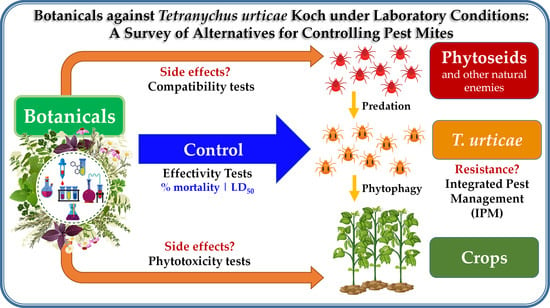
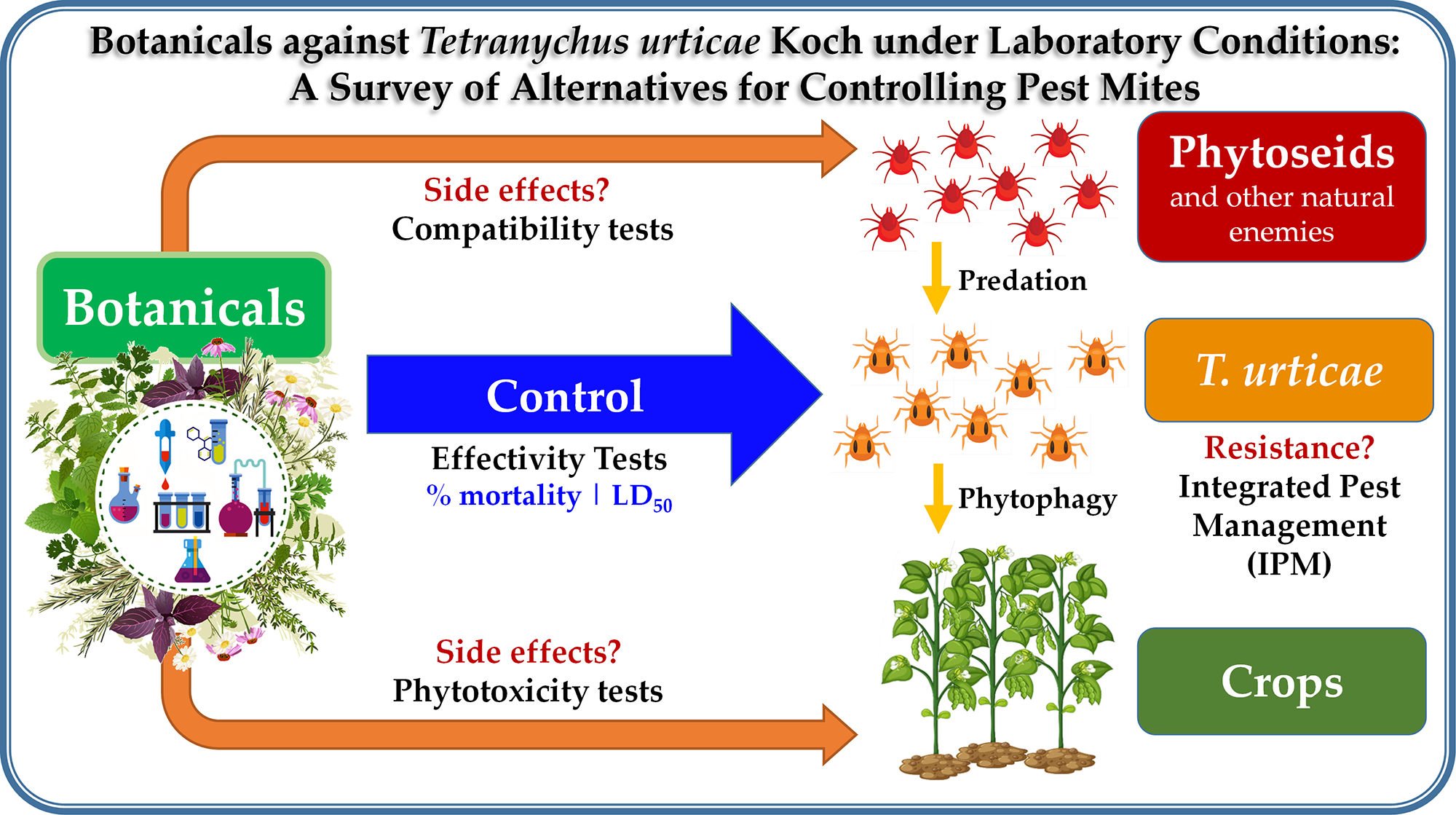
Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites



Abstract
:
1. Introduction
2. Characteristics of T. urticae
2.1. The Biology of T. urticae Koch
2.2. Characteristics of Resistance of T. urticae Koch to Acaricides
Function of Detoxifying Enzymes
2.3. Relationship Between Resistance and the Host Plant
3. Control Strategies for T. urticae Koch
3.1. Other Methods for T. urticae Koch Control
3.2. The Use of Plant Extracts for the Control of T. urticae Koch in The Field
3.2.1. Methods for the Evaluation of Extracts under Laboratory Conditions
3.2.2. Bioassays
Slide Dip Methods
Petri Dish Methods
Leaf Disc Methods
Leaf Absorption Method
Whole Plant Direct Method
Filter Paper Difussion Methods (Fumigant Bioassays)
3.3. Studies Carried Out for the Control of T. urticae Koch from Plant Extracts Grouped by Plant Families
3.3.1. Family Amaranthaceae
3.3.2. Family Amaryllidaceae
3.3.3. Family Annonaceae
3.3.4. Family Apiaceae
3.3.5. Family Asteraceae
3.3.6. Family Boraginaceae
3.3.7. Family Burseraceae
3.3.8. Family Cannabaceae
3.3.9. Family Caryophyllaceae
3.3.10. Family Combretaceae
3.3.11. Family Convolvulaceae
3.3.12. Family Cupressaceae
3.3.13. Family Euphorbiaceae
3.3.14. Family Fabaceae
3.3.15. Family Gramineae (Poaceae)
3.3.16. Family Lamiaceae
3.3.17. Family Meliaceae
3.3.18. Family Myrtaceae
3.3.19. Piperaceae Family
3.3.20. Family Ranunculaceae
3.3.21. Family Rutaceae
3.3.22. Family Santalaceae
3.3.23. Family Scrophulariaceae
3.3.24. Family Simarubaceae
3.3.25. Family Solanaceae
3.3.26. Verbenaceae Family
3.4. Additional Studies with Isolated Compounds Obtained after Plant Extract Fractionation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Family | Plant Species | Source | Concentration | Bioassaya | T. urticae Koch Stage | Effect on T. urticae Koch | Identified Compounds | Ref. |
|---|---|---|---|---|---|---|---|---|
| Amaranthaceae | Amaranthus viridis L. | Whole plant extract | 5000 ppm | G | Adults | Mortality between 40 and 60% | - | [108] |
| Amaranthaceae | Amaranthus viridis L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [108] |
| Amaranthaceae | Blepharis linariifolia Pers. | Whole plant extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [108] |
| Amaranthaceae | Blepharis linariifolia Pers. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [108] |
| Amaranthaceae | Blepharis sp. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [108] |
| Amaranthaceae | Blepharis sp. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [108] |
| Amaranthaceae | Celosia Trygina L. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [108] |
| Amaranthaceae | Celosia Trygina L. | Whole plant extract | 2500 ppm | G | Adults | More than 80% of mortality | - | [108] |
| Amaranthaceae | Chenopodium ambrosioides Mosyakin et Clemants | Emulsifiable Concentrate | 0.50% | A,C | Adults and eggs | 94.7% of mortality | - | [41] |
| Amaranthaceae | Chenopodium quinoa Willd. | Seeds extract | 6–9% w/v [1.24% w/v (LD50)] | E,F | Adult females and nymphs | Mortalities ranged from 30% to 99% | - | [89] |
| Amaranthaceae | Kochia scoparia (L.) Schrad. | - | 98.13% (chloroform extraction) | A,E,H | Adult females | 92.58% of mortality | - | [52] |
| Amaryllidaceae | Allium cepa L. | Essential oil | - | D | Larvae and adults. | Mortalities of 65% (larvae) and 67% (adults) | - | [88] |
| Amaryllidaceae | Allium cepa L. | Peel fruit extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Amaryllidaceae | Allium galanthum Kar. & Kir. | Whole plant extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Amaryllidaceae | Allium obliquum L. | Whole plant extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Amaryllidaceae | Allium sativum L. | - | 7.2 g/L | A,G | Adult females | LD50 | - | [78] |
| Amaryllidaceae | Allium sativum L. | Bulb extract | 7.49 and 13.5 mg/L | E,F | Adult females | LD50 and LD90 (respectively) | - | [77] |
| Amaryllidaceae | Allium sativum L. | Essential oil | - | D | Larvae and adults | Mortalities of 86% (larvae) and 61% (adults) | - | [88] |
| Amaryllidaceae | Pancratium maritimum L. | Alkaloidal ethanolic extract and bulb essential oil | 0.2%. 0.36% and 1.5% respectively | - | LD50 | - | [76] | |
| Amaryllidaceae | Ungernia severtzovii Regel | Root extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Anacardiaceae | Cotinus coggygria Scop. | Essential oil | - | D | Larvae and adults | Mortalities of 58% (larvae) and 58% (adults) | - | [88] |
| Anacardiaceae | Cotinus coggygria Scop. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Anacardiaceae | Pistacia lentiscus L. | Essential oil | - | D | Larvae and adults | Mortalities of 22% (larvae) and 23% (adults) | - | [88] |
| Annonaceae | Annona glabra L. | Seed extract | 1000 ppm | D,G | Eggs | No effects | - | [80] |
| Annonaceae | Cananga odorata (Lam.) Hook.F. & Thomson | Essential oil | 0.1% | G | Adult females | 24.2% of mortality | - | [112] |
| Annonaceae | Xilopia sericea A.St.-Hill. | Leaves and fruits essential oils | 4.08 µL/L | C | Adult females | LD50 | α-pinene (0.41% leaves, 17.18% fruits), β-pinene (45.59% fruits), cubenol (57.43% leaves), myrcene (9.13% fruits), between others | [44] |
| Apiaceae | Ammi visnaga | Seed extract | 17 µg/cm2 | D,I | Eggs | LD50 | Kheline and visnagine | [65] |
| Apiaceae | Carum carvi L. | Essential oil | 19 × 10−3 µL/mL of air. | J | Adults | 100% of mortality | - | [42] |
| Apiaceae | Carum carvi L. | Essential oil | 22.4 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Apiaceae | Carum carvi L. | Essential oils mixed with Fatty acid potassium salts | 570 ppm of essential oil and 2478 ppm of potassium salts | E | Adults | 83.4% of mortality | - | [83] |
| Apiaceae | Conium maculatum L. | Flowers and leaves extract | 10–50% | A,B | Adult females | Mortalities of 95.18% and 81.11%, respectively | - | [60] |
| Apiaceae | Coriandrum sativum L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 92% of mortality | - | [42] |
| Apiaceae | Cuminum cyminum L. | Essential oil from seeds | 3.74 µL/L air | C,J | Adult females | LD50 | α-pinene (29.1%), limonene (22%), 1,8-cineole (17.9%) | [85] |
| Apiaceae | Daucus carota L. | Essential oil | - | D | Larvae and adults | Mortalities of 5% (larvae) and 3% (adults) | - | [88] |
| Apiaceae | Deverra scoparia Coss. & Durieu | Essential oil | 1.79 and 3.2 mg/L | E | Young females | LD50 and LD90, respectively | α-pinene, ∆3-carene and terpinen-4-ol | [43] |
