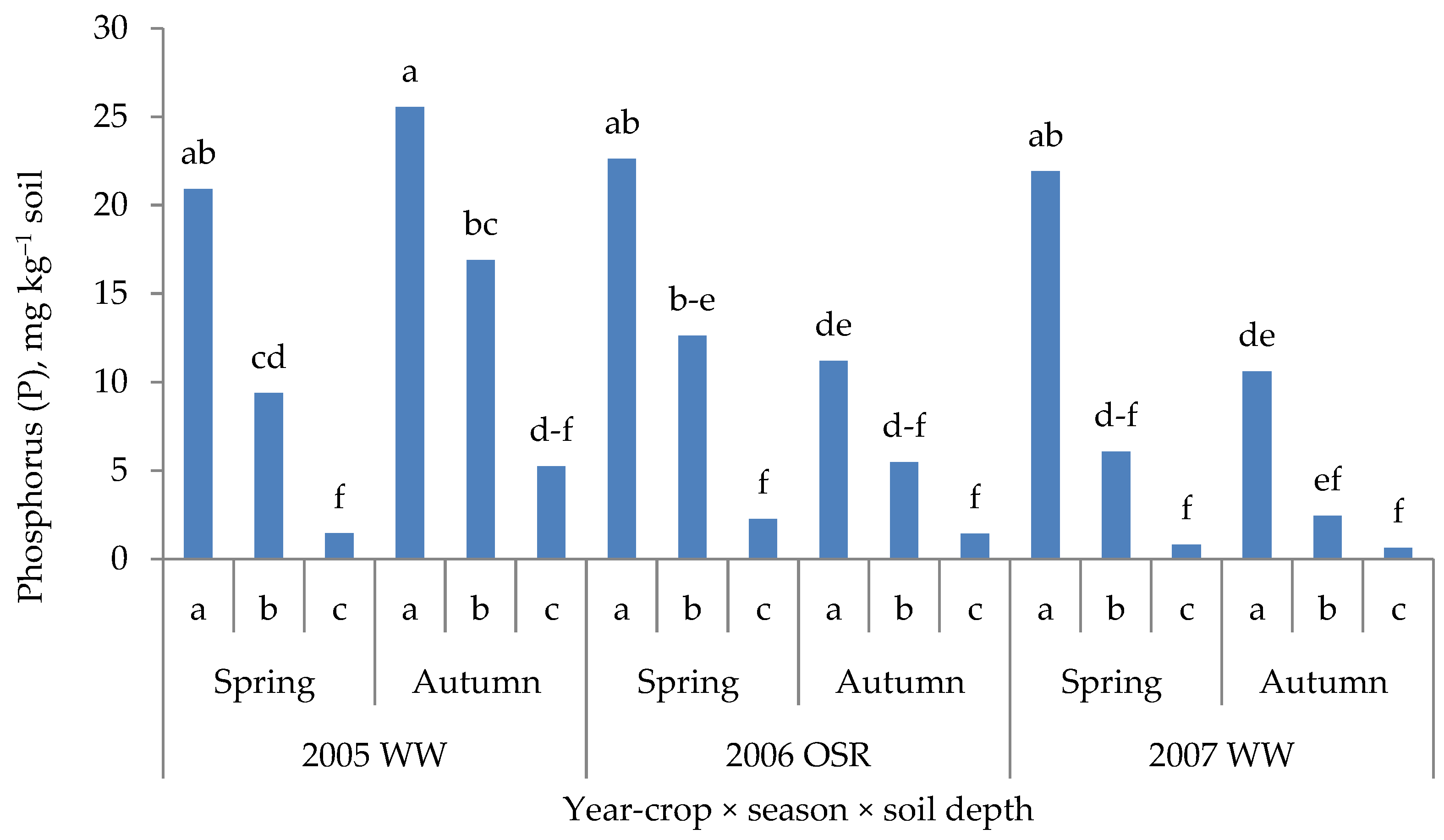
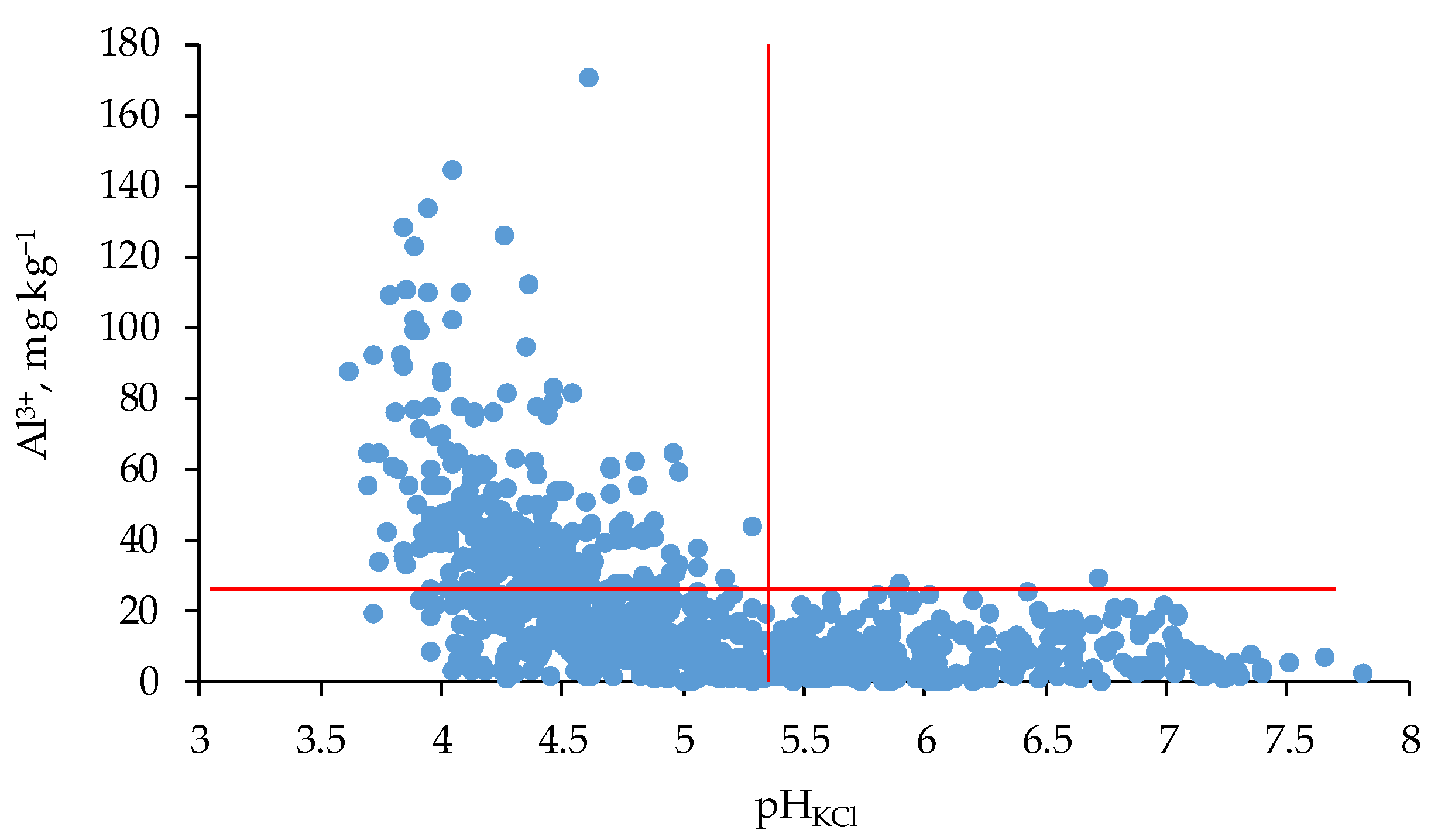
3.5. Soil Reaction
3.6. Soil Salinity
3.7. Soil Organic Matter
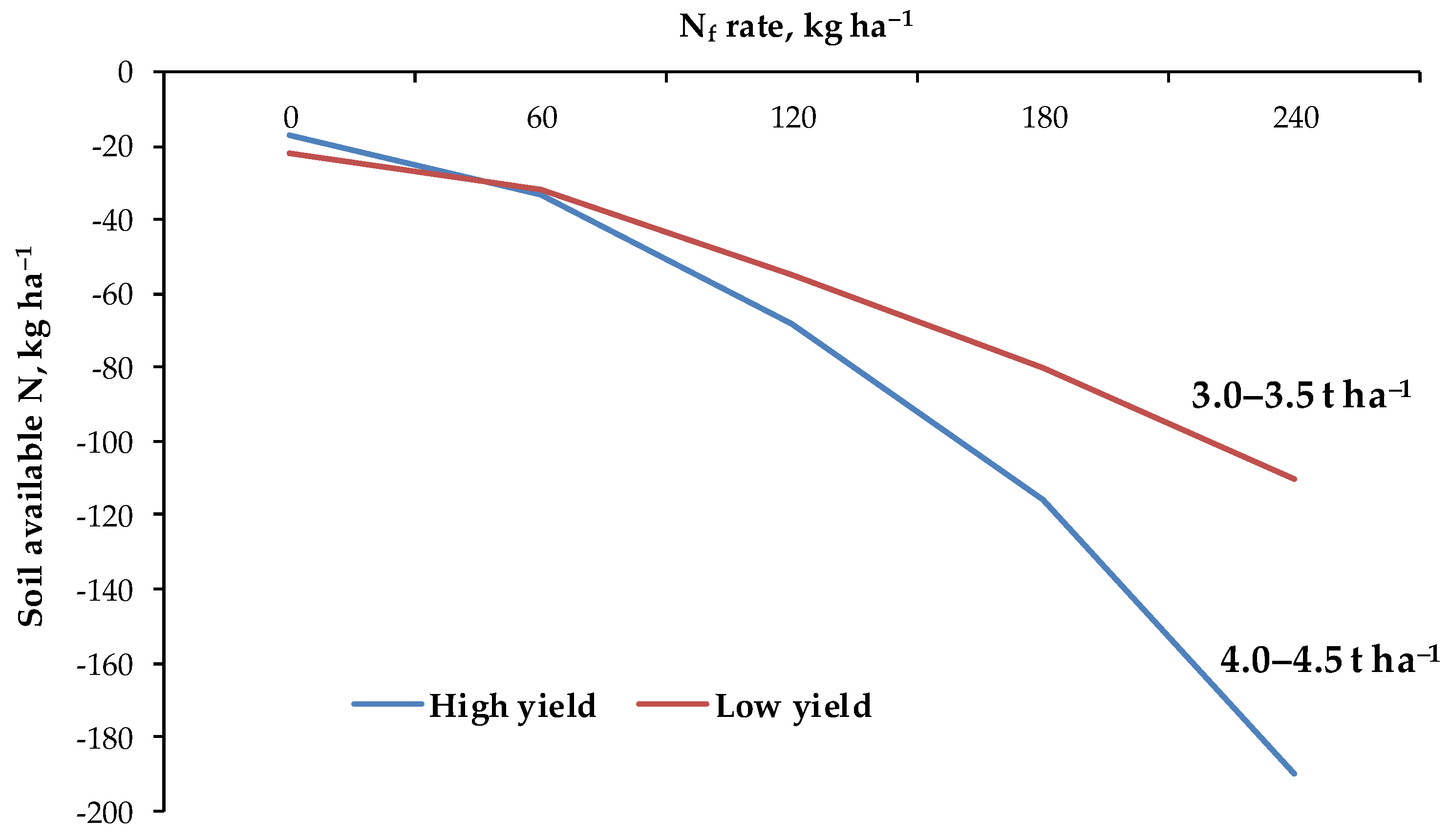
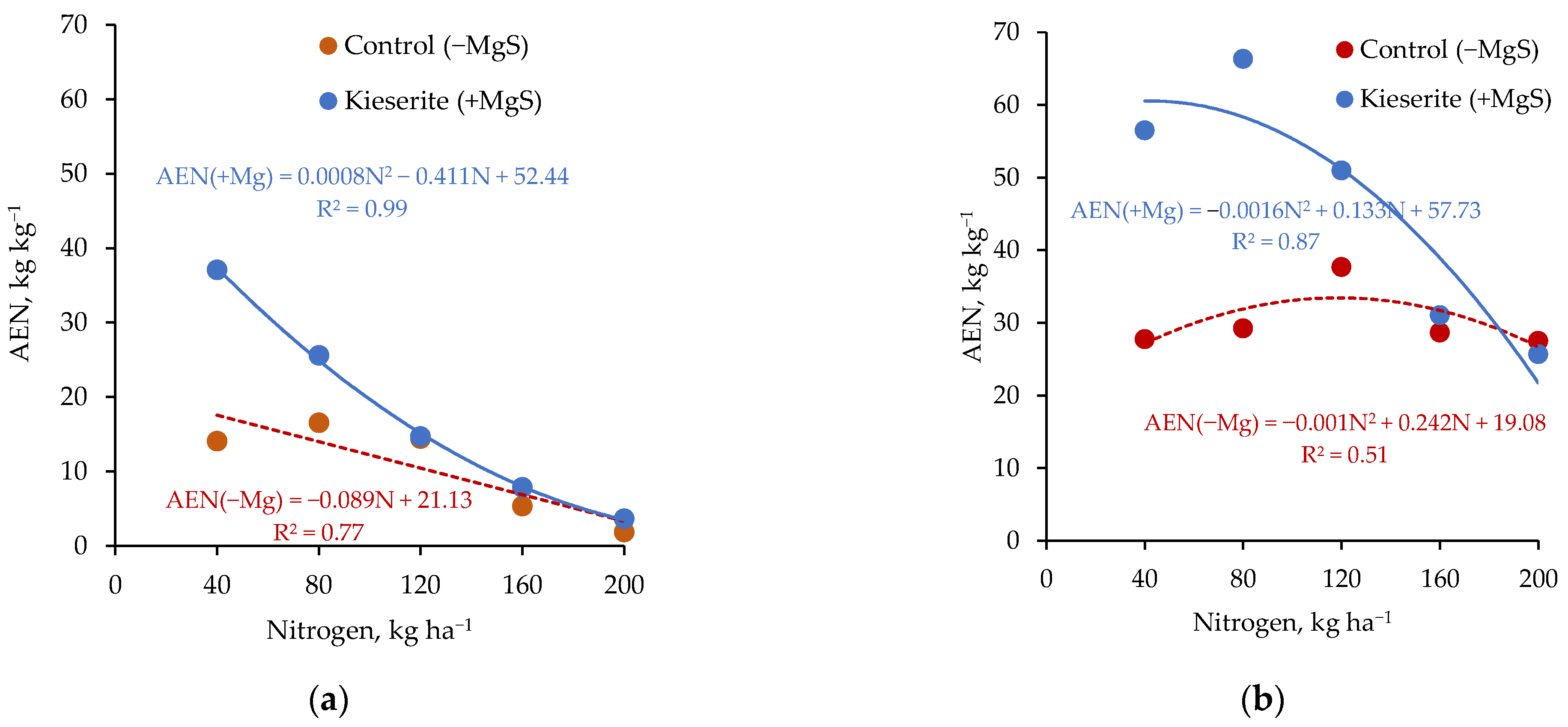
3.8. Nutrient Shortage
4. Innovations on the Fertilizer Market
5. FUE—A Message for Agricultural Practice
- Determine the maximum attainable yield (Yattmax). This is the basis for choosing the most suitable variety for the actual climatic and soil conditions of the farm;
- Identify soil conditions that constrain:
- growth and architecture of the root system
- water and nutrient availability;
- Divide the whole field area into units of homogenous productivity;
- Identify Nitrogen Hotspots both on the farm and on the specific field;
- Observe the viability of plants at stages preceding the cardinal phases of yield formation;
- Schedule the correction of the plant nutritional status during the season to exploit its yield potential.
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Licker, R.; Johnston, M.; Foley, J.A.; Barford, C.; Kucharik, C.J.; Monfreda, C.; Ramankutty, N. Mind the gap: How do climate and agricultural management explain the ‘yield gap’ of croplands around the world? Glob. Ecol. Biogeogr. 2010, 19, 769–782. [Google Scholar] [CrossRef]
- Tandzi, N.L.; Mutengwa, S.C. Factors affecting yields of crops. In Agronomy—Climate Change and Food Security; Amanullah, Ed.; IntechOpen: London, UK, 2020; p. 16. [Google Scholar]
- Sattari, S.Z.; Van Ittersum, M.K.; Bouwman, A.F.; Smit, A.L.; Janssen, B.H. Crop yield response to soil fertility and N, P, K inputs in different environments: Testing and improving the QEFTS model. Field Crops Res. 2014, 157, 35–46. [Google Scholar] [CrossRef]
- Lollato, R.P.; Edwards, J.T. Maximum attainable wheat yield and resources-use efficiency in the Southern Great Plains. Crop Sci. 2015, 55, 2863–2875. [Google Scholar] [CrossRef] [Green Version]
- Wallace, A.; Wallace, G.A. Closing the Crop-Yield Gap through Better Soil and Better Management; Wallace Laboratories: Los Angeles, CA, USA, 2003; p. 162. [Google Scholar]
- Grzebisz, W.; Łukowiak, R.; Sassenrath, G. Virtual nitrogen as a tool for assessment of nitrogen at the field scale. Field Crops Res. 2018, 218, 182–184. [Google Scholar] [CrossRef]
- Dobermann, A.R. Nitrogen use efficiency—State of the art. In Agronomy & Horticulture Faculty Publications, Proceedings of the IFA International Workshop on Enhanced Efficiency Fertilizers, Frankfurt, Germany, 28–30 June 2005; University of Nebraska–Lincoln: Lincoln, NE, USA, 2005; Volume 316, pp. 1–16. [Google Scholar]
- Grzebisz, W.; Łukowiak, R. Nitrogen Gap Amelioration is a Core for Sustainable Intensification of Agriculture—A Concept. Agronomy 2021, 11, 419. [Google Scholar] [CrossRef]
- Fixen, P.; Brentrup, F.; Bruulsema, T.; Garcia, F.; Norton, R.; Zingore, S. Nutrient/fertilizer use efficiency: Measurement, current situation and trends. In Managing Water and Fertilizer for Sustainable Agricultural Intensification; Drechsel, P., Heffer, P., Magen, H., Mikkelsen, R., Wichelns, D., Eds.; IFA: Paris, France, 2015; pp. 8–38. [Google Scholar]
