

Soil Contamination with Europium Induces Reduced Oxidative Damage in Hordeum vulgare Grown in a CO2-Enriched Environment
 , ,
, ,
Abstract
:1. Introduction
2. Results
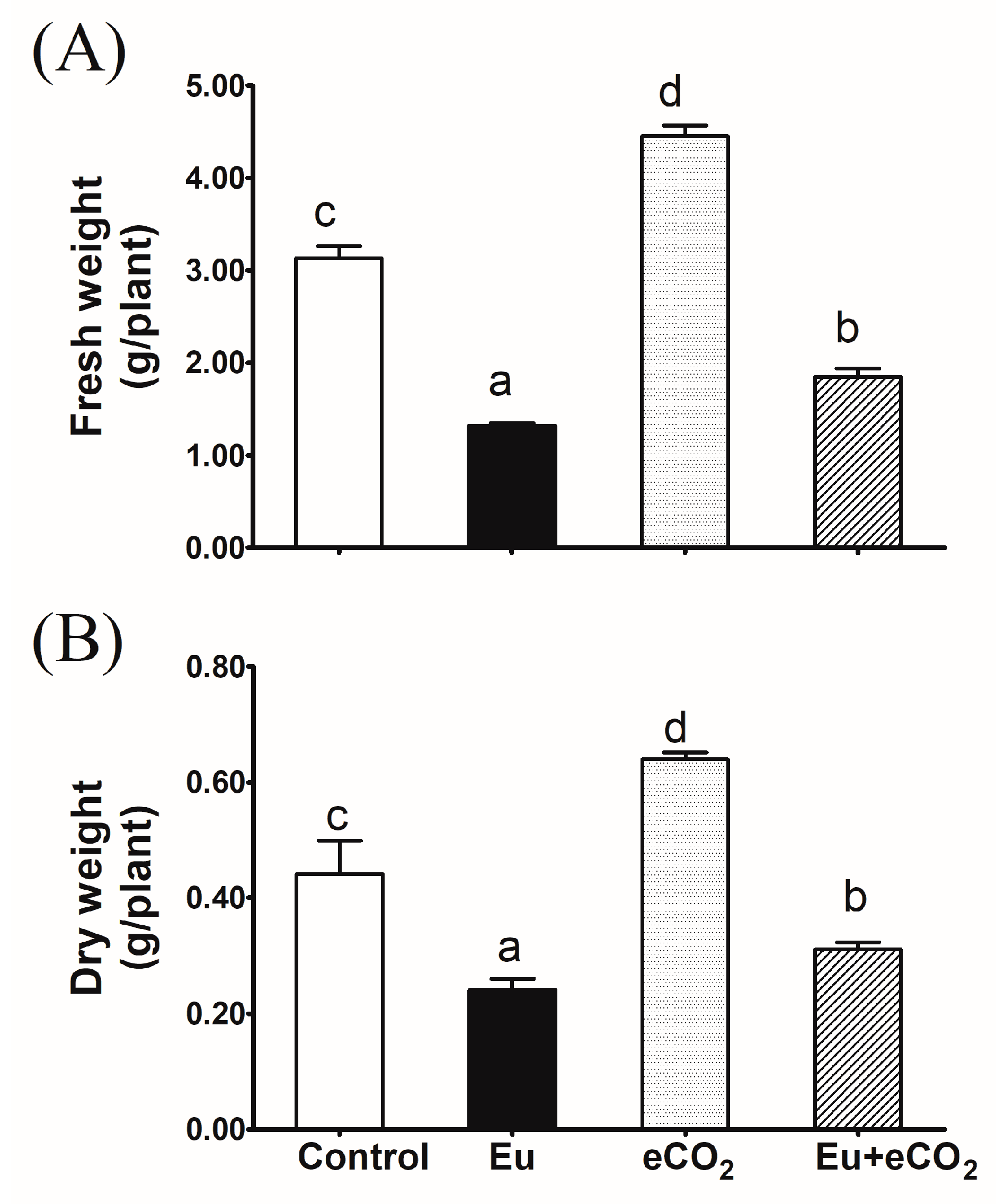
2.1. Biomass Production
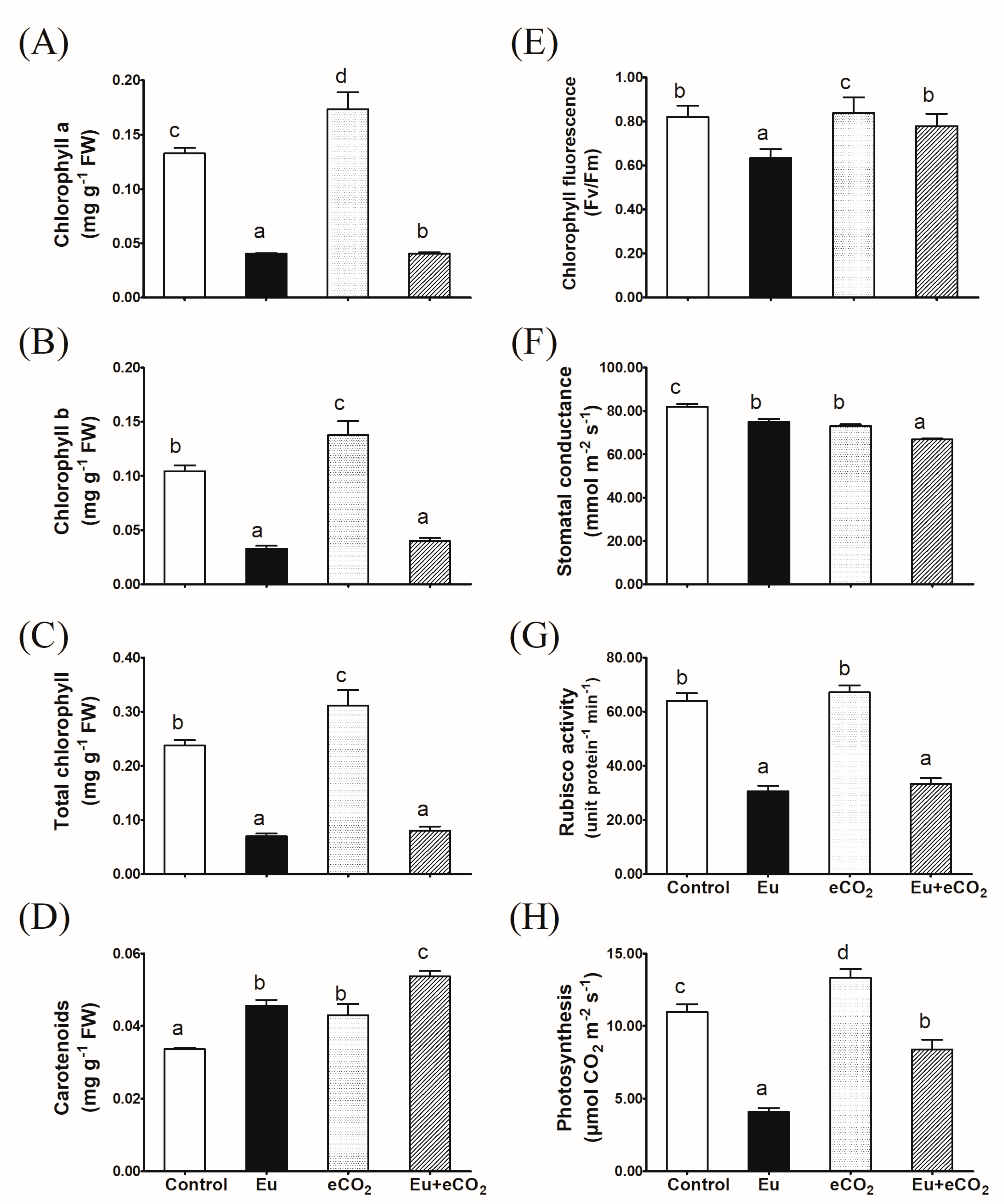
2.2. Photosynthesis-Related Parameters
2.3. Stress Markers, Antioxidant Molecules, and GST Activity
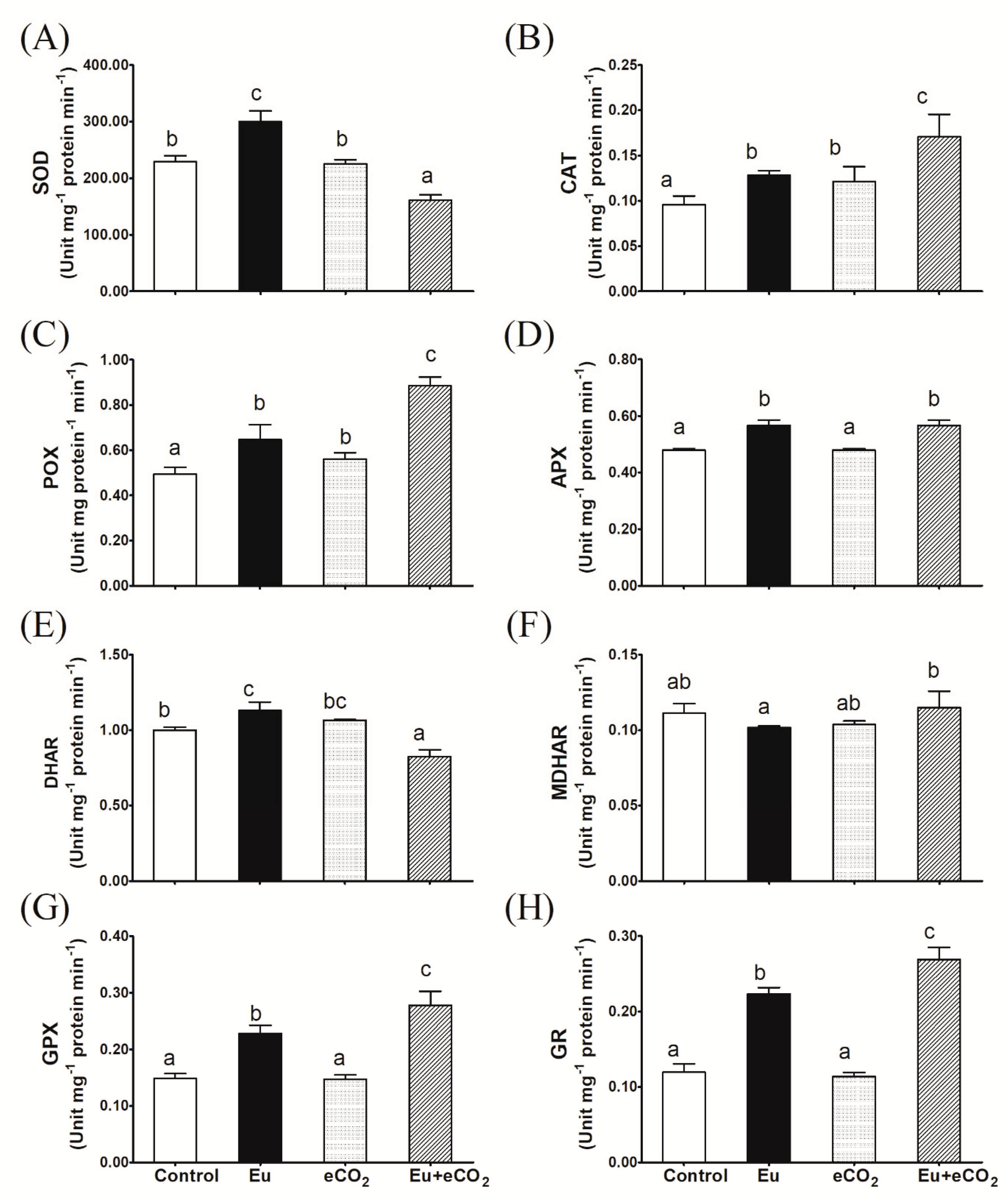
2.4. Antioxidant Enzyme Activity
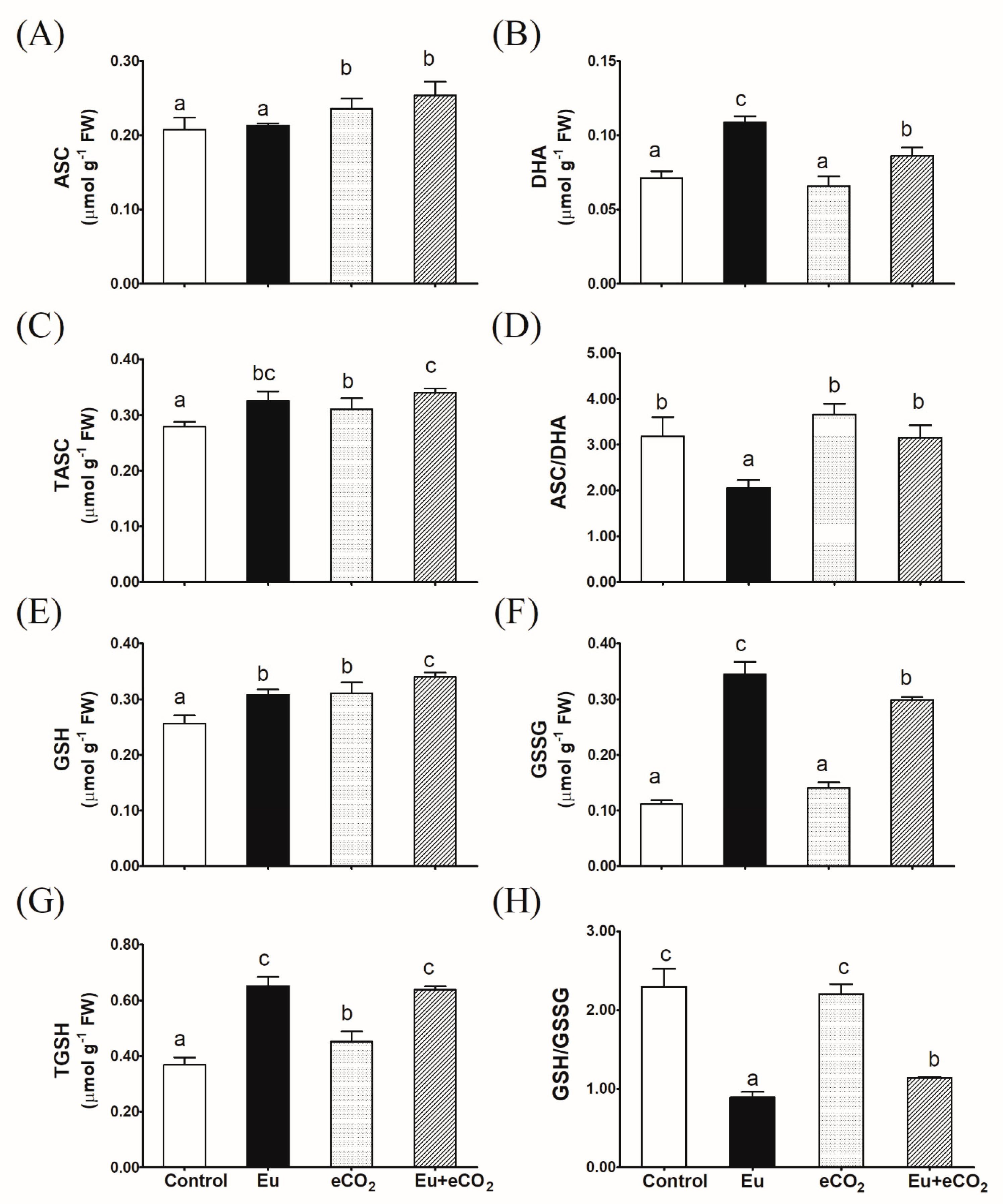
2.5. ASC—GSH Cycle Metabolites
3. Discussion
3.1. The Accumulation of Eu in Plant Shoots Slows Growth and Photosynthesis
3.2. Elevated CO2 Reduces the Accumulation of Eu and Mitigates its Impact on Growth and Photosynthesis
3.3. Eu Disrupts the Redox Homeostasis in Barley Plants but eCO2 Has an Antagonizing Action
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. Photosynthesis-Related Parameters
4.3. Determination of Eu Accumulation
4.4. Oxidative Stress Markers
4.5. Total Antioxidant Capacity and Antioxidant Metabolites
4.6. Antioxidant Enzymes and Glutathione-S-Transferase
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tyler, G. Rare Earth Elements in Soil and Plant Systems—A Review. Plant Soil 2004, 267, 191–206. [Google Scholar] [CrossRef]