| Apiaceae | Deverra scoparia Coss. & Durieu | Essential oil | - | D | larvae and adults | Mortalities of 98% (larvae) and 97% (adults) | - | [88] |
| Apiaceae | Ferula gumosa Boiss. | Essential oil | 6.98 and 6.52 μL/L air | C,J | Eggs and adults, respectively | LD50 | β-pinene (50.1%), α-pinene (14.9%), δ-3-carene (6.7%) | [86] |
| Apiaceae | Foeniculum vulgare Mill. | Seed essential oil vapors | 5.75 µL/L (females), 1.17 µL/L (eggs) | J | Eggs and adults | LD50 | - | [84] |
| Apiaceae | Foeniculum vulgare Mill. | Essential oil | 1.17% | E | Adults | LD50 | - | [83] |
| Apiaceae | Heracleum persicum Desf. Ex. Fisch. | Fruit essential oils vapors | 3.15 µL/L (females)-1.53 µL/L (eggs) | J | Eggs and adults | LD50 | - | [84] |
| Apiaceae | Heracleum persicum Desf. Ex. Fisch. | Essential oil | 1.53% | E | Adults | LD50 | - | [83] |
| Apiaceae | Smyrnium olusatrum L. | Inflorescence extract | 1.9 and 42.7 µg/mL, respectively for isolated compounds | D | Adult females | LD50 (chronic toxicity after 5 days) | Isolation of isofuranodiene and germacrone, separately evaluated | [87] |
| Apocynaceae | Vinca erecta Regel & Schmalh | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Apocynaceae | Vinca minor L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Araceae | Arum korolkovii Regel | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asclepiadaceae | Calotropis gigantea W.T. Aiton | Leaf extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Asclepiadaceae | Calotropis gigantea W.T. Aiton | Leaf extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Asteraceae | Achillea mellifolium L. | Essential oil from aerial part | 1.208% v/v (leaf dipping) and 1.801 µL/L air (fumigation) | G,J | Adult females | LD50 | Piperitone (12.8%), p-cymene (10.6%) | [64] |
| Asteraceae | Achillea millefolium L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Acroptilon repens (L.) DC. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Ajania fastigiata (C. Winkler) Poljakov | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Anaphalis rosea-alba Krasch. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Anthemis nobilis L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 69% of mortality | - | [42] |
| Asteraceae | Anthemis vulgaris L. | Flower extract | 7–50% | A,B | Adult females | Mortalities of 92.37% and 92.34%, respectively | - | [60] |
| Asteraceae | Anthemis vulgaris L. | Leaf extract | 13–50% | A,B | Adult females | Mortalities of 82.33% and 76.63%, respectively | - | [60] |
| Asteraceae | Artemisia absinthium L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 97% of mortality | - | [42] |
| Asteraceae | Artemisia absinthium L. | Essential oil | 0.043 mg/cm2 | A | - | LD50 | - | [45] |
| Asteraceae | Artemisia absinthium L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Artemisia aschurbajewii C. Winkl. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Artemisia cinae O. Berg & C.F. Schmidt ex. Pljakov | Leaf extract | 1326.53 ppm | A,B | Adult females | LD50 | - | [60] |
| Asteraceae | Artemisia compacta Fisch. Ex. Besser | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Artemisia dracunculus L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Artemisia judaica L. | Leaves (acetone extract) | 0.56 μg/mL | C,G | Adult females | LD50 | - | [39] |
| Asteraceae | Artemisia panciflora | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Artemisia vulgaris L. | Leaf extract | 15-50% | A,B | Adult females | Mortalities of 54.13% and 75.12%, respectively | - | [60] |
| Asteraceae | Artemisia vulgaris L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Calendula officinalis L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Chamomilla recutita L. | Essential oil | 0.65–1.17% | C | Adults ggs | LD50 | α-basabolol oxide (35.25%), Trans β-farersene (7.76%) | [89] |
| Asteraceae | Chrisanthemum coronarium L. | Essential oil | - | D | Larvae and adults | Mortalities of 88% (larvae) and 93% (adults) | - | [88] |
| Asteraceae | Handelia trichopylla Heimerl | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20%. | - | [99] |
| Asteraceae | Hertia cheirifolia (L.) Kuntze | Essential oil | 3.43 mg/L | E | Adult females | LD50 and side-effect over fecundity | [46] | |
| Asteraceae | Hertia cheirifolia (L.) Kuntze | Essential oil | - | D | Larvae and adults | Mortalities of 81% (larvae) and 89% (adults) | - | [88] |
| Asteraceae | Hieracium dschirgalanicum E. Nikit. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Inula helenium L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Jurinea capussi Franch. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Lamyropappus schakaptaricus Knorr & Tamamsch. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Matricaria chamomilla L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Matricaria matricarioides (Less.) Porter | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Matricaria recutita L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Pseudoglossanthis litwinowii (Tzvel.) R. Kam. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Pyrethrum alatavicum O. & B. Fedtsch. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Pyrethrum branchanthemoides R. Kam. & Lazkov | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Pyrethrum cinerariifolium Trev. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Pyrethrum sovetkinae Kovalevsk | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Pyrethrum sussamyrense Lazkov | Root extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Santolina africana Jord. & Fourr. | Essential oil | 2.35 mg/L | E | Adult females | LD50 and side-effect over fecundity | Terpinen-4-ol (54.96%) | [46] |
| Asteraceae | Santolina africana Jord. & Fourr. | Essential oil | - | D | Larvae and adults | Mortalities of 77% (larvae) and 68% (adults) | - | [88] |
| Asteraceae | Senecio saposhnikovii Krasch et. Schipcz. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Seriphidium herba-album (Asso) Sojak | Essential oil | - | D | Larvae and adults | Mortalities of 54% (larvae) and 37% (adults) | - | [88] |
| Asteraceae | Tagetes minuta L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Tanacetopsis ferganensis Kovalevsk | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Tanacetopsis setacea Kovalevsk | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Tanacetopsis submarginata Kovalevsk | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Tanacetum boreale Fisch. Ex. DC. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Asteraceae | Tanacetum pseudoachillea C. Winkl. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Asteraceae | Tanacetum vulgare L. | Essential oil | 4% | A | - | 75.6% of mortality | - | [45] |