- Oenema, O.; Kros, H.; De Vries, W. Approaches and uncertainties in nutrient budgets: Implications for nutrient management and environmental policies. Eur. J. Agron. 2003, 20, 3–16. [Google Scholar] [CrossRef]
- Pradhan, P.; Fischer, G.; van Velthuizen, H.; Reusser, D.E.; Kropp, J.P. Closing yield gaps: How sustainable can we be? PLoS ONE 2015, 10, e0129487. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants; Elsevier: Cambridge, MA, USA; Academic Press: London, UK , 1995; p. 899. [Google Scholar]
- Parry, A.J.; Andralojc, P.J.; Scales, J.C.; Salvucci, M.E.; Carmo-Silva, A.E.; Alonso, H.; Whitney, S.M. Rubisco activity and regulation as targets for crop improvement. J. Exp. Bot. 2013, 6493, 717–730. [Google Scholar] [CrossRef]
- Guan, P. Dancing with hormones: A current perspective of nitrate signaling and regulation in Arabidopsis. Front. Plant Sci. 2017, 8, 1697. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture. J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef]
- Marschner, P. (Ed.) Marchnerr’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; p. 672. [Google Scholar]
- Blecharczyk, A.; Zawada, D.; Sawińska, Z.; Małcka-Jankowiak, I.; Waniorek, W. Impact of crop seqeunce and fetilization on yield of winter wheat. Fragm. Agron. 2019, 36, 27–35. [Google Scholar]
- Tabak, M.; Lepiarczyk, A.; Filipek–Mazur, B.; Lisowska, A. Efficiency of Nitrogen Fertilization of Winter Wheat Depending on Sulfur Fertilization. Agronomy 2020, 10, 1304. [Google Scholar] [CrossRef]
- Roberts, T.L. Improving Nutrient Use Efficiency. Turk. J. Agric. For. 2008, 32, 177–182. [Google Scholar]
- Zhang, F.; Niu, J.; Zhang, W.; Chen, X.; Li, C.; Yuan, L.; Xie, J. Potassium nutrition of crops under varied regimes of nitrogen supply. Plant Soil 2010, 335, 21–34. [Google Scholar] [CrossRef]
- Schulte Auf’m Erley, G.; Behrens, T.; Ulas, A.; Wiesler, F.; Horst, W.J. Agronomic traits contributing to nitrogen efficiency of winter oilseed rape cultivars. Field Crops Res. 2011, 124, 114–123. [Google Scholar] [CrossRef]
- Barłóg, P.; Grzebisz, W. Effect of timing and nitrogen fertilizer application on winter oilseed rape (Brassica napus L.). I. Growth dynamics and seed yield. J. Agron. Crop Sci. 2004, 190, 305–313. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J.; Przygocka-Cyna, K. Nutritional status of winter oilseed rape in cardinal stages of growth as the yield indicator. Plant Soil Environ. 2015, 61, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Wild, A.; Breeze, V. Nutrient uptake in relation to growth. In Physiological Processes Limiting Plant Productivity; Johnston, C.B., Ed.; Butterworths: London, UK, 1981; pp. 331–345. [Google Scholar]
- Meyer, U. BBCH Monograph. In Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Center for Agriculture and Forestry: Berlin, Germany, 2001; Available online: http://www.jki.bund.de/fileadmin/dam_uploads/_veroeff/bbch/BBCH-Skala_Englisch.pdf (accessed on 14 March 2021).