- Pecharsky, V.K.; Gschneidner, K.A. Rare-Earth Element, Encyclopaedia Britannica. Encycl. Br. 2014, 10, 2017. [Google Scholar]
- Patil, A.S.; Patil, A.V.; Dighavkar, C.G.; Adole, V.A.; Tupe, U.J. Synthesis Techniques and Applications of Rare Earth Metal Oxides Semiconductors: A Review. Chem. Phys. Lett. 2022, 796, 139555. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, W.; Wang, Z.; Witkamp, G.J. Distributions of Rare Earths and Heavy Metals in Field-Grown Maize after Application of Rare Earth-Containing Fertilizer. Sci. Total Environ. 2002, 293, 97–105. [Google Scholar] [CrossRef]
- Golroudbary, S.R.; Makarava, I.; Kraslawski, A.; Repo, E. Global Environmental Cost of Using Rare Earth Elements in Green Energy Technologies. Sci. Total Environ. 2022, 832, 155022. [Google Scholar] [CrossRef]
- Sánchez-López, A.L.; Perfecto-Avalos, Y.; Sanchez-Martinez, A.; Ceballos-Sanchez, O.; Sepulveda-Villegas, M.; Rincón-Enríquez, G.; Rodríguez-González, V.; Garcia-Varela, R.; Lozano, L.M.; Navarro-López, D.E.; et al. Influence of Erbium Doping on Zinc Oxide Nanoparticles: Structural, Optical and Antimicrobial Activity. Appl. Surf. Sci. 2022, 575, 151764. [Google Scholar] [CrossRef]