| Asteraceae | Thitonia diversifolia Hemsl. | Methanolic extract | 150 µg/cm3 | D | Adult females | Mortality less than 50% | - | [92] |
| Asteraceae | Tripleurospermum inodorum Sch. Bip. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Asteraceae | Xanthium strumarium L. | Fruit extract | 9-50% | A,B | Adult females | Mortalities of 68.24% and 85.88%, respectively | - | [60] |
| Asteraceae | Xanthium strumarium L. | Leaf extract | 11-50% | A,B | Adult females | Mortalities of 52.48% and 79.85%, respectively | - | [60] |
| Berveridaceae | Berberis iliensis Popov | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Bignonianceae | Jacaranda obtusifolia Bonpl. | Leaf extract | 0.06% | C,G | Adult females | Mortality of 64.4% | [124] | |
| Boraginaceae | Echium vulgare L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Boraginaceae | Onosma visianii Clem. | Root extract | 2.6 µg/mL | D | Adult females | LD50 (chronic toxicity after 5 days) | Shikonin derivatives (naphthoquinones), i.e., isobutylshikonin and isovalerylshikonin | [93] |
| Brassicaceae | Armoracia rusticana G. Gaertn., B. Mey. & Scherb. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Brassicaceae | Barbarea vulgaris W.T. Aiton | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Brassicaceae | Capsella bursa-pastoris L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Brassicaceae | Cardaria repens Schrenk | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Brassicaceae | Lepidium latifolium L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Burseraceae | Boswellia carterii Birdw. | Essential oil | 0.1% | G | Adult females | 24.8% of mortality | - | [112] |
| Burseraceae | Commiphora myrrha (Nees) Engl. | Essential oil | 0.1% | G | Adult females | 22.8% of mortality | - | [112] |
| Burseraceae | Protium bahianum Daly | Fresh and old resin essential oils | - | J | Adult females | Mortalities of 79.6% (fresh resin) and 59% (old resin) | - | [97] |
| Caesalpiniaceae | Cassia mimosoides L. | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Caesalpiniaceae | Cassia mimosoides L. | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Caesalpiniaceae | Cassia occidentalis L. | Whole plant extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Caesalpiniaceae | Cassia occidentalis L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Caesalpiniaceae | Cassia tora L. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Caesalpiniaceae | Cassia tora L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Campanulaceae | Codonopsis clematidea Schrenk | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Cannabaceae | Cannabis sativa L. | Essential oil from panicles | 0.10% | G | Adult females | 83.28% of mortality | β-myrcene (18.5%), trans-caryophyllene (35.6%) | [98] |
| Cannabaceae | Cannabis sativa L. | Aerial part and root extracts | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Cannabaceae | Humulus lupulus L. | Flower extract | 5-50% | A,B | Adult females | Mortalities of 56.37% and 67.84%, respectively | - | [60] |
| Cappandaceae | Clome viscosa L. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Cappandaceae | Clome viscosa L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Capparidaceae | Boscia senagalensis (Pers.) Lam. Ex. Poir. | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Capparidaceae | Boscia senagalensis (Pers.) Lam. Ex. Poir. | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Caprifoliaceae | Sambucus nigra L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Caryophyllaceae | Saponaria officinalis L. | Root extract | 0.31% (eggs), 1.18% (adulst), 0.91% (oviposition) w/v | I | Eggs, adults and oviposition | LD50 | - | [66] |
| Caryophyllaceae | Silene sussamyrica Lazkov | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Chenopodiaceae | Anabasis aphylla L. | Seed and bark extracts | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Chenopodiaceae | Anthochlamis tianschanica Iljin | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Chenopodiaceae | Chenopodium álbum (L.) Mosc. Ex. Moq. | Flower and leaf extracts | 8–50%. | A,B | Adult females | Mortalities of 96.99% and 91.15%, respectively | - | [60] |
| Combretaceae | Cobretum micranthum G. Don | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Cobretum micranthum G. Don | Whole plant extract | 2500 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Combretum glutinosum Perr. Ex. DC. | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Combretum glutinosum Perr. Ex. DC. | Leaf extract | 2500 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Combretum glutinosum Perr. Ex. DC. | Stem extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Combretaceae | Combretum glutinosum Perr. Ex. DC. | Stem extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Combretaceae | Guiera senegalensis J.F. Gmel. | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Guiera senegalensis J.F. Gmel. | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Combretaceae | Guiera senegalensis J.F. Gmel. | Stem extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Combretaceae | Guiera senegalensis J.F. Gmel. | Stem extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Combretaceae | Piloitigma vetilicolin | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Combretaceae | Piloitigma vetilicolin | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Convolvulaceae | Convolvulus arvensis L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Convolvulaceae | Convolvulus krauseanus Regel. & Schmalh | Root extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Convolvulaceae | Ipomaea asarifolia (Desr.) Roem. & Schult. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Convolvulaceae | Ipomaea asarifolia (Desr.) Roem. & Schult. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Convolvulaceae | Ipomaea sp. L. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Convolvulaceae | Ipomaea sp. L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Cupressaceae | Cupressus macrocarpa Hartw. ex Gordon | Leaf extract | 5.69 µL/L air | A | Adult females | LD50 | β-citronellol (35.92%) | [55] |
| Cupressaceae | Cupressus sempervirens L. | Essential oil | 0.1%. | G | Adult females | 28.9% of mortality | - | [112] |
| Cupressaceae | Juniperus communis L. | Essential oil | 0.1%. | G | Adult females | 42.6% of mortality | - | [112] |