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H. A flexible sigmoid function of determinate growth. Ann. Bot. 2003, 91, 361–371. [Google Scholar] [CrossRef]
- Sylvester-Bradley, R.; Lunn, G.; Foulkes, J.; Shearman, V.; Spink, J.; Ingram, J. Management Strategies for Yield of Cereals and Oilseed Rape. In Proceedings of the 18 HGCA Conference on Agronomic Intelligence: The Basis for Profitable Production, London, UK, 16–17 January 2002; Available online: www.hgca.com/publications (accessed on 14 May 2022).
- Bergmann, W. Nutritional Disorders of Plants; Gustav Fischer Verlag: Jena, Germany, 1992; p. 741. [Google Scholar]
- Ritchie, S.W.; Hanway, J.J.; Benson, G.O. How a Corn Plant Develops; Special Report No. 48; Iowa State University of Science and Technology Cooperative Extension Service: Ames, IA, USA, 1986; p. 21. [Google Scholar]
- Subedi, K.; Ma, B. Nitrogen uptake and partitioning in stay-green and leafy maize hybrids. Crop. Sci. 2005, 45, 740–747. [Google Scholar] [CrossRef]
- Grzebisz, W.; Wrońska, M.; Diatta, J.B.; Szczepaniak, W. Effect of zinc foliar application at early stage of maize growth on the patterns of nutrients and dry matter accumulation by the canopy. Part II: Nitrogen uptake and dry matter accumulation patterns. J. Elem. 2008, 13, 29–39. [Google Scholar]
- Elwali, A.M.O.; Gascho, G.J.; Summer, M.E.E. Dris norm for 11 nutrients in corn leaves. Agron. J. 1985, 77, 506–508. [Google Scholar] [CrossRef]
- Potarzycki, J. Influence of balanced fertilization on nutritional status of maize at anthesis. Fertil. Fertil. 2010, 39, 90–108. (In Polish) [Google Scholar]
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J. An assessment of the effect of potassium fertilizing systems on maize nutritional status in critical stages of growth by plant analysis. J. Elem. 2014, 19, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Grzebisz, W.; Łukowiak, R.; Kotnis, K. Evaluation of nitrogen fertilization systems based on the in-season variability of nitrogenous growth factors and soil fertility factors: A case of winter oilseed rape (Brassica napus L.). Agronomy 2020, 10, 1701. [Google Scholar] [CrossRef]
- Klepper, B.; Rickman, R.W.; Waldman, S.; Chevalier, P. The physiological life cycle of wheat: Its use in breeding and crop management. Euphytica 1998, 100, 341–347. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, D.; Schnurbusch, T. Plant and floret growth at distinct developmental stages during the stem elongation phase in wheat. Front. Plant Sci. 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Belete, F.; Dechassa, N.; Molla, A.; Tana, T. Effect of split application of different N rates on productivity and nitrogen use effciency of bread wheat (Triticum aestivum L.). Agric. Food Sec. 2018, 7, 92. [Google Scholar] [CrossRef]
- Beaufils, E.R. Diagnosis and Recommendation Integrated System (DRIS): A General Scheme of Experimentation and Calibration Based on Principles Developed from Research in Plant Nutrition; Soil Science Bulletin 1; University of Natal, Department of Soil Science and Agrometeorology: Pietermaritzburg, South Africa, 1973; p. 132. [Google Scholar]
- Parent, L.E.; Dafir, M.A. Theoretical concept of compositional nutrient diagnosis. J. Am. Soc. Hortic. Sci. 1992, 117, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.R.; Plank, C.O. Reference sufficiency ranges field crops—Corn. In Reference Sufficiency Ranges for Plant Analysis in the Southern Region of the United States; Southern Cooperative Series Bulletin 394; Campbell, C.R., Ed.; Southern Region Agricultural Experiment Station: Fayetteville, AR, USA, 2000. Available online: http://www.ncagr.gov/agronomi/saaesd/corn.htm (accessed on 9 June 2022).
- Soltanpour, P.N.; Malakouti, M.J.; Ronaghi, A. Comparison of diagnosis and recommendation integrated system and nutrient sufficiency range for corn. Soil Sci. Soc. Am. J. 1995, 59, 133–139. [Google Scholar] [CrossRef]
- Barłóg, P. Diagnosis of sugar beet (Beta vulgaris L.) nutrient imbalance by DRIS and CND-clr methods at two stages during early growth. J. Plant Nutr. 2016, 39, 1–16. [Google Scholar] [CrossRef]
- Frąckowiak, K.; Potarzycki, J.; Grzebisz, W.; Szczepaniak, W. Potato nutritional status at the onset of tuberization—A yield prediction tool. Plant Soil Environ. 2020, 66, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Schulte, E.E.; Kelling, K.A. Analysis: A Diagnostic Tool; NCH-46 Crop Fertilization 4/91; University of Wisconsin-Madison: Madison, WI, USA, 1992; Available online: http://ces.purdue.edu/extmedia/NCH/NCH-46.html (accessed on 11 May 2012).
- Jones, J.B.; Eck, H.V.; Voss, R. Plant analysis as an aid in fertilizing corn and grain sorghum. In Soil Testing and Plant Analysis, 3rd ed.; Westerman, R.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1990; pp. 521–527. [Google Scholar]
- Reid, R.; Hayes, J. Mechanisms and control of nutrient uptake in plants. Int. Rev. Cytol. 2003, 229, 73–114. [Google Scholar] [PubMed]
- Kell, D.B. Breeding crop plants with deep roots: Their role in sustainable carbon, nutrient and water sequestration. Ann. Bot. 2011, 108, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Gemenet, D.C.; Villordon, A. Root system architecture and abiotic stress tolerance: Current knowledge in root and tuber crops. Front. Plant Sci. 2016, 7, 1584. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.J. Are plant roots only “in” soil or are they “of’ soil it? Roots, soil formation and function. Eur. J. Soil Sci. 2022, 73, e13219. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef]
- Fradgley, N.; Evans, G.; Biernaskie, J.M.; Cockram, J.; Marr, E.C.; Oliver, A.G.; Ober, E.; Jones, H. Effects of breeding history and crop management on the root architecture of wheat. Plant Soil 2020, 452, 587–600. [Google Scholar] [CrossRef]
- Rich, S.M.; Watt, M. Soil conditions and cereal root system architecture: Review and considerations for linking Darwin and Weaver. J. Exp. Bot. 2013, 64, 1193–1208. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.L. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Gerwitz, A.; Page, R.E. An empirical mathematical model to describe plant root systems. J. Appl. Ecol. 1974, 11, 773–781. [Google Scholar] [CrossRef]
- Fan, J.; McConkey, B.; Wang, H.; Janzen, H. Roots distribution by depth for temperate agricultural crops. Field Crops Res. 2016, 189, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Metselaar, K.; Rodrigues Pinheiro, E.A.; De Jong van Lier, Q. mathematical descriptio of rooting profiles of agricultural crops and its efefct on transpiration prediction by a hyrological model. Soil Syst. 2019, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Wojciechowski, T. opportunities and challegens in the subsoil: Pathways to deeper rooted crops. J. Exp. Bot. 2015, 66, 2199–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renger, M.; Strebel, O. Jährliche Grundwasserneubildung in Abhängigkeit von Bodennutzung und Bodeneigenschaften. Wasser Boden 1980, 32, 362–366. [Google Scholar]
- Haberle, J.; Svoboda, P. Calculation of available water supply in crop root zone ant the water balance of crops. Contr. Geophys. Geod. 2015, 45, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Olfs, H.-W.; Blankenau, K.; Brentrup, F.; Jasper, J.; Link, A.; Lammel, J. Soil- and plant-based nitrogen-fertilizer recommendations in arable farming. J. Plant Nutr. Soil Sci. 2005, 168, 414–431. [Google Scholar] [CrossRef]
- Kautz, T.; Amelung, W.; Ewert, F.; Gaiser, T.; Horn, R.; Jahn, R.; Javaux, M.; Kemna, A.; Kuzyakova, Y.; Munch, J.-C.; et al. Nutrient acquisition from arable subsoils in temperate climates: A review. Soil Biol. Biochem. 2013, 57, 1003–1022. [Google Scholar] [CrossRef]
- Siebers, N.; Wang, L.; Funk, T.; Von Tucher, S.; Merbach, I.; Schweitzer, K.; Kruse, J. Subsoils—A sink for excess fertilizer P but a minor contribution to P plant nutrition: Evidence from long-term fertilization trials. Environ. Sci. Eur. 2021, 33, 60. [Google Scholar] [CrossRef]
- Barłóg, P.; Łukowiak, R.; Grzebisz, W. Predicting the content of soil mineral nitrogen based on the content of calcium chloride-extractable nutrients. J. Plant Nutr. Soil Sci. 2017, 180, 624–635. [Google Scholar] [CrossRef]