- Moll, H.; Sachs, S.; Geipel, G. Plant Cell (Brassica Napus) Response to Europium(III) and Uranium(VI) Exposure. Environ. Sci. Pollut. Res. 2020, 27, 32048–32061. [Google Scholar] [CrossRef] [PubMed]
- Jessat, J.; Moll, H.; John, W.A.; Bilke, M.L.; Hübner, R.; Kretzschmar, J.; Steudtner, R.; Drobot, B.; Stumpf, T.; Sachs, S. A Comprehensive Study on the Interaction of Eu(III) and U(VI) with Plant Cells (Daucus Carota) in Suspension. J. Hazard. Mater. 2022, 439, 129520. [Google Scholar] [CrossRef]
- Liu, H.; Liu, H.; Yang, Z.; Wang, K. Bone Mineral Density in Population Long-Term Exposed to Rare Earth Elements from a Mining Area of China. Biol. Trace Elem. Res. 2021, 199, 453–464. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.J.; Liu, Y.; Liu, Z.; Zhang, A.N. Quantitative Contributions of the Major Sources of Heavy Metals in Soils to Ecosystem and Human Health Risks: A Case Study of Yulin, China. Ecotoxicol. Environ. Saf. 2018, 164, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Aranjuelo, I.; Doustaly, F.; Cela, J.; Porcel, R.; Müller, M.; Aroca, R.; Munné-Bosch, S.; Bourguignon, J. Glutathione and Transpiration as Key Factors Conditioning Oxidative Stress in Arabidopsis Thaliana Exposed to Uranium. Planta 2014, 239, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Serre, N.B.C.; Alban, C.; Bourguignon, J.; Ravanel, S. Uncovering the Physiological and Cellular Effects of Uranium on the Root System of Arabidopsis Thaliana. Environ. Exp. Bot. 2019, 157, 121–130. [Google Scholar] [CrossRef]
- Ramos, S.J.; Dinali, G.S.; Oliveira, C.; Martins, G.C.; Moreira, C.G.; Siqueira, J.O.; Guilherme, L.R.G. Rare Earth Elements in the Soil Environment. Curr. Pollut. Rep. 2016, 2, 28–50. [Google Scholar] [CrossRef]
- Tian, H.E.; Gao, Y.S.; Li, F.M.; Zeng, F. Effects of Europium Ions (Eu3+) on the Distribution and Related Biological Activities of Elements in Lathyrus sativus L. Roots. Biol. Trace Elem. Res. 2003, 93, 257–269. [Google Scholar] [CrossRef]
- Zenk, M.H. Heavy Metal Detoxification in Higher Plants-a Review. Gene 1996, 179, 21–30. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Sheteiwy, M.S.; Saleh, A.M.; Mohammed, A.E.; Alotaibi, M.O.; Beemster, G.T.S.; Madany, M.M.Y.; van Dijk, J.R. Elevated CO2 Differentially Mitigates Chromium (VI) Toxicity in Two Rice Cultivars by Modulating Mineral Homeostasis and Improving Redox Status. Chemosphere 2022, 307, 135880. [Google Scholar] [CrossRef]