| Cupressaceae | Juniperus phoenicea L. | Essential oil | - | D | Larvae and adults | Mortalities of 60% (larvae) and 56% (adults) | - | [88] |
| Cupressaceae | Thuja orientalis L. | Leaf extract | 7.51 µL/L air | A | Adult females | LD50 | α-pinene (35.49%) | [55] |
| Elaeagnaceae | Elaeagnus angustifolia L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Equisetaceae | Equisetum arvense L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Euforbiaceae | Jatropha curcas L. | Leaf extract | 0.06% | C,G | Adult females | Mortality of 63.3% | - | [124] |
| Euphorbiaceae | Chrozophora oblongifolia (Delile) Spreng. | Whole plant extract | 312.72 and 206.91 ppm | G | Adult females and larvae | LD50 | 7-O-β-D-[2”,6”-bis(4-hydroxy-E-cinnamoyl)] glucopyranoside, apigenin 7-O-ß-D-glucopyranoside isolated from butanol fration | [125] |
| Euphorbiaceae | Cnidoscolus aconitifolius (Mill) I.M. Johnst. | Leaf extract | 2000 µg/mL | C,G | Adult females | 92% of mortality | - | [61] |
| Euphorbiaceae | Euphorbia ferganensis B. Fedtsch. | Root extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Euphorbiaceae | Euphorbia kansui S.L. Liou S.B. Ho | Root extract | 3-5 g/L | C | Adult females | Mortalities of 27% and 55%, respectively | 3-O-(2,3-dimethylbutanoyl)-13-dodecanoylingenol y 3-O-(2′E,4′Z-decadienoyl)-ingenol | [100] |
| Fabaceae | Acacia cyanophylla Lindl. | Essential oil | - | D | Larvae and adults | Mortalities of 58% (larvae) and 26% (adults) | - | [88] |
| Fabaceae | Amnopiptanthus nanus (M. pop) Cheng | Pod extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Fabaceae | Bowdichia virgilioides Kunth | Leaf extract | 0.06% w/v | C,G | Adult females | Mortality of 64.4% | [126] | |
| Fabaceae | Gleditschia spp. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Fabaceae | Glycirrhisa uralensis L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Fabaceae | Hedysarum cephalotes Franchet | Whole plant extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Fabaceae | Hedysarum daraut-kurganicum Sultanova | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Fabaceae | Hymenaea courbaril L. | Leaf extract | 0.06% w/v | C,G | Adult females | Mortality of 59.4% | [126] | |
| Fabaceae | Medicago minima L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Fabaceae | Melilotus officinalis L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Fabaceae | Millettia pinnata L. | Laef oil | 0.004% | C | Adult females | LD50 (after 4 days) | - | [101] |
| Fabaceae | Oxytropis rosea Bunge | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Fabaceae | Sophora korolkovii Koehne. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Fabaceae | Sophora secundiflora (Ortega) Lag. Ex. DC. | Essential oil | - | D | Larvae and adults | Mortalities of 68% (larvae) and 61% (adults) | - | [88] |
| Fabaceae | Vicia cracca L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Geraniaceae | Pelargonium graveolens L’Her | Leaf extract | 12.27 µL/L air | A | Adult females | LD50 | terpinen-4-ol (20.29%) | [55] |
| Geraniaceae | Pelargonium graveolens L’Hér. | Essential oil | 19 × 10−3 µL/mL of air. | K | Adults | 100% of mortality | - | [42] |
| Geraniaceae | Pelargonium graveolens L’Hér. | Essential oil | - | D | Larvae and adults | Mortalities of 78% (larvae) and 70% (adults) | - | [88] |
| Geraniaceae | Pelargonium roseum Willd | Essential oil | 0.1%. | G | Adult females | 30% of mortality | - | [112] |
| Gramineae | Chrysopogon zizanioides (L.) | Essential oil | 18.82 μg/mL | J | Adult females | LD50 | - | [127] |
| Gramineae | Cymbopogon citratus (DC.) Stapf | Essential oil | 19 × 10−3 µL/mL of air. | J | Adults | 100% of mortality | - | [42] |
| Gramineae | Cymbopogon citratus (DC.) Stapf | Essential oil | 0.1%. | G | Adult females | 17.8% of mortality | - | [112] |
| Gramineae | Cymbopogon flexuosus (Nees ex Steud.) W. Watson | Essential oil | 17.23 μg/mL | J | Adult females | LD50 | - | [127] |
| Gramineae | Cymbopogon Martini (Roxb.) W. Watson | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 67% of mortality | - | [42] |
| Gramineae | Cymbopogon nardus (L) Rendle | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 99% of mortality | - | [42] |
| Gramineae | Cymbopogon nardus (L) Rendle | Essential oil | 22.5 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Gramineae | Cymbopogon winterianus Jowitt ex. Bor | Essential oil | 0.1% | G | Adult females | 27.6% of mortality | - | [112] |
| Gramineae | Lolium perenne L. | Leaf and flower methanolic extracts | 6-50% | A,B | Adult females | Mortalities of 91.43% and 93.5%, respectively | - | [60] |
| Guttiferae | Hypericum perforatum L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Iridaceae | Iris sogdiana Regel. | Leaf extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Juglandecaea | Juglans regia L. | Leaf extract | 12% v/w | C,G | Adult females and nymphs | Mortality between 83 and 90% | - | [128] |
| Lamiaceae | Acinos thymoides (L.) Moench | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Lamiaceae | Ajuga australis R.Br. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Callicarpa pedunculata R.Br. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Ceratanthus longicornis (F.Muell.) G. Taylor | Leaf extract | 1% | C | - | 100% of mortality | - | [103] |
| Lamiaceae | Clerodendrum floribundum R.Br. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Clerodendrum inerme (L.) Gaertn. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Clerodendrum tomentosum (Vent.) R.Br. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Clerodendrum traceyi F. Muell. | Leaf extract | 1% | C | - | 100% of mortality | - | [103] |
| Lamiaceae | Faradaya albertissii F. Muell. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Faradaya splendida F. Muell. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Glossocarya calcicola Domin. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Glossocarya hemiderma Benth. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Gmelina leichardtii (F.Muell.) Benth | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Hemiandra australis B.J. Conn. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Hemiandra leiantha Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Hemiandra pungens R.Br. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Hemigenia humilis Benth. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Hemigenia sericea Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Hemigenia westringioides Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Hyssopus officinalis L. | Aerial part extract | 1% | G | Adult females | Less than 20% of mortality | - | [99] |