- Łukowiak, R.; Grzebisz, W.; Sassenrath, G.F. New insights into phosphorus management in agriculture—A crop rotation approach. Sci. Total Environ. 2016, 542, 1062–1077. [Google Scholar] [CrossRef] [Green Version]
- Giehl, R.F.H.; Gruber, B.D.; Von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Van Noordwijk, M.; Dde Willigen, P. Root functions in agricultural soils. In Plant Roots and Their Environment; Developments in Agricultural and Managed Forest Ecology 24; McMichael, B.L., Persson, H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1991; p. 649. [Google Scholar]
- Kurepa, J.; Smalle, J.A. Auxin/cytokinin antagonistic control of the shoot/root growth ration and its relevance for adaptation to growth and nutrient deficiency stress. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef] [PubMed]
- McMichael, B.L.; Burke, J.J. Soil temperaturę and root growth. HortScience 1998, 33, 947–951. [Google Scholar] [CrossRef]
- Grzebisz, W. The sepecific adsorption rate and nitrogen absorption rate of winter rye grown in the long-term fertilization experiment. Zesz. Probl. Postępów Nauk. Rol. 1995, 421, 123–135. (In Polish) [Google Scholar]
- Grzebisz, W.; Kryszak, J. Effect of soil fertility on root morphology of winter rye. In Root Ecology and Its Practical Application; Kutschera, L., Hübl, E., Lichtenegger, E., Persson, H., Sobotik, M., Eds.; Verein fot Wurzelforschung: Klagenfurt, Austria, 1992; pp. 389–392. [Google Scholar]
- Passioura, J. Increasing crop productivity when water is scarce—From breeding to field management. Agric. Water Manag. 2006, 80, 176–196. [Google Scholar] [CrossRef] [Green Version]
- Beeckman, T.; Friml, J. Nitrate contra auxin: Nutrient sensing by roots. Dev. Cell 2010, 18, 877–878. [Google Scholar] [CrossRef] [PubMed]
- Ötvös, K.; Marconi, M.; Vega, A.; O’brian, J.; Johnston, A.; Abualla, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.; Yan, S.; et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Durand, J.-L.; Gastal, F. Water deficit and nitrogen nutrition of crops: A review. Agron. Sustain. Dev. 2010, 30, 529–544. [Google Scholar] [CrossRef] [Green Version]
- Raynaud, X.; Leadley, P.W. Soil characteristics play a key role in modeling nutrient competition in plant communities. Ecology 2004, 85, 2200–2214. [Google Scholar] [CrossRef]
- Clarkson, D.T. Nutrient interception and transport by root system. In Physiological Processes Limiting Plant Productivity; Johnson, C.B., Ed.; Butterworths: London, UK, 1981; pp. 307–330. [Google Scholar]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1995. [Google Scholar]
- Noordwijk, M. Functional interpretation of root densities in the field for nutrient and water uptake. In Wurzeloekologie and Ihre Nutzwendung; Instituut voor Bodemvruchtbaarheid: Irdning, Austria, 1983; pp. 207–226. [Google Scholar]
- White, C.; Sylvester-Bradley, R.; Berry, P.M. Root length densities of UK wheat and oilseed rape crops with implications for water capture and yield. J. Exp. Bot. 2015, 66, 2293–2303. [Google Scholar] [CrossRef] [Green Version]
- Barraclough, P.B. Root growth, macro-nutrient uptake dynamics and soil fertility requirements of a high-yielding winter oilseed rape crop. Plant Soil 1989, 119, 59–70. [Google Scholar] [CrossRef]
- Krouk, G.; Ruffel, S.; Gutiérrez, R.A.; Gojon, A.; Crawford, N.M.; Coruzzi, G.M.; Lacombe, B. A framework integrating plant growth with hormones and nutrients. Trends Plant Sci. 2011, 16, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.-Q.; Shu, J.-Q.; Li, W.-M.; Wang, G.-Z. Role of auxin and nitrate signaling in the development of root system architecture. Front. Plant Sci. 2021, 12, 690363. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.T. Soils: Principles, Properties and Management; Springer: Dordrecht, The Netherlands, 2013; p. 271. [Google Scholar] [CrossRef]
- Kome, G.K.; Enang, R.K.; Tabi, F.O.; Yerima, B.P.K. Influence of Clay Minerals on Some Soil Fertility Attributes: A Review. Open J. Soil Sci. 2019, 9, 155–188. [Google Scholar] [CrossRef] [Green Version]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef]
- Merino, C.; Nannipieri, P.; Matus, F. Soil Carbon Controlled by Plant, Microorganism and Mineralogy Interactions. J. Soil Sci. Plant Nutr. 2015, 15, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, B.; Singh, M.; Mandal, S.; Churchman, G.J.; Bolan, N.S. Clay Minerals—Organic Matter Interactions in Relation to Carbon Stabilization in Soils. In The Future of Soil Carbon: Its Conservation and Formation; Garcia, C., Nannipieri, P., Hernandez, T., Eds.; Academic Press: London, UK, 2018; Chapter 3; pp. 71–86. [Google Scholar]
- Lal, R. Soil organic matter and water retention. Agron. J. 2020, 112, 3265–3277. [Google Scholar] [CrossRef]
- Carter, M.R. Soil quality for sustainable land management: Organic matter and aggregation interactions that maintain soil function. Agron. J. 2002, 94, 38–47. [Google Scholar] [CrossRef]
- Raheb, A.; Heidari, A. Effects of Clay Mineralogy and Physicochemical Properties on Potassium Availability under Soil Aquic Conditions. J. Soil Sci. Plant Nutr. 2012, 12, 747–761. [Google Scholar]
- Moraru, S.S.; Ene, A.; Badila, A. Physical and Hydro-Physical Characteristics of Soil in the Context of Climate Change. A Case Study in Danube River Basin, SE Romania. Sustainability 2020, 12, 9174. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Nieder, R.; Dinesh, K.B.; Scherer, H.W. Fixation and Defixation of Ammonium in Soils: A Review. Biol. Fertil. Soils 2011, 47, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Parent, L.E.; MacLeod, J.A. Influence of soil texture on fertilizer and soil phosphorus transformations in Gleysolic soils. Can. J. Soil Sci. 2003, 83, 395–403. [Google Scholar] [CrossRef]
- Wilson, M.J. Weathering of the primary rock-formingminerals: Processes, products and rates. Clay Miner. 2004, 39, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Erlandsson Lampa, M.; Sverdrup, H.U.; Bishop, K.H.; Belyazid, S.; Ameli, A.; Köhler, S.J. Catchment export of base cations: Improved mineral dissolution kinetics influence the role of water transit time. Soil 2020, 6, 231–244. [Google Scholar] [CrossRef]
- Gavrilescu, M. Water, Soil, and Plants Interactions in a Threatened Environment. Water 2021, 13, 2746. [Google Scholar] [CrossRef]
- Trinh, T.H.; KuShaari, K.; Basit, A. Modeling the Release of Nitrogen from Controlled-Release Fertilizer with Imperfect Coating in Soils and Water. Ind. Eng. Chem. Res. 2015, 54, 6724–6733. [Google Scholar] [CrossRef]
- Alaoui, I.; El-ghadraoui, O.; Serbouti, S.; Ahmed, H.; Mansouri, I.; El-Kamari, F.; Taroq, A.; Ousaaid, D.; Squalli, W.; Farah, A. The Mechanisms of Absorption and Nutrients Transport in Plants: A Review. Trop. J. Nat. Prod. Res. 2022, 6, 8–14. [Google Scholar] [CrossRef]
- Comerford, N.B. Soil Factors Affecting Nutrient Bioavailability. In Nutrient Acquisition by Plants; Ecological Studies; Bassiri, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 181, p. 14. [Google Scholar]
- Oliveira, E.M.M.; Ruiz, H.A.; Alvarez, V.V.H.; Ferreira, P.A.; Costa, F.O.; Almeida, I.C.C. Nutrient supply by mass flow and diffusion to maize plants in response to soil aggregate size and water potential. Rev. Bras. Ciência Solo 2020, 34, 317–327. [Google Scholar] [CrossRef]
- Gransee, A.; Führs, H. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil 2013, 368, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Hinsinger, P.; Brauman, A.; Devau, N.; Gérard, F.; Jourdan, C.; Laclau, J.P.; Le Cadre, E.; Jaillard, B.; Plassard, C. Acquisition of phosphorus and other poorly mobile nutrients by roots. Where do plant nutrition models fail? Plant Soil 2011, 348, 29–61. [Google Scholar]
- Rengel, Z. Availability of Mn, Zn and Fe in the rhizosphere. J. Soil Sci. Plant Nutr. 2015, 15, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Mauceri, A.; Bassolino, L.; Lupini, A.; Badeck, F.; Rizza, F.; Schiavi, M. Genetic variation in eggplant for nitrogen use efficiency under contrasting NO3− supply. J. Integr. Plant Biol. 2020, 62, 393–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurel, C.; Nacry, P. Root architecture and hydraulics converge for acclimation to changing water availability. Nat. Plants 2020, 6, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Haling, R.E.; Brown, L.K.; Bengough, A.G.; Young, I.M.; Hallett, P.D.; White, P.J.; George, T.S. Root hairs improve root penetration, root-soil contact, and phosphorus acquisition in soils of different strength. J. Exp. Bot. 2013, 64, 3711–3721. [Google Scholar] [CrossRef] [Green Version]