- Saleh, A.M.; Hassan, Y.M.; Habeeb, T.H.; Alkhalaf, A.A.; Hozzein, W.N.; Selim, S.; AbdElgawad, H. Interactive Effects of Mercuric Oxide Nanoparticles and Future Climate CO2 on Maize Plant. J. Hazard. Mater. 2021, 401, 123849. [Google Scholar] [CrossRef]
- Mittova, V.; Guy, M.; Tal, M.; Volokita, M. Salinity Up-Regulates the Antioxidative System in Root Mitochondria and Peroxisomes of the Wild Salt-Tolerant Tomato Species Lycopersicon Pennellii. J. Exp. Bot. 2004, 55, 1105–1113. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kral, A. Ecotype-Specific Pathways of Reactive Oxygen Species Deactivation in Facultative Metallophyte Silene Vulgaris (Moench) Garcke Treated with Heavy Metals. Antioxidants 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- NOAA Carbon Dioxide Now More than 50% Higher than Pre-Industrial Levels. 2022. Available online: https://www.noaa.gov/news-release/carbon-dioxide-now-more-than-50-higher-than-pre-industrial-levels (accessed on 28 July 2023).
- Murray, V.; Ebi, K.L. IPCC Special Report on Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation (SREX). J. Epidemiol. Community Health 2012, 66, 759–760. [Google Scholar] [CrossRef] [PubMed]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of IPCC the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA; 1535p.