| Lamiaceae | Hyssopus officinalis L. | Essential oil | 0.1%. | G | Adult females | 28.1% of mortality | - | [112] |
| Lamiaceae | Lachnostachys eriobotrya (F. Muell.) Druce | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Lavandula angustifolia Mill. | Leaf extract | 4.93 µL/L | J | Adult females | LD50 | 1,8-cineole, camphor, β-pinene | [129] |
| Lamiaceae | Lavandula latifolia Medik. | Essential oil from twigs with leaves andflowers | 0.20–0.25% v/v | A,C | Adult females | Mortality between 95 and 100% | linalool (37.8%), 1,8-cineole (24.9%), camphor (18.7%) | [56] |
| Lamiaceae | Lavandula officinalis Chaix | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 97% of mortality | - | [42] |
| Lamiaceae | Lavandula officinalis Chaix | Essential oil | - | D | Larvae and adults | Mortalities of 38% (larvae) and 41% (adults) | - | [88] |
| Lamiaceae | Lavandula vera DC. | Essential oil | 0.1% | G | Adult females | 26.1% of mortality | - | [112] |
| Lamiaceae | Leonorus turkestanicus V. Krecz. & Kupr. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Lamiaceae | Lycopus australis R. Br. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Majorana hortensis Moench | Essential oil | 1.84–6.26% | C | Adults and eggs (respectively) | LD50 | terpinen-4-ol (23.86%), p-cymene (23.40%) and sabinene (10.90%) | [89] |
| Lamiaceae | Melissa officinalis L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Lamiaceae | Menta spicata L. | Essential oil | 15 μL/L | C,J | Eggs | LD50 | carvone (68.5%) | [130] |
| Lamiaceae | Mentha arvensis L. | Aerial part extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Lamiaceae | Mentha longifolia L. | Essential oil | 11.08 μg/mL | J | Adult females | LD50 | - | [127] |
| Lamiaceae | Mentha piperita L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 100% of mortality | - | [42] |
| Lamiaceae | Mentha piperita L. | Essential oil | 22.8 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Lamiaceae | Mentha piperita L. | Essential oil | 0.1%. | G | Adult females | 23.7% of mortality | - | [112] |
| Lamiaceae | Mentha piperita L. | Essential oil | 15.86 μg/mL | J | Adult females | LD50 | - | [127] |
| Lamiaceae | Mentha pulegium L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 100% of mortality | - | [42] |
| Lamiaceae | Mentha pulegium L. | Essential oil | 23.7 µg/cm3 | J | Adults | LD50 | - | [68] |
| Lamiaceae | Mentha pulegium L. | Essential oil | - | D | Larvae and adults | Mortalities of 90% (larvae) and 91% (adults) | - | [88] |
| Lamiaceae | Mentha spicata L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 100% of mortality | - | [42] |
| Lamiaceae | Mentha spicata L. | Essential oil | 38.8 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Lamiaceae | Mentha spicata L. | essential oil from leaves | 7.53 µL/L air | C,J | Adult females | LD50 | carvone (59.4%), limonene (9.8%), 1,8-cineole (7.4%) | [85] |
| Lamiaceae | Mentha sylvestris L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Lamiaceae | Microcorys capitata (Bartl.) Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Microcorys sp. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Micromeria fruticosa L. | Essential oil vapors | 2 µL/L of air | J | Adults and nimphs | 96.7% of mortality | - | [105] |
| Lamiaceae | Micromeria fruticosa L. | Essential oil vapors | 2 µL/L | J | Adults and nimphs | 96.7% of mortality | - | [105] |
| Lamiaceae | Nepeta racemosa L. | Essential oil vapors | 2 uL/L of air | J | Adults and nimphs | 95% of mortality | - | [105] |
| Lamiaceae | Nepeta racemosa L. | Essential oil vapors | 2 µL/L | J | Adults and nimphs | 95% of mortality | - | [105] |
| Lamiaceae | Ocimum basilicum L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 88% of mortality | - | [42] |
| Lamiaceae | Ocimum basilicum L. | Essential oil | 39.5 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Lamiaceae | Ocimum basilicum L. | Essential oil | 0.1% | G | Adult females | 21% of mortality | - | [112] |
| Lamiaceae | Ocimum basilicum L. | Essential oil | 0.6 μL/L | C,J | Adult females | LD50 | linalool (65.7%) | [130] |
| Lamiaceae | Origanum majorana L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 92% of mortality | - | [42] |
| Lamiaceae | Origanum majorana L. | Essential oil | 0.1% | G | Adult females | 7.1% of mortality | - | [112] |
| Lamiaceae | Origanum vulgare L. | Essential oil | 8.52 µL/L air | A | Adult females | LD50 | pulegone (77.45%) | [55] |
| Lamiaceae | Origanum vulgare L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Lamiaceae | Origanum vulgare L. | Essential oil vapors | 2 uL/L of air | J | Adults and nimphs | 95% of mortality | - | [105] |
| Lamiaceae | Origanum vulgare L. | Essential oil vapors | 2 µL/L | J | Adults and nimphs | 95% of mortality | - | [105] |
| Lamiaceae | Otostelgia olgae (Regel.) Korsch. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Lamiaceae | Pityrodia bartlingii (Lehm.) Benth. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Pityrodia verbascina (F. Muell.) Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus actites P.I. Forst. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus alloplectus S.T. Blake | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Plectranthus amoenus P.I. Forst | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus apreptus S.T. Blake | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus argentatus S.T. Blake | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus cremnus B. J. Conn. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus diversus S.T. Blake | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Plectranthus fasciculatus P.I. Forst. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus foetidus Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus glabriflorus P.I. Forst | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Plectranthus gratus S.T. Blake | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus graveolens R.Br. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus habrophyllus P.I. Forst | Leaf extract | 1% | C | - | 100% of mortality | - | [103] |