- Tron, S.; Bodner, G.; Laio, F.; Ridolfi, L.; Leitner, D. Can diversity in root architecture explain plant water use efficiency? A modeling study. Ecol. Model. 2015, 312, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.M.; Chen, Y. Moderate Drought Stress Affected Root Growth and Grain Yield in Old, Modern and Newly Released Cultivars of Winter Wheat. Front. Plant Sci. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupini, A.; Preiti, G.; Badagliacca, G.; Abenavoli, M.R.; Sunseri, F.; Monti, M.; Bacchi, M. Nitrogen Use Efficiency in Durum Wheat Under Different Nitrogen and Water Regimes in the Mediterranean Basin. Front. Plant Sci. 2021, 11, 607226. [Google Scholar] [CrossRef]
- Becker, M.; Asch, F. Iron toxicity in rice—Conditions and management concepts. J. Plant Nutr. Soil Sci. 2005, 168, 558–573. [Google Scholar] [CrossRef]
- Vepraskas, M.J. Plant response mechanisms to soil compaction. In Plant-Environment Interactions; Wilkonson, R.E., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 263–287. [Google Scholar]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Correa, J.; Postma, J.A.; Watt, M.; Wojciechowski, T. Soil compaction and the architectural plasticity of root systems. J. Exp. Bot. 2019, 70, 6019–6034. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.J.; Whalley, W.R.; Barraclough, P.B. How do roots penetrate strong soil? Plant Soil 2003, 255, 93–104. [Google Scholar] [CrossRef]
- Whalley, W.R.; Bengough, A.G.; Dexter, A.R. Water stress induced by PEG decreases the maximum growth pressure of the roots of pea seedlings. J. Exp. Bot. 1998, 49, 1689–1694. [Google Scholar] [CrossRef]
- Iijima, M.; Morita, S.; Barlow, P.W. Structure and function of the root cap. Plant Prod. Sci. 2008, 11, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Hunbury, C.D.; Atwell, B.J. Growth dynamics of mechanically impeded lupin roots: Does altered morphology induce hypoxia? Ann. Bot. 2005, 96, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.; Shen, J.; Ashton, R.W.; Dodd, I.C.; Parry, M.A.; Whalley, W.R. How do roots elongate in a structured soil? J. Exp. Bot. 2013, 64, 4761–4777. [Google Scholar] [CrossRef]
- Bengough, A.G. Root elongation is restricted by axial but not by radial pressures: So what happens in field soil? Plant Soil 2012, 360, 15–18. [Google Scholar] [CrossRef]
- Ramalingam, P.; Kamoshita, A.; Deshmukh, V.; Yaginuma, S.; Uga, Y. Association between root growth angle and root length density of a near-isogenic line of IR64 rice with DEEPER ROOTING 1 under different levels of soil compaction. Plant Prod. Sci. 2017, 20, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Thorup-Kristensen, K.; Cortasa, M.S.; Loges, R. Winter wheat roots grow twice as deep as spring wheat roots, is this important for N uptake and N leaching losses? Plant Soil 2009, 322, 101–114. [Google Scholar] [CrossRef]
- Batey, T. Soil compaction and soil management—A review. Soil Use Manag. 2009, 25, 335–345. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Valentine, T.A.; Hallett, P.D.; Binnie, K.; Young, M.W.; Squire, G.R.; Hawes, C.; Bengough, A.G. Soil strength and macropore volume limit root elongation rates in many UK agricultural soils. Ann. Bot. 2012, 110, 259–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitaula, B.K.; Hansen, S.; Sitaula, J.I.B.; Bakken, L.R. Effects of soil compaction on N2O emission in agricultural soil. Chemosphere-Glob. Chang. Sci. 2000, 2, 367–371. [Google Scholar] [CrossRef]
- Chamindu Deepagoda, T.K.K.; Clough, T.J.; Thomas, S.M.; Balaine, N.; Elberling, B. Density Effects on Soil-Water Characteristics, Soil-Gas Diffusivity, and Emissions of N2O and N2 from a Re-packed Pasture Soil. Soil Sci. Soc. Am. J. 2018, 83, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Ruser, R.; Flessa, H.; Russow, R.; Schmidt, G.; Buegger, F.; Munch, J.C. Emission of N2O, N2 and CO2 from soil fertilized with nitrate: Effect of compaction, soil moisture and rewetting. Soil Biol. Biochem. 2006, 38, 263–274. [Google Scholar] [CrossRef]
- Soane, B.; Van Ouwerkerk, C. Implications of soil compaction in crop production for the quality of the environment. Soil Till Res. 1995, 35, 5–22. [Google Scholar] [CrossRef]
- Sommer, S.G.; Hutchings, N.J. Ammonia emission from field applied manure and its reduction. Eur. J. Agron. 2001, 15, 1–15. [Google Scholar] [CrossRef]
- Onwuka, B.; Mang, B. Effects of soil temperature on some soil properties and plant growth. Adv. Plants Agric. Res. 2018, 8, 34–37. [Google Scholar] [CrossRef]
- Fang, C.M.; Smith, P.; Moncrieff, J.B.; Smith, J.U. Similar response of labile and resistant soil organic matter pools to changes in temperature. Nature 2005, 436, 881–883. [Google Scholar] [CrossRef] [Green Version]
- Elbasiouny, H.; El-Ramady, H.; Elbehiry, F.; Rajput, V.D.; Minkina, T.; Mandzhieva, S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability 2022, 14, 914. [Google Scholar] [CrossRef]
- Gahoonia, T.S.; Nielsen, N.E. Phosphorus uptake and growth of root hairless barley mutant (bald root barley) and wild type in low and high–p soils. Plant Cell Environ. 2003, 26, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Yilvainio, K.; Pettovuori, T. Phosphorus acquisition by barley (Hordeum yulgar) at suboptimal soil temperature. Agric. Food Sci. 2012, 21, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Lahti, M.; Aphalo, P.; Finér, L.; Ryyppö, A.; Lehto, T.; Mannerkoski, H. Effects of soil temperature on shoot and root growth and nutrient uptake of 5-year-old Norway spruce seedlings. Tree Physiol. 2005, 25, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavelienė, V.; Jurkonienė, S.; Jankovska-Bortkevič, E.; Švegždienė, D. Effects of Elevated Temperature on Root System Development of Two Lupine Species. Plants 2022, 11, 192. [Google Scholar] [CrossRef]
- Huang, B.R.; Taylor, H.M.; McMichael, B.L. Effects of Temperature on the Development of Metaxylem in Primary Wheat Roots and Its Hydraulic Consequence. Ann. Bot. 1991, 67, 163–166. [Google Scholar] [CrossRef]
- Falah, M.A.F.; Wajima, T.; Yasutake, D.; Sago, Y.; Kitano, M. Responses of root uptake to high temperature of tomato plants (Lycopersicon esculentum Mill.) in soil-less culture. J. Agric. Technol. 2010, 6, 543–558. [Google Scholar]
- Xia, Z.; Zhang, S.; Wang, Q.; Zhang, G.; Fu, Y.; Lu, H. Effects of Root Zone Warming on Maize Seedling Growth and Photosynthetic Characteristics Under Different Phosphorus Levels. Front. Plant Sci. 2021, 12, 746152. [Google Scholar] [CrossRef]
- Fageria, N.K.; Barbosa Filho, M.B. Influence of pH on Productivity, Nutrient Use Efficiency by Dry Bean, and Soil Phosphorus Availability in a No-Tillage System. Commun. Soil Sci. Plant Anal. 2008, 39, 1016–1025. [Google Scholar] [CrossRef]
- Jackson, K.; Meetei, T.T. Influence of soil pH on nutrient availability: A Review. J. Emerg. Technol. Innov. Res. 2018, 5, 708–713. [Google Scholar]
- Neina, D. The Role of Soil pH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Bojórquez-Quintal, E.; Escalante-Magaña, C.; Echevarría-Machado, I.; Martínez-Estévez, M. Aluminum, a friend or foe of higher plants in acid soils. Front. Plant Sci. 2017, 8, 1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Błaszyk, R. Effects of Lime Fertilizer Reactivity on Selected Chemical Properties of Soil and Crop Yielding in Two Different Tillage Systems. Doctoral Thesis, Department of Agricultural Chemistry and Environmental Biogeochemistry, Poznan University of Life Sciences, Poznań, Poland, 2020; p. 237. (In Polish). [Google Scholar]
- McCauley, A.; Jones, C.; Jacobsen, J. Soil pH and Organic Matter. Montana State University (MSU) Extension. Available online: https://apps.msuextension.org/publications/pub.html?sku=4449-8 (accessed on 26 June 2022).