- Al Jaouni, S.; Saleh, A.M.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; AbdElgawad, H. Elevated CO2 Induces a Global Metabolic Change in Basil (Ocimum basilicum L.) and Peppermint (Mentha piperita L.) and Improves Their Biological Activity. J. Plant Physiol. 2018, 224–225, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Selim, S.; Al Jaouni, S.; AbdElgawad, H. CO2 Enrichment Can Enhance the Nutritional and Health Benefits of Parsley (Petroselinum crispum L.) and Dill (Anethum graveolens L.). Food Chem. 2018, 269, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Zinta, G.; AbdElgawad, H.; Domagalska, M.A.; Vergauwen, L.; Knapen, D.; Nijs, I.; Janssens, I.A.; Beemster, G.T.S.; Asard, H. Physiological, Biochemical, and Genome-Wide Transcriptional Analysis Reveals That Elevated CO2 Mitigates the Impact of Combined Heat Wave and Drought Stress in Arabidopsis Thaliana at Multiple Organizational Levels. Glob. Chang. Biol. 2014, 20, 3670–3685. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Beemster, G.T.S.; Janssens, I.A.; Asard, H. Future Climate CO2 Levels Mitigate Stress Impact on Plants: Increased Defense or Decreased Challenge? Front. Plant Sci. 2016, 7, 556. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Mohammed, A.E.; van Dijk, J.R.; Beemster, G.T.S.; Alotaibi, M.O.; Saleh, A.M. The Impact of Chromium Toxicity on the Yield and Quality of Rice Grains Produced under Ambient and Elevated Levels of CO 2. Front. Plant Sci. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- AbdElgawad, H.; El-Sawah, A.M.; Mohammed, A.E.; Alotaibi, M.O.; Yehia, R.S.; Selim, S.; Saleh, A.M.; Beemster, G.T.S.; Sheteiwy, M.S. Increasing Atmospheric CO2 Differentially Supports Arsenite Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Wheat and Soybean Plants. Chemosphere 2022, 296, 134044. [Google Scholar] [CrossRef]
- Saleh, A.M.; Hassan, Y.M.; Selim, S.; AbdElgawad, H. NiO-Nanoparticles Induce Reduced Phytotoxic Hazards in Wheat (Triticum aestivum L.) Grown under Future Climate CO2. Chemosphere 2019, 220, 1047–1057. [Google Scholar] [CrossRef]
- Singh, L.; Park, R.F.; Dracatos, P.; Ziems, L.; Singh, D. Understanding the Expression and Interaction of Rph Genes Conferring Seedling and Adult Plant Resistance to Puccinia Hordei in Barley. Can. J. Plant Pathol. 2021, 43, S218–S226. [Google Scholar] [CrossRef]
- Vaezi, B.; Pour-Aboughadareh, A.; Mohammadi, R.; Armion, M.; Mehraban, A.; Hossein-Pour, T.; Dorii, M. GGE Biplot and AMMI Analysis of Barley Yield Performance in Iran. Cereal Res. Commun. 2017, 45, 500–511. [Google Scholar] [CrossRef]
- OECD Consensus Document on Compositional Considerations for New Varieties of Cotton (Gossypium Hirsutum and Gossypium Barbadense): Key Food and Feed Nutrients and Anti-Nutrients. Ser. Saf. Nov. Foods Feed. 2004, 32.
- FAO. World Food and Agriculture Statistical Yearbook 2021; FAO: Rome, Italy, 2021. [Google Scholar]
- Rahmani, A.; Asghari, A.; Jafari, H.; Sofalian, O. QTL Mapping for Physiological Traits Affecting Lead Tolerance in the Hordeum Vulgare L. Environ. Stress. Crop Sci. 2021, 14, 849–860. [Google Scholar]
- Gao, Y.; Zeng, F.; Yi, A.; Ping, S.; Jing, L. Research of the Entry of Rare Earth Elements Eu 3+ and La 3+ into Plant Cell. Biol. Trace Elem. Res. 2003, 91, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Shan, X.; Huai, Q.; Wen, B.; Zhu, X. Uptake and Distribution of Rare Earth Elements in Rice Seeds Cultured in Fertilizer Solution of Rare Earth Elements. Chemosphere 2001, 43, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Tagami, K.; Uchida, S. Transfer of REEs from Nutrient Solution to Radish through Fine Roots and Their Distribution in the Plant. J. Alloys Compd. 2006, 408, 409–412. [Google Scholar] [CrossRef]