| Lamiaceae | Plectranthus Koonyum Range | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus leiperi P.I. Forst. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus mirus S.T. Blake | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus nitidus P.I. Forst. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus omissus P. I. Forst. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus parviflorus Willd | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus scutellarioides (L.) R.Br. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Plectranthus sp. buchanans Fort | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus sp. Hann Tableland | Leaf extract | 1% | C | - | 100% of mortality | - | [103] |
| Lamiaceae | Plectranthus sp. Pinnacle | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus spectabilis S.T. Blake | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Plectranthus suaveolens S.T. Blake | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Pogostemon cablin Benth. | Essential oil | 0.1% | G | Adult females | 20.3% of mortality | - | [112] |
| Lamiaceae | Premna acuminata R.Br. | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Premna serratifolia L. | Leaf extract | 1% | C | - | 100% of mortality | - | [103] |
| Lamiaceae | Prostanthera incisa Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Prostanthera lasianthos Labill. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Prostanthera nivea A. Cunn. Ex. Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Prostanthera rotundifolia R.Br. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Prostanthera spinosa F. Muell. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Prostanthera stricta R.T. Baker | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Rosmarinus officinalis L. | Essential oil | 0.10, 0.15, 0.20, and 0.25%. | A,C | Adult females and eggs | Mortalities of 15%, 79%, 100% and 100% for females, respectively | 1,8-cineole (26.7%), camphor (17.5%), α-pinene (18.6%), camphene (11.8%), myrcene (9%), bornyl acetate (4%), β-pinene (2.8%), humulene (0.5%), borneol (1.8%), β-caryophyllene (1.5%), linalool (1%), Verbennone (0.9%), α-terpineol (0.8%) | [53] |
| Lamiaceae | Rosmarinus officinalis L. | Essential oil | - | D | Larvae and adults | Mortalities of 61% (larvae) and 53% (adults) | - | [88] |
| Lamiaceae | Rosmarinus officinalis L. | Essential oil | 10 mL/L | D | - | LD50 | 1,8-cineole and α-pinene (mortalities of 88 ± 4.8% and 32 ± 4.8%, respectively with each compound) | [104] |
| Lamiaceae | Rosmarinus officinalis L. | Essential oil | 0.1% | G | Adult females | 11.7% of mortality | - | [112] |
| Lamiaceae | Salvia desertorum | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Lamiaceae | Salvia fruticosa Mill. | Leaf extract | 3.77 µL/L | J | Adult females | LD50 | [129] | |
| Lamiaceae | Salvia officinalis L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 100% of mortality | - | [42] |
| Lamiaceae | Salvia officinalis L. | Essential oil | 0.10%. 0.15%. 0.20% and 0.25% | A,C | Adult females and eggs | 100% of female mortality in all concentrations | α-tujone (42.5), 1,8-cineole (10.3%), camphor (11%), α-pinene (6.7%), camphene (6.5%), β-tujone (6.6%), myrcene (1.4%), bornyl acetate (0.7%), β-pinene (3.4%), humulene (2.4%), viridiflorol (2.2%), borneol (2%), β-caryophyllene (1.5%), cymene (1%) | [53] |
| Lamiaceae | Salvia officinalis L. | Essential oil | 63.7 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Lamiaceae | Salvia officinalis L. | Essential oil | - | D | Larvae and adults | Mortalities of 61% (larvae) and 57% (adults) | - | [88] |
| Lamiaceae | Salvia sclarea L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 61% of mortality | - | [42] |
| Lamiaceae | Salvia sclarea L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Lamiaceae | Salvia sclarea L. | Essential oil | 0.1%. | G | Adult females | 71% of mortality | - | [112] |
| Lamiaceae | Salvia vvedenskyi E. Nikit. | Root extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Lamiaceae | Satureja sahendica Bornm. | Essential oil vapors | 0.98 µL/L (females), 0.54 µL/L (eggs) | J | Eggs and adults | LD50 | - | [84] |
| Lamiaceae | Satureja sahendica Bornm. | Essential oil | 0.54% | E | Adults | LD50 | - | [83] |
| Lamiaceae | Scutellaria mollis R. Br. | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Stachys tschatkalensis Knorr. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Lamiaceae | Teucrium racemosum R. Br. | Leaf extract | 1% | C | - | Mortality between 20 and 49% | - | [103] |
| Lamiaceae | Thymbra capitata (L.) Cav. | Essential oil | - | D | Larvae and adults | Mortalities of 61% (larvae) and 52% (adults) | - | [88] |
| Lamiaceae | Thymus vulgaris L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 93% of mortality | - | [42] |
| Lamiaceae | Thymus vulgaris L. | Essential oil | 22.7 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Lamiaceae | Thymus vulgaris L. | Essential oil | 0.1% | G | Adult females | 62.2% of mortality | - | [112] |
| Lamiaceae | Vitex lignum-vitae Schauer | Leaf extract | 1% | C | - | Mortality between 50 and 89% | - | [103] |
| Lamiaceae | Viticipremna queenslandica Munir | Leaf extract | 1% | C | - | Mortality between 90 and 99% | - | [103] |
| Lamiaceae | Westringia eremicola A. Cunn. Ex. Benth. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Westringia glabra R.Br. | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Westringia saxatilis B.J. Conn | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Westringia viminalis B.J. Conn & Tozer | Leaf extract | 1% | C | - | Less than 20% of mortality | - | [103] |
| Lamiaceae | Ziziphora clinopodioides Lam. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Lauraceae | Cinnamomum zeylandicum Blume | Essential oil | 0.1% | G | Adult females | 23.6% of mortality | - | [112] |
| Lauraceae | Laurus nobilis L. | Leaf extract | 17–50% | A,B | Adult females | Mortalities of 66.11% and 69.72%, respectively | - | [60] |
| Lauraceae | Laurus nobilis L. | Essential oil | - | D | Larvae and adults | Mortalities of 63% (larvae) and 46% (adults) | - | [88] |
| Lauraceae | Licaria puchury-major (Mart.) Kosterm. | Essential oil | 30.8 µg/mL | J | Adult females | LD50 | safrole (38.8%), 1,8-cineole (21.7%) | [131] |
| Lilliaceae | Convallaria majalis L. | Root extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Malvaceae | Abutilon theophasti Medic. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Malvaceae | Corchorus sp. | Whole plant extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Malvaceae | Corchorus sp. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Malvaceae | Hybiscus sp. | Whole plant extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Malvaceae | Hybiscus sp. | Whole plant extract | 2500 ppm | G | Adults | - | - | [71] |