- Rangel, A.F.; Madhusudana, R.I.; Johannes, H.W. Intracellular distribution and binding state of aluminum in root apices of two common bean (Phaseolus vulgaris) genotypes in relation to Al toxicity. Physiol. Plant. 2009, 135, 162–173. [Google Scholar] [CrossRef]
- Rahman, M.A.; Lee, S.H.; Ji, H.C.; Kabir, A.H.; Jones, C.S.; Lee, K.W. Importance of Mineral Nutrition for Mitigating Aluminum Toxicity in Plants on Acidic Soils: Current Status and Opportunities. Int. J. Mol. Sci. 2018, 19, 3073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, J.; Babourina, O.; Rengel, Z. Role of magnesium in alleviation of aluminum toxicity in plants. J. Exp. Bot. 2011, 62, 2251–2264. [Google Scholar] [CrossRef] [Green Version]
- Čiamporová, M. Morphological and Structural Responses of Plant Roots to Aluminium at Organ, Tissue, and Cellular Levels. Biol. Plant. 2002, 45, 161–171. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, D.K.; Singh, S.; Sharma, S.; Dubey, N.K.; Chauhan, D.K.; Vaculík, M. Toxicity of aluminium on various levels of plant cells and organism: A review. Environ. Exp. Bot. 2017, 137, 177–193. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F. Aluminum–Nitrogen Interactions in the Soil–Plant System. Front. Plant Sci. 2018, 9, 807. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Wang, J.; Xue, J.; Yue, Z.; Xia, S.; Song, L.; Gao, H. Effects of Soil pH on Gaseous Nitrogen Loss Pathway via Feammox Process. Sustainability 2021, 13, 10393. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Teixeira, D.M.; Pinto, A.P.; Brito, I.; Barrulas, P.; Carvalho, M. Aluminium, Iron and Silicon Subcellular Redistribution in Wheat Induced by Manganese Toxicity. Appl. Sci. 2021, 11, 8745. [Google Scholar] [CrossRef]
- Stavi, I.; Thevs, N.; Priori, S. Soil Salinity and Sodicity in Drylands: A Review of Causes, Effects, Monitoring, and Restoration Measures. Front. Environ. Sci. 2021, 9, 712831. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanavelu, A.; Naganna, S.R.; Al-Ansari, N. Irrigation Induced Salinity and Sodicity Hazards on Soil and Groundwater: An Overview of Its Causes, Impacts and Mitigation Strategies. Agriculture 2021, 11, 983. [Google Scholar] [CrossRef]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic Adjustment and Energy Limitations to Plant Growth in saline Soil. New Phytol. 2019, 225, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahriaripour, R.; Tajabadi Pour, A.; Mozaffari, V. Effects of Salinity and Soil Phosphorus Application on Growth and Chemical Compositiono Pistachio Seedlings. Commun. Soil Sci. Plant Anal. 2011, 42, 144–158. [Google Scholar] [CrossRef]
- Rengasamy, P. Soil processes affecting crop production in salt-affected soils. Funct. Plant Biol. 2010, 37, 613–620. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849–858. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Raddatz, N.; Morales de los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Rosales, M.A.; Franco-Navarro, J.D.; Peinado-Torrubia, P.; Díaz-Rueda, P.; Álvarez, R.; Colmenero-Flores, J.M. Chloride Improves Nitrate Utilization and NUE in Plants. Front. Plant Sci. 2020, 11, 442. [Google Scholar] [CrossRef]
- Acikbas, S.; Ozyazici, M.A.; Bektas, H. The Effect of Salinity on Root Architecture in Forage Pea (Pisum sativum ssp. arvense L.). Legume Res. 2021, 44, 407–412. [Google Scholar] [CrossRef]
- Lal, R. Soil health and carbon management. Food Energy Secur. 2016, 5, 212–222. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Dalal, R.C.; Finn, D.; Menzies, N.W. Global changes in soil stocks of carbon, nitrogen, phosphorus, and sulphur as influenced by long-term agricultural production. Glob. Chang. Biol. 2017, 23, 2509–2519. [Google Scholar] [CrossRef] [PubMed]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Yang, W.; Ding, W.; Jia, L.; Jiang, L.; Liu, Y.; Xu, X.; Yang, Y.; He, P.; Yang, J. Estimation of Nitrogen Supply for Summer Maize Production through a Long-Term Field Trial in China. Agronomy 2021, 11, 1358. [Google Scholar] [CrossRef]
- Luis, L.; Gilles, B.; Bruna, G.; Juliette, A.; Josette, G. 50 year trends in nitrogen use efficiency of world cropping systems: The relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar]
- Wei, X.; Shao, M.; Gale, W.; Li, L. Global pattern of soil carbon losses due to the conversion of forests to agricultural land. Sci. Rep. 2014, 4, 4062. [Google Scholar] [CrossRef]
- Xue, Z.; An, S. Changes in Soil Organic Carbon and Total Nitrogen at a Small Watershed Scale as the Result of Land Use Conversion on the Loess Plateau. Sustainability 2018, 10, 4757. [Google Scholar] [CrossRef] [Green Version]
- Livsey, J.; Alavaisha, E.; Tumbo, M.; Lyon, S.W.; Canale, A.; Cecotti, M.; Lindborg, R.; Manzoni, S. Soil Carbon, Nitrogen and Phosphorus Contents along a Gradient of Agricultural Intensity in the Kilombero Valley, Tanzania. Land 2020, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Haddaway, N.R.; Hedlund, K.; Jackson, L.E.; Kätterer, T.; Lugato, E.; Thomsen, I.K.; Jørgensen, H.B.; Isberg, P.E. How does tillage intensity afect soil organic carbon? A systematic review. Environ. Evid. 2017, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Krauss, M.; Wiesmeier, M.; Don, A.; Cuperus, F.; Gattinger, A.; Gruberf, S.; Haagsma, W.K.; Peign’e, J.; Chiodelli Palazzoli, M.; Schulz, F.; et al. Reduced tillage in organic farming affects soil organic carbon stocks in temperate Europe. Soil Tillage Res. 2022, 2016, 105262. [Google Scholar] [CrossRef]
- Powlson, D.S.; Bhogal, A.; Chambers, B.J.; Coleman, K.; Macdonald, A.J.; Goulding, W.T.; Whitmore, A.P. The potential to increase soil carbon stocks through reduced tillage or organic material additions in England and Wales: A case study. Agric. Ecosyst. Environ. 2012, 146, 23–33. [Google Scholar] [CrossRef]
- Szajdak, L.; Życzyńska-Bałoniak, I.; Meysner, T.; Blecharczyk, A. Bound amino acids in humic acids from arable cropping systems. J. Plant Nutr. Soil Sci. 2004, 167, 562–567. [Google Scholar] [CrossRef]
- Khalil, M.I.; Rahman, M.S.; Schmidhalter, U.; Olfs, H.-W. Nitrogen fertilizer–induced mineralization of soil organic C and N in six contrasting soils of Bangladesh. J. Plant Nutr. Soil Sci. 2007, 170, 210–218. [Google Scholar] [CrossRef]
- Barre, P.; Montagnier, C.; Chenu, C.; Abbadie, L.; Velde, B. Clay minerals as a soil potassium reservoir: Observation and quantification through X-ray diffraction. Plant Soil 2008, 302, 213–220. [Google Scholar] [CrossRef]
- Khan, H.R.; Elahi, S.F.; Hussain, M.S.; Adachi, T. Soil Characteristics and Behavior of Potassium under Various Moisture Regimes. Soil Sci. Plant Nutr. 1994, 40, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Pogłodziński, R.; Barłóg, P.; Grzebisz, W. Effect of nitrogen and magnesium sulfate application on sugar beet yield and quality. Plant Soil Environ. 2021, 67, 507–517. [Google Scholar] [CrossRef]
- The Royal Society. Ammonia: Zero-Carbon Fertiliser, Fuel and Energy Store: Policy Briefing. 2020. Available online: https://royalsociety.org/-/media/policy/projects/green-ammonia/green-ammonia-policy-briefing.pdf (accessed on 5 May 2022).
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef]
- Trenkel, M.E. Slow- and Controlled-Release and Stabilized Fertilizers: An Option for Enhancing Nutrient Use Efficiency in Agriculture; International Fertilizer Industry Association (IFA): Paris, France, 2010; p. 160. [Google Scholar]
- Rashid, M.; Hussain, Q.; Khan, K.S.; Alwabel, M.I.; Hayat, R.; Akmal, M.; Ijaz, S.S.; Alvi, S.; Rehman, O.U. Carbon-Based Slow-Release Fertilizers for Efficient Nutrient Management: Synthesis, Applications, and Future Research Needs. J. Soil Sci. Plant Nutr. 2021, 21, 1144–1169. [Google Scholar] [CrossRef]
- Wesołowska, M.; Rymarczyk, J.; Góra, R.; Baranowski, P.; Sławiński, C.; Klimczyk, M.; Supryn, G.; Schimmelpfennig, L. New slow-release fertilizers—Economic, legal and practical aspects: A Review. Int. Agrophys. 2021, 35, 11–24. [Google Scholar] [CrossRef]
- Hojjatie, M.M. Urea triazone fertilizers-A slow-release nitrogen fertilizer. J. Agric. Sci. Food Technol. 2021, 7, 272–276. [Google Scholar]
- Guo, Y.; Zhang, M.; Liu, Z.; Zhao, C.; Lu, H.; Zheng, L.; Li, Y.C. Applying and Optimizing Water-Soluble, Slow-Release Nitrogen Fertilizers for Water-Saving Agriculture. ACS Omega 2020, 5, 11342–11351. [Google Scholar] [CrossRef] [PubMed]
- Medina, L.C.; Sartain, J.B.; Obreza, T.A.; Hall, W.L.; Thiex, N.J. Evaluation of a Soil Incubation Method to Characterize Nitrogen Release Patterns of Slow- and Controlled-Release Fertilizers. J. AOAC Int. 2014, 97, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Azeem, B.; Kushaari, K.; Man, Z.B.; Basit, A.; Thanh, T.H. Review on materials & methods to produce controlled release coated urea fertilizer. J. Control. Release 2014, 181, 11–21. [Google Scholar]
- Roshanravan, B.; Soltani, S.M.; Mahdavi, F.; Rashid, S.A.; Yusop, M.K. Preparation of encapsulated urea-kaolinite controlled release fertiliser and their effect on rice productivity. Chem. Speciat. Bioavailab. 2014, 26, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Majeed, Z.; Ramli, N.K.; Mansor, N.; Man, Z. A comprehensive review on biodegradable polymers and their blends used in controlled-release fertilizer processes. Rev. Chem. Eng. 2015, 31, 69–95. [Google Scholar] [CrossRef]