- Stadler, J.; Vogel, M.; Steudtner, R.; Drobot, B.; Kogiomtzidis, A.L.; Weiss, M.; Walther, C. The Chemical Journey of Europium(III) through Winter Rye (Secale cereale L.)—Understanding through Mass Spectrometry and Chemical Microscopy. Chemosphere 2023, 313, 137252. [Google Scholar] [CrossRef]
- Malik, B.; Pirzadah, T.B.; Tahir, I.; Rehman, R.U. Growth and Physiological Responses in Chicory towards Mercury Induced in Vitro Oxidative Stress. Plant Physiol. Rep. 2019, 24, 236–248. [Google Scholar] [CrossRef]
- Chaudhary, S.; Sharma, P.; Kumar, S.; Alex, S.A.; Kumar, R.; Mehta, S.K.; Mukherjee, A.; Umar, A. A Comparative Multi-Assay Approach to Study the Toxicity Behaviour of Eu2O3 Nanoparticles. J. Mol. Liq. 2018, 269, 783–795. [Google Scholar] [CrossRef]
- Kupcsik, L. Estimation of Cell Number Based on Metabolic Activity: The MTT Reduction Assay. Mamm. Cell Viability Methods Protoc. 2011, 740, 13–19. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Fernandez, V.; Barnaby, J.Y.; Tomecek, M.; Codling, E.E.; Ziska, L.H. Elevated CO2 May Reduce Arsenic Accumulation in Diverse Ecotypes of Arabidopsis Thaliana. J. Plant Nutr. 2018, 41, 645–653. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Hassan, Y.M.; Alotaibi, M.O.; Mohammed, A.E.; Saleh, A.M. C3 and C4 Plant Systems Respond Differently to the Concurrent Challenges of Mercuric Oxide Nanoparticles and Future Climate CO2. Sci. Total Environ. 2020, 749, 142356. [Google Scholar] [CrossRef]
- Habeeb, T.H.; Abdel-mawgoud, M.; Yehia, R.S.; Khalil, A.M.A.; Saleh, A.M.; AbdElgawad, H. Interactive Impact of Arbuscular Mycorrhizal Fungi and Elevated CO2 on Growth and Functional Food Value of Thymus Vulgare. J. Fungi 2020, 6, 168. [Google Scholar] [CrossRef]
- Saleh, A.M.; Kebeish, R. Coumarin Impairs Redox Homeostasis in Wheat Aleurone Layers. J. Plant Res. 2018, 131, 157–163. [Google Scholar] [CrossRef]
- Lejay, L.; Wirth, J.; Pervent, M.; Cross, J.M.-F.; Tillard, P.; Gojon, A. Oxidative Pentose Phosphate Pathway-Dependent Sugar Sensing as a Mechanism for Regulation of Root Ion Transporters by Photosynthesis. Plant Physiol. 2008, 146, 2036–2053. [Google Scholar] [CrossRef]
- Quan, L.-J.; Zhang, B.; Shi, W.-W.; Li, H.-Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Moon, J.-C.; Kim, C.; Manoharan, K.; Kim, W. Reactive Oxygen Species in Plants: Their Generation, Signal Transduction, and Scavenging Mechanisms. Aust. J. Crop Sci. 2011, 5, 709–725. [Google Scholar]
- Foyer, C.H.; Noctor, G. Tansley Review No. 112 Oxygen Processing in Photosynthesis: Regulation and Signalling. New Phytol. 2000, 146, 359–388. [Google Scholar] [CrossRef]
- Ghnaya, A.B.; Charles, G.; Hourmant, A.; Hamida, J.B.; Branchard, M. Physiological Behaviour of Four Rapeseed Cultivar (Brassica napus L.) Submitted to Metal Stress. C. R. Biol. 2009, 332, 363–370. [Google Scholar] [CrossRef]
- Saleh, A.M.; Madany, M. Investigation of the Allelopathic Potential of Alhagi Graecorum Boiss. Asian J. Agric. Res. 2014, 8, 42–50. [Google Scholar] [CrossRef]
- AbdElgawad, H.; de Soua, A.; Alotaibi, M.O.; Mohammed, A.E.; Schoenaers, S.; Selim, S.; Saleh, A.M. The Differential Tolerance of C3 and C4 Cereals to Aluminum Toxicity Is Faded under Future CO2 Climate. Plant Physiol. Biochem. 2021, 169, 249–258. [Google Scholar] [CrossRef]
- Yadav, S.K. Heavy Metals Toxicity in Plants: An Overview on the Role of Glutathione and Phytochelatins in Heavy Metal Stress Tolerance of Plants. South African J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef]