| Malvaceae | Malva pusilla Sm. | Whole plant extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Meliaceae | Azadirachta indica A. Juss. | Commercialformulation | 1%. | C,D | Adult females | 97.5% of mortality | - | [37] |
| Meliaceae | Azadirachta indica A. Juss. | Leaf extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Meliaceae | Azadirachta indica A. Juss. | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Meliaceae | Melia azedarach L. | Fruit extract | 14–50% | A,B | Adult females | Mortalities of 74.57% and 76.45%, respectively | - | [60] |
| Meliaceae | Melia azedarach L. | Essential oil | - | D | Larvae and adults | Mortalities of 77% (larvae) and 75% (adults) | - | [88] |
| Meliaceae | Melia azedarach L. | Acetone and petroleum ether methanolic extracts | - | C | Larvae | Lethal and fecundity effects | - | [107] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Stem extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Stem extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Fruit extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Mimosaceae | Prosopis chinensis (Molina) Stuntz | Fruit extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Myrtaceae | Callistemon viminals (Sol. ex Gaertn.) G. Don | Leaf extract | 40.66 µL/L air | A | Adult females | LD50 | 1,8-cineole (71.77%) | [55] |
| Myrtaceae | Eucalyptus camaldulensis Dehnh. | Leaf extract | 18–50% | A,B | Adult females | Mortalities of 62.61% and 55.57%, respectively | - | [60] |
| Myrtaceae | Eucalyptus camaldulensis Dehnh. | Flower extract | 20–50% | A,B | Adult females | Mortalities of 51.91% and 47.15%, respectively | - | [60] |
| Myrtaceae | Eucalyptus citriodora Hook | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 100% of mortality | - | [42] |
| Myrtaceae | Eucalyptus citriodora Hook | Essential oil | 19.3 µg/cm3 | Adults | LD50 | - | [68] | |
| Myrtaceae | Eucalyptus ghomphocephala A. Cunn. Ex. DC. | Essential oil | - | D | Larvae and adults | Mortalities of 60% (larvae) and 34% (adults) | - | [88] |
| Myrtaceae | Eucalyptus globulus Labill. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 89% of mortality | - | [42] |
| Myrtaceae | Eucalyptus globulus Labill. | Essential oil | 0.1% | G | Adult females | 19.7% of mortality | - | [112] |
| Myrtaceae | Eucalyptus microtheca F. Muell. | Essential oil vapors from fruits and leaves | 1.52 µL/L (females), 5.7 µL/L (eggs) | J | Eggs and adults | LD50 | - | [84] |
| Myrtaceae | Eucalyptus microtheca F. Muell. | Fruits and leaves essential oils | 0.56% (leaves), 2.36% (fruits) | E | Adults | LD50 | - | [83] |
| Myrtaceae | Eucalyptus oleosa L. | Essential oil | 2.42 µL/L air | J | Adult females | LD50 | 1,8-Cineole (31.96%), α-pinene (15.25%), trans-anethole (7.32%) | [132] |
| Myrtaceae | Eucalyptus radiata Sieber ex. DC. | Essential oil | 0.1% | G | Adult females | 27.9% of mortality | - | [112] |
| Myrtaceae | Eucalyptus torquata L. | Essential oil | 3.59 µL/L air | J | Adult females | LD50 | 1,8-cineole (28.57%), α-pinene (15.74%), globulol (13.11%) | [132] |
| Myrtaceae | Eucapyptus sp. | Essential oil | 2.18–7.33% | C | Adults and eggs | LD50 | - | [89] |
| Myrtaceae | Eugenia caryophyllata Thunb. | Bud and leaf essential oils | 19 × 10−3 µL/mL of air | J | Adults | Mortalities of 80% (buds) and 66% (leaves) | - | [42] |
| Myrtaceae | Eugenia caryophyllata Thunb. | Essential oil | 23.6 µg/cm3 | C,J | Adults | LD50 | - | [68] |
| Myrtaceae | Melaleuca alternifolia Maiden & Betche ex. Cheel | Essential oil | 0.1% | G | Adult females | 28.6% of mortality | - | [112] |
| Myrtaceae | Melaleuca leucadendron L. | Essential oil | 0.1% | G | Adult females | 23.5% of mortality | - | [112] |
| Myrtaceae | Melaleuca viridiflora Sol. Ex. Gaertn. | Essential oil | 0.1% | G | Adult females | 26.8% of mortality | - | [112] |
| Myrtaceae | Myrtus communis L. | Essential oil | - | D | Larvae and adults | Mortalities of 82% (larvae) and 47% (adults) | - | [88] |
| Myrtaceae | Pimenta racemosa (Mill.) J.W. Moore | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 60% of mortality | - | [42] |
| Myrtaceae | Syzygium aromaticum (L.) Merr. & L.M. Perry | Essential oil from flower buds | 6.13 µL/L air | C,J | Adult females | LD50 | eugenol (78.5%), β-caryophyllene (13.8%) | [85] |
| Myrtaceae | Syzygium aromaticum (L.) Merr. & L.M. Perry | Essential oil | 0.1% | G | Adult females | 41.3% of mortality | - | [112] |
| Myrtaceae | Syzygium cumini (L.) Skeels | Ethanolic, hexane and ether ethyl acetate extracts | 75. 150 and 300 µg/mL | C | Adult females | Mortalities of 98.5% (ethanolic extract), 94% (hexane extract) and 90% (ether-ethyl acetate extract) | - | [108] |
| Nitrariaceae | Peganum harmala L. | Essential oil | - | D | Larvae and adults | Mortalities of 34% (larvae) and 12% (adults) | - | [88] |
| Nitrariaceae | Peganum harmala L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Nyctaginaceae | Bougainvilleae spectabilis Willd. | Leaf extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Nyctaginaceae | Bougainvilleae spectabilis Willd. | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Papaveraceae | Chelidonium majus L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Papaveraceae | Papaver pavoninum Schrenk. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Papaveraceae | Papaver rhoeas L. | Essential oil | - | D | Larvae and adults | Mortalities of 43% (larvae) and 34% (adults) | - | [88] |
| Papaveraceae | Papaver rhoeas L. | Aerial part extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Papaveraceae | Roemeria refracta DC. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Pinaceae | Cedrus atlantica (Endl.) Manetti ex. Carriére | Essential oil | 0.1% | G | Adult females | 12.4% of mortality | - | [112] |
| Pinaceae | Picea schrenkiana Fisch & Mey. | Leaf extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Pinaceae | Pinus sylvestris L. | Essential oil | 0.1% | G | Adult females | 50.4% of mortality | - | [112] |
| Piperaceae | Piper aduncum L. | Leaf essential oil compounds | - | J | - | Mortality effect [(E)-nerolidol, α–Humulene and β-caryophyllene)] and repellence (β-caryophyllene) | (E)-nerolidol, α–Humulene and β-caryophyllene | [109] |
| Piperaceae | Piper nigrum L. | Essential oil | 0.1% | G | Adult females | 22.8% of mortality | - | [112] |
| Plantaginaceae | Globularia alypum L. | Essential oil | - | D | Larvae and adults | Mortalities of 8% (larvae) and 2% (adults) | - | [88] |
| Plantaginaceae | Plantago major L. | Aerial part extract | 1% | G | Adult females | Mortality between 80 nd 100% | - | [99] |