- Ma, X.; Chen, J.; Yang, Y.; Su, X.; Zhang, S.; Gao, B.; Li, L.C. Siloxane and polyether dual modification improves hydrophobicity and interpenetrating polymer network of bio-polymer for coated fertilizers with enhanced slow release characteristics. Chem. Eng. J. 2018, 350, 1125–1134. [Google Scholar] [CrossRef]
- Beig, B.; Niazi, M.B.K.; Jahan, Z.; Hussain, A.; Zia, M.H.; Mehran, M.T. Coating materials for slow release of nitrogen from urea fertilizer: A review. J. Plant Nutr. 2020, 43, 1510–1533. [Google Scholar] [CrossRef]
- Thind, H.S.; Bijay, S.; Pannu, R.P.S.; Yadvinder, S.; Varinderpal, S. Relative performance of neem (Azadirachta indica) coated ureavis-a-vis ordinary urea applied to rice on the basis of soil test orfollowing need based nitrogen management using leaf colour chart. Nutr. Cycl. Agroecosyst. 2010, 87, 1–8. [Google Scholar] [CrossRef]
- Shivay, Y.S.; Prasad, R.; Singh, R.K.; Pal, M. Relative efficiency of zinc-coated urea and soil and foliar application of zinc sulphate on yield, nitrogen, phosphorus, potassium, zinc and iron biofortification in grains and uptake by basmati rice (Oryza sativa L.). J. Agric. Sci. 2015, 7, 161. [Google Scholar] [CrossRef] [Green Version]
- Ghafoor, I.; Habib-ur-Rahman, M.; Ali, M.; Afza, M.; Ahmed, W.; Gaiser, T.; Ghaffar, A. Slow-release nitrogen fertilizers enhance growth, yield, NUE in wheat crop and reduce nitrogen losses under an arid environment. Environ. Sci. Pollut. Res. 2021, 28, 43528–43543. [Google Scholar] [CrossRef]
- Wen, P.; Wu, Z.; Han, Y.; Cravotto, G.; Wang, J.; Ye, B. Microwaveassisted synthesis of a novel biochar based slow release nitrogen fertilizer with enhanced water-retention capacity. ACS Sustain. Chem. Eng. 2017, 5, 7374–7382. [Google Scholar] [CrossRef]
- Gil-Ortiz, R.; Naranjo, M.A.; Ruiz-Navarro, A.; Atares, S.; García, C.; Zotarelli, L.; Bautista, A.S.; Vicente, S. Enhanced Agronomic Efficiency Using a New Controlled-Released, Polymeric-Coated Nitrogen Fertilizer in Rice. Plants 2020, 9, 1183. [Google Scholar] [CrossRef]
- Guo, Y.; Ma, Z.; Ren, B.; Zhao, B.; Liu, P.; Zhang, J. Effects of Humic Acid Added to Controlled-Release Fertilizer on Summer Maize Yield, Nitrogen Use Efficiency and Greenhouse Gas Emission. Agriculture 2022, 12, 448. [Google Scholar] [CrossRef]
- Shaviv, A. Advances in controlled-release fertilizers. Adv. Agron. 2001, 71, 1–49. [Google Scholar]
- Lawrencia, D.; Wong, S.K.; Low, D.Y.S.; Goh, B.H.; Goh, J.K.; Ruktanonchai, U.R.; Soottitantawat, A.; Lee, L.H.; Tang, S.Y. Controlled Release Fertilizers: A Review on Coating Materials and Mechanism of Release. Plants 2021, 10, 238. [Google Scholar] [CrossRef]
- Panáková, Z.; Slamka, P.; Ložek, O. Effect of nitrification inhibitors on the content of available nitrogen forms in the soil under maize (Zea mays, L.) growing. J. Cent. Eur. Agric. 2016, 17, 1013–1032. [Google Scholar] [CrossRef]
- Khalil, M.J.; Gutser, R.; Schmidhalter, U. Effects of urease and nitrification inhibitors added to urea on nitrous oxide emissions from a loess soil. J. Plant Nutr. Soil Sci. 2009, 172, 651–660. [Google Scholar] [CrossRef]
- Chien, S.H.; Prochnow, L.I.; Cantarella, H. Recent developments of fertilizer production and use to improve nutrient efficiency and minimize environmental impacts. Adv. Agron. 2009, 102, 267–322. [Google Scholar]
- Rehman, H.U.; Alharby, H.F.; Al-Zahrani, H.S.; Bamagoos, A.A.; Alsulami, N.B.; Alabdallah, N.M.; Iqbal, T.; Wakeel, A. Enriching Urea with Nitrogen Inhibitors Improves Growth, N Uptake and Seed Yield in Quinoa (Chenopodium quinoa Willd) Affecting Photochemical Efficiency and Nitrate Reductase Activity. Plants 2022, 11, 371. [Google Scholar] [CrossRef]
- Yang, M.; Fang, Y.; Sun, D.; Shi, Y. Efficiency of two nitrification inhibitors (dicyandiamide and 3,4-dimethypyrazole phosphate) on soil nitrogen transformations and plant productivity: A meta-analysis. Sci. Rep. 2016, 6, 22075. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Ayuso, M.; Gabriel, J.L.; Quemada, M. Nitrogen use efficiency and residual effect of fertilizers with nitrification inhibitors. Eur. J. Agron. 2016, 80, 1–8. [Google Scholar] [CrossRef] [Green Version]
- d’Onofrio, G.; Dupuy, J.; Gaudin, R. Comparative effects of the application of prilled urea and urea supergranules on flooded rice in Madagascar. J. Agric. Stud. 2018, 6, 218–233. [Google Scholar] [CrossRef] [Green Version]
- Fertahi, S.; Bertrand, I.; Amjoud, M.; Oukarroum, A.; Arji, M.; Barakat, A. Properties of Coated Slow-Release Triple Superphosphate (TSP) Fertilizers Based on Lignin and Carrageenan Formulations. ACS Sustain. Chem. Eng. 2019, 7, 10371–10382. [Google Scholar] [CrossRef]
- Sofyane, A.; Lahcini, M.; El Meziane, A.; Khouloud, M.; Dahchour, A.; Caillol, S.; Raihane, M. Properties of Coated Controlled Release Diammonium Phosphate (DAP) Fertilizers Prepared with the Use of Bio-based Amino Oil. J. Am. Oil Chem. Soc. 2020, 97, 751–763. [Google Scholar] [CrossRef]
- Fertahi, S.; Bertranda, I.; Ilsouk, M.; Oukarroum, A.; Amjoud, B.; Zerouald, Y.; Barakat, A. New generation of controlled release phosphorus fertilizers based on biological macromolecules: Effect of formulation properties on phosphorus release. Int. J. Biol. Macromol. 2020, 143, 153–162. [Google Scholar] [CrossRef]
- Barbosa, C.F.; Correa, D.A.; Carneiro, J.S.D.S.; Melo, L.C.A. Biochar Phosphate Fertilizer Loaded with Urea Preserves Available Nitrogen Longer than Conventional Urea. Sustainability 2022, 14, 686. [Google Scholar] [CrossRef]
- Saied, H.S.H.; Aboelenin, S.M.; Kesba, H.; El-Sherbieny, A.E.A.; Helmy, A.M.; Dahdouh, S.M.; Soliman, M.M. Chemical evaluation of partially acidulated phosphate rocks and their impact on dry matter yield and phosphorus uptake of maize. Saudi J. Biol. Sci. 2022, 29, 3511–3518. [Google Scholar] [CrossRef]
- Bulina, N.V.; Makarova, S.V.; Baev, S.G.; Matvienko, A.A.; Gerasimov, K.B.; Logutenko, O.A.; Bystrov, V.S. A Study of Thermal Stability of Hydroxyapatite. Minerals 2021, 11, 1310. [Google Scholar] [CrossRef]
- Essah, S.Y.C. Application of Phosphorus Fertilizer with ‘NutriCharge’ Improves Potato Tuber Yield and Phosphorus Use Efficiency. Available online: https://potatoes.colostate.edu/wp-content/uploads/2020/01/2019-Article-for-Spud-Item.pdf (accessed on 13 June 2022).
- Palanivell, P.; Ahmed, O.H.; Omar, L.; Abdul Majid, N.M. Nitrogen, Phosphorus, and Potassium Adsorption and Desorption Improvement and Soil Buffering Capacity Using Clinoptilolite Zeolite. Agronomy 2021, 11, 379. [Google Scholar] [CrossRef]
- Rafique, M.; Sultan, T.; Ortas, I.; Chaudhary, H.J. Enhancement of maize plant growth with inoculation of phosphate-solubilizing bacteria and biochar amendment in soil. J. Soil Sci. Plant Nutr. 2017, 63, 460–469. [Google Scholar] [CrossRef]
- Ali, M.S.; Sutradhar, A.; Edano, M.L.; Edwards, J.T.; Girma, K. Response of Winter Wheat Grain Yield and Phosphorus Uptake to Foliar Phosphite Fertilization. Int. J. Agron. 2014, 2014, 801626. [Google Scholar] [CrossRef] [Green Version]
- Souri, M.K. Aminochelate fertilizers: The new approach to the old problem; a review. Open Agric. 2016, 1, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, W.F.; Fagan, E.B.; Soares, L.H.; Soares, J.N.; Reichardt, K.; Neto, D.D. Seed and Foliar Application of Amino Acids Improve Variables of Nitrogen Metabolism and Productivity in Soybean. Crop. Front. Plant Sci. 2018, 9, 396. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Deng, X.; Wang, B.; Zhang, N.; Zhu, C.; Li, R.; Shen, Q. Effects of foliar application of amino acid liquid fertilizers, with or without Bacillus amyloliquefaciens SQR9, on cowpea yield and leaf microbiota. PLoS ONE 2019, 14, e0222048. [Google Scholar] [CrossRef]
- Nargesi, M.M.; Sedaghathoor, S.; Hashemabadi, D. Effect of foliar application of amino acid, humic acid and fulvic acid on the oil content and quality of olive. Saudi J. Biol. Sci. 2022, 29, 3473–3481. [Google Scholar] [CrossRef]
- Gondek, K.; Mierzwa-Hersztek, M. Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy 2021, 11, 2582. [Google Scholar] [CrossRef]
- Ramli, R.A. Slow release fertilizer hydrogels: A review. Polym. Chem. 2019, 10, 6073–6090. [Google Scholar] [CrossRef]
- Ullah, F.; Othman, M.B.H.; Javed, F.; Ahmad, Z.; Akil, H.M. Classification, processing and application od hydrogels: A review. Mater. Sci. Eng. C 2015, 57, 414–433. [Google Scholar] [CrossRef]
- Kareem, S.A.; Dere, I.; Gungula, D.T.; Andrew, F.P.; Saddiq, A.M.; Adebayo, E.F.; Tame, V.T.; Kefas, H.M.; Joseph, J.; Patrick, D.O. Synthesis and Characterization of Slow-Release Fertilizer Hydrogel Based on Hydroxy Propyl Methyl Cellulose, Polyvinyl Alcohol, Glycerol and Blended Paper. Gels 2021, 7, 262. [Google Scholar] [CrossRef]
- Michalik, R.; Wandzik, I. A Mini-Review on Chitosan-Based Hydrogels with Potential for Sustainable Agricultural Applications. Polymers 2020, 12, 2425. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Joshi, A.; Tian, D.-D.; Rajput, V.D.; Singh, M.; Arora, J.; Minkina, T.; Li, Y.-R. Recent Trends in Nano-Fertilizers for Sustainable Agriculture under Climate Change for Global Food Security. Nanomaterials 2022, 12, 173. [Google Scholar] [CrossRef]
- Commission Recommendation of 18 October 2011 on the Definition of Nanomaterial. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32011H0696 (accessed on 13 June 2022).