- Abdelgawad, H.; Farfan-vignolo, E.R.; De Vos, D.; Asard, H. Elevated CO2 Mitigates Drought and Temperature-Induced Oxidative Stress Differently in Grasses and Legumes. Plant Sci. 2015, 231, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta (BBA)-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Agusa, T.; Kunito, T.; Yasunaga, G.; Iwata, H.; Subramanian, A.; Ismail, A.; Tanabe, S. Concentrations of Trace Elements in Marine Fish and Its Risk Assessment in Malaysia. Mar. Pollut. Bull. 2005, 51, 896–911. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Jiang, Z.-Y.; Woollard, A.C.S.; Wolff, S.P. Hydrogen Peroxide Production during Experimental Protein Glycation. Febs Lett. 1990, 268, 69–71. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. [2] Ferric Reducing/Antioxidant Power Assay: Direct Measure of Total Antioxidant Activity of Biological Fluids and Modified Version for Simultaneous Measurement of Total Antioxidant Power and Ascorbic Acid Concentration. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 15–27. [Google Scholar]
- Potters, G.; Horemans, N.; Bellone, S.; Caubergs, R.J.; Trost, P.; Guisez, Y.; Asard, H. Dehydroascorbate Influences the Plant Cell Cycle through a Glutathione-Independent Reduction Mechanism. Plant Physiol. 2004, 134, 1479–1487. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.A.; Barrow, C.J. A Simple 96-Well Microplate Method for Estimation of Total Polyphenol Content in Seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colorimetric Methods. J. Food Drug Anal. 2002, 10. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.L.; Reid, D.M. Leaf Senescence and Lipid Peroxidation: Effects of Some Phytohormones, and Scavengers of Free Radicals and Singlet Oxygen. Physiol. Plant. 1982, 56, 453–457. [Google Scholar] [CrossRef]
- Kumar, K.B.; Khan, P.A. Peroxidase and Polyphenol Oxidase in Excised Ragi (Eleusine Corocana Cv PR 202) Leaves during Senescence. Indian J. Exp. Biol. 1982, 20, 412–416. [Google Scholar]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. ISBN 0076-6879. [Google Scholar]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate Quantification of Enzymes of the Plant Ascorbate–Glutathione Cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Diopan, V.; Shestivska, V.; Adam, V.; Macek, T.; Mackova, M.; Havel, L.; Kizek, R. Determination of Content of Metallothionein and Low Molecular Mass Stress Peptides in Transgenic Tobacco Plants. Plant Cell Tissue Organ Cult. 2008, 94, 291–298. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Eu | eCO2 | Eu + eCO2 |
|---|---|---|---|---|
|
Eu content (mg g−1 DW) | ND | 38.69 ± 1.25 b | ND | 21.65 ± 0.52 a |
|
H2O2 (nmol g−1 FW) | 562.49 ± 13.63 b | 964.04 ± 45.26 d | 495.1 ± 24.88 a | 708.33 ± 13.6 c |
|
MDA (nmol g−1 FW) | 2.48 ± 0.06 a | 4.31 ± 0.24 c | 2.33 ± 0.23 a | 3.61 ± 0.35 b |
|
FRAP (mmol trolox g−1 FW) | 36.77 ± 2.87 a | 46.07 ± 1.54 b | 43.55 ± 0.61 b | 54.71 ± 2.28 c |
|
Polyphenol (mg gallic acid g−1 FW) | 15.23 ± 0.4 a | 20.53 ± 1.07 c | 18.94 ± 0.46 b | 25.32 ± 1.26 d |
|
Flavonoids (mg quercetin g−1 FW) | 35.02 ± 1.37 c | 26.07 ± 1.07 a | 37.47 ± 1.34 d | 29.27 ± 0.81 b |
|
GST (unit mg−1 protein min−1) | 0.19 ± 0.01 a | 0.18 ± 0.01 a | 0.19 ± 0 a | 0.27 ± 0.01 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amer, H.E.A.; AbdElgawad, H.; Madany, M.M.Y.; Khalil, A.M.A.; Saleh, A.M. Soil Contamination with Europium Induces Reduced Oxidative Damage in Hordeum vulgare Grown in a CO2-Enriched Environment. Plants 2023, 12, 3159. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12173159
Amer HEA, AbdElgawad H, Madany MMY, Khalil AMA, Saleh AM. Soil Contamination with Europium Induces Reduced Oxidative Damage in Hordeum vulgare Grown in a CO2-Enriched Environment. Plants. 2023; 12(17):3159. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12173159
Chicago/Turabian StyleAmer, Hanaa E. A., Hamada AbdElgawad, Mahmoud M. Y. Madany, Ahmed M. A. Khalil, and Ahmed M. Saleh. 2023. "Soil Contamination with Europium Induces Reduced Oxidative Damage in Hordeum vulgare Grown in a CO2-Enriched Environment" Plants 12, no. 17: 3159. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12173159