- Shalaby, T.A.; Bayoumi, Y.; Eid, Y.; Elbasiouny, H.; Elbehiry, F.; Prokisch, J.; El-Ramady, H.; Ling, W. Can Nanofertilizers Mitigate Multiple Environmental Stresses for Higher Crop Productivity? Sustainability 2022, 14, 3480. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Almoshadak, A.S.; Shafi, M.E.; Albagami, N.M.; Saad, A.M.; El-Tahan, A.M.; El-Sayed, M.D.; Elnahal, A.S.M.; Almakas, A.; Abd El-Mageed, A.A.; et al. Vital roles of sustainable nano-fertilizers in improving plant quality and quantity-an updated review. Saudi J. Biol. Sci. 2021, 28, 7349–7359. [Google Scholar] [CrossRef]
- Babu, S.; Singh, R.; Yadav, D.; Rathore, S.S.; Raj, R.; Avasthe, R.; Yadave, S.K.Y.; Das, A.; Yadav, V.; Yadav, B.; et al. Nanofertilizers for agricultural and environmental sustainability. Chemosphere 2022, 292, 133451. [Google Scholar] [CrossRef]
- Bhardwaj, A.K.; Arya, G.; Kumar, R.; Hamed, L.; Pirasteh-Anosheh, H.; Poonam Jasrotia, P.; Kashyap, P.L.; Singh, G.P. Switching to nanonutrients for sustaining agroecosystems and environment: The challenges and benefits in moving up from ionic to particle feeding. J. Nanobiotechnol. 2022, 20, 19. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosh, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Bernela, M.; Rani, R.; Malik, P.; Mukherjeed, T.K. Nanofertilizers: Applications and Future Prospects. In Nanotechnology: Principles and Applications; Sindhu, R.K., Chitkara, M., Sandhu, I.S., Eds.; Jenny Stanford Publishing Pte. Ltd.: Singapore, 2021; Chapter 9; pp. 289–332. [Google Scholar]
- Manikandan, A.; Subramanian, K.S. Evaluation of zeolite-based nitrogen nano-fertilizers on maize growth, yield and quality on inceptisols and alfisols. Int. J. Plant Soil Sci. 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.M.M.; Hasaneen, M.N.A.; Omer, A.M. Nano chitosan-NPK fertilizer enhances the growth and productivity of wheat plants grown in sandy soil. Span. J. Agric. Res. 2016, 14, e0902. [Google Scholar] [CrossRef]
- Anand, R.; Bhagat, M. Silver nanoparticles (AgNPs): As nanopesticides and nanofertilizers. MOJ Biol. Med. 2019, 4, 19–20. [Google Scholar] [CrossRef]
- Kottegoda, N.; Sandaruwan, C.; Priyadarshana, G.; Siriwardhana, A.; Rathnayake, U.A.; Berugoda Arachchige, D.M.; Kumarasinghe, A.R.; Dahanayake, D.; Karunaratne, V.; Amaratunga, G.A.J. Urea-hydroxyapatite nanohybrids for slow release of nitrogen. ACS Nano 2017, 11, 1214–1221. [Google Scholar] [CrossRef]
- Raguraj, S.; Wijayathunga, V.M.S.; Gunaratne, G.P.; Amali, R.K.A.; Priyadarshana, G.; Sandaruwan, C.; Karunaratne, V.; Hettiarachchi, L.S.K.; Kottegoda, N. Urea–hydroxyapatite nanohybrid as an efficient nutrient source in Camelliasinensis (L.) Kuntze (tea). J. Plant Nutr. 2020, 43, 2383–2394. [Google Scholar] [CrossRef]
- Li, Q.; Ma, C.; White, J.C.; Xing, B. Effects of Phosphorus Ensembled Nanomaterials on Nutrient Uptake and Distribution in Glycine max L. under Simulated Precipitation. Agronomy 2021, 11, 1086. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Lombi, E.; Wang, P.; Schjoerring, J.K.; Husted, S. Nanomaterials as fertilizers for improving plant mineral nutrition and environmental outcomes. Environ. Sci. Nano 2019, 6, 3513. [Google Scholar] [CrossRef]
- Husen, A.; Siddiqi, K.S. Carbon and fullerene nanomaterials in plant System. J. Nanobiotechnol. 2014, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.C.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef] [Green Version]
- Schauberger, B.; Ben-Ari, T.; Makowski, D.; Kato, T.; Kato, H.; Ciais, P. Yield trends, variability and stagnation analysis of major crops in France over more than a century. Sci. Rep. 2018, 8, 16865. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilization strategies for improved nutrient uptake by plants. Boil. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef] [Green Version]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. A review. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Grzebisz, W.; Niewiadomska, A.; Przygocka-Cyna, K. Nitrogen Hotspots on the farm—A practice-oriented approach. Agronomy 2022, 12, 1305. [Google Scholar] [CrossRef]
- Berbell, J.; Martinez-Dalmau, J. A simple agro-economic model for optimal farm nitrogen application under yield uncertainty. Agronomy 2021, 11, 1107. [Google Scholar] [CrossRef]
- Córdova, C.; Barrera, J.A.; Magna, C. Spatial variation in nitrogen mineralization as a guide for variable application of nitrogen fertilizer to cereal crops. Nutr. Cycl. Agroecosyst. 2018, 110, 83–88. [Google Scholar] [CrossRef]
- Herath, A.; Ma, B.L.; Shang, J.; Liu, J.; Dong, T.; Jiao, X.; Kovacs, J.M.; Walters, D. On-farm spatial characterization of soil mineral nitrogen, crop growth, and yield of canola as affected by different rates of nitrogen application. Can. J. Soil Sci. 2018, 98, 1–14. [Google Scholar] [CrossRef]
- Stamatiadis, S.; Schepers, J.S.; Evangelou, E.; Tsadilas, C.; Glampedakis, M.; Dercas, N.; Spyropoulos, N.; Dalezios, N.R.; Eskridge, K. Variable-rate nitrogen fertilization of winter wheat under high spatial resolution. Precis. Agric. 2018, 19, 570–587. [Google Scholar] [CrossRef]
- Vizzari, M.; Santaga, F.; Benincasa, P. Sentinel 2-based nitrogen VRT fertilization in wheat: Comparison between traditional and simple precision practices. Agronomy 2019, 9, 278. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients (% or mg kg−1 of Dry Weight) | Maize | Sugar Beet | ||
|---|---|---|---|---|
| BBCH 17 | BBCH 17 | BBCH 41 | BBCH 33 | |
| Bergmann [28] | Schulte and Kelling [45] | Bergmann [28] | Barłóg [43] | |
| Nitrogen, N, % | 3.5–5.0 | 4.0–5.0 | 4.5–5.5 | 3.8–6.0 |
| Phosphorus, P, % | 0.35–0.6 | 0.4–0.6 | 0.3–0.6 | 0.27–0.46 |
| Potassium, K, % | 3.5–4.5 | 3.0–5.0 | 3.8–7.0 | 3.8–8.6 |
| Magnesium, Mg, % | 0.25–0.50 | 0.3–0.6 | 0.25–0.8 | 0.12–0.45 |
| Calcium, Ca, % | 0.3–1.0 | 0.51–1.6 | 0.6–1.5 | 0.28–0.85 |
| Zinc, Zn, mg kg−1 | 30–70 | 25–60 | 20–80 | 15–45 |
| Nutrients (% or mg kg−1 of Dry Weight) | Authors | |||
|---|---|---|---|---|
| Schulte and Kelling [45] | Jones et al. [46] | Campbell and Plank [41] | Potarzycki [33] | |
| Nitrogen, N, % | 3.0–4.0 | 2.6–3.6 | 2.8–4.0 | 2.1–3.33 |
| Phosphorus, P, % | 0.3–0.45 | 0.22–0.4 | 0.25–0.5 | 0.23–0.35 |
| Potassium, K, % | 2.0–3.0 | 1.8–4.5 | 1.8–3.0 | 1.9–2.5 |
| Magnesium, Mg, % | 0.2–0.8 | 0.43–1.0 | 0.25–0.8 | 0.41–0.67 |
| Calcium, Ca, % | 0.2–1.0 | 0.27–0.34 | 0.15–0.6 | 0.28–0.36 |
| Zinc, mg kg−1 | 20–70 | 19–75 | 20–70 | 40–70 1 |
| Nutrient | Ion | Dw, cm2 s−1 | Deff, cm2 s−1 |
|---|---|---|---|
| Nitrogen | NH4+ | 1.96 × 10−5 | 6.1 × 10−8 |
| NO3− | 1.90 × 10−5 | 2.7 × 10−6 | |
| Phosphorus | H2PO4− | 0.89 × 10−5 | 0.3–3.33 × 10−9 |
| Potassium | K+ | 2.00 × 10−5 | 1–28 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barłóg, P.; Grzebisz, W.; Łukowiak, R. Fertilizers and Fertilization Strategies Mitigating Soil Factors Constraining Efficiency of Nitrogen in Plant Production. Plants 2022, 11, 1855. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141855
Barłóg P, Grzebisz W, Łukowiak R. Fertilizers and Fertilization Strategies Mitigating Soil Factors Constraining Efficiency of Nitrogen in Plant Production. Plants. 2022; 11(14):1855. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141855
Chicago/Turabian StyleBarłóg, Przemysław, Witold Grzebisz, and Remigiusz Łukowiak. 2022. "Fertilizers and Fertilization Strategies Mitigating Soil Factors Constraining Efficiency of Nitrogen in Plant Production" Plants 11, no. 14: 1855. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141855