| Plumbaginaceae | Limonium tianschanicum Lincz. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Polygonaceae | Polygonum aviculare L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Polygonaceae | Polygonum toktoquilicum Lazkov | Root extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Polygonaceae | Rumex acetosa L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Primulaceae | Anagallis arvensis L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Ranunculaceae | Aconitum soongaricum Stapf | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Ranunculaceae | Adonis parviflora Fisch. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Ranunculaceae | Ceratocephallus testiculata (Crantz.) Bess. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Ranunculaceae | Clematis orientalis L. | Seed extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Ranunculaceae | Clematis songarica Bge. | Seed extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Ranunculaceae | Nigella sativum L. | Seed extract | 708.57 ppm | G | Adult females | LD50 | - | [91] |
| Ranunculaceae | Ranunculus polyanthemus L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Rosaceae | Geum urbanum L. | Aerial part extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Rosaceae | Padus avium Mill. | Leaf extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Rosaceae | Prunus laurocerasus L. | Leaves, flower and seed extract | 12% v/w | A,C | Eggs and adult females | Mortality between 37 and 100% | - | [54] |
| Rubiaceae | Boirerio radiata | Whole plant extract | 5000 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Rubiaceae | Boirerio radiata | Whole plant extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Rubiaceae | Galium verum L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Rubiaceae | Gardenia jasminoides J. Ellis | Fruits extract | 10000 ppm | I,J | Adult females and nymphs | 49 and 66% of mortality, respectively | - | [133] |
| Rutaceae | Citrus aurantium L. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 68% of mortality | - | [42] |
| Rutaceae | Citrus aurantium L. | Essential oil | - | D | Larvae and adults | Mortalities of 63% (larvae) and 55% (adults) | - | [88] |
| Rutaceae | Citrus aurantium L. | Fruit epicarp essential oil | 1% | I,J | Adult females | Repellent effect due to all 27 identified compounds | d-limonene | [67] |
| Rutaceae | Citrus aurantium L. var. Armara | Essential oil | 0.1% | G | Adult females | 21.4% of mortality | - | [112] |
| Rutaceae | Citrus bergamia Risso & Poit. | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 87% of mortality | - | [42] |
| Rutaceae | Citrus bergamia Risso & Poit. | Essential oil | 0.1% | G | Adult females | 11% of mortality | - | [112] |
| Rutaceae | Citrus limon (L.) Burm. F. | Essential oil | 0.1% | G | Adult females | 34.9% of mortality | - | [112] |
| Rutaceae | Citrus paradisi Macfad | Essential oil | 6.96 µL/L air | A | Adult females | LD50 | limonene (74.29%) | [55] |
| Rutaceae | Citrus paradisi Macfad. | Essential oil | 0.1% | G | Adult females | 30.6% of mortality | - | [112] |
| Rutaceae | Citrus sinensis Osbeck | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 61% of mortality | - | [42] |
| Rutaceae | Citrus sinensis Osbeck | Fruit epicarp essential oil | 1% | I,J | Adult females | Repellent effect due to all 27 identified compounds | d-limonene | [67] |
| Rutaceae | Citrus sinensis Osbeck | Essential oil | 0.1% | G | Adult females | 45.6% of mortality | - | [112] |
| Rutaceae | Haplophyllum tuberculatum (Forssk.) A. Juss. | Essential oil | - | D | Larvae and adults | Mortalities of 94% (larvae) and 93% (adults) | - | [88] |
| Rutaceae | Ruta chalepensis L. | Essential oil | - | D | Larvae and adults | Mortalities of 66% (larvae) and 61% (adults) | - | [88] |
| Rutaceae | Zanthoxylum armatum DC. | Leaf extract | 5000 and 10000 ppm | G | Adults | Mortalities of 36% and 39%, respectively | - | [111] |
| Santalaceae | Santalum sp. | Essential oil | 0.1% | G | Adult females | 87.2% of mortality and fecundity decrease | - | [112] |
| Scrophulariaceae | Calceolaria andina Benth | Two extract compounds | 80 and 30 ppm | G | - | LD50 | 2-(1,1-dimethylprop-2-enyl)-3-hydroxi-1,4-naphthoquinone and 2-acetoxy-3-(1,1-dimethylprop-2-enyl)-1,4-naphthoquinone | [113] |
| Scrophulariaceae | Verbascum thapsus L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Simarubaceae | Ailanthus altissima (Mill.) Swingle | Leaf extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Simarubaceae | Quassia sp. | Aerial part extract | 10000 ppm and 47 ppm (chaparinone) | C | - | LD50 (chaparinone) | Chaparinone quasinoid | [114] |
| Solanaceae | Capsicum annuum L. | Aerial part extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Solanaceae | Capsicum annuum L. | Fruit extract | - | B,J | Adult females | 45% of mortality | - | [116] |
| Solanaceae | Capsicum baccatum L. | Fruit extract | - | B,J | Adult females | Repellent effect | - | [116] |
| Solanaceae | Capsicum chinense Jacq. | Fruit extract | - | B,J | Adult females | Repellent effect | - | [116] |
| Solanaceae | Capsicum frutescens L. | Fruit extract | - | B,J | Adult females | Repellent effect | - | [116] |
| Solanaceae | Datura stramonium L. | Seed and leaf extracts | 167.25 (leaves) and 145.75 (seeds) mg/L | C,D | Adults | Mortalities of 98% (leaves) and 25% (seeds) | - | [5] |
| Solanaceae | Hyoscyamus niger L. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80%. | - | [99] |
| Solanaceae | Lycopersicon hirsutum Dunal | - | - | J | Adult females | Repellent activity | Dihidrofarnesoic acid | [115] |
| Solanaceae | Solanum nigrum L. | Flower and leaf extracts | 12–50% | A,B | Adult females | Mortalities of 69.88% and 79.36%, respectively | - | [60] |
| Solanaceae | Solanum nigrum L. | Fruit extract | 19–50% | A,B | Adult females | Mortalities of 68.78% and 53.29%, respectively | - | [60] |
| Solanaceae | Solanum nigrum L. | Leaf extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Solanaceae | Solanum nigrum L. | Leaf extract | 279.69 µg/mL | C,G | Adult females | LD50 after 72 h | - | [134] |
| Sterculiaceae | Waltheria indica L. | Whole plant extract | 5000 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
| Sterculiaceae | Waltheria indica L. | Whole plant extract | 2500 ppm | G | Adults | Mortality between 40 and 60% | - | [71] |
| Styracaceae | Styrax officinalis L. | Seed cover extract | 16–50% | A,B | Adult females | Mortalities of 64.11% and 73.25%, respectively | - | [60] |
| Styracaceae | Styrax officinalis L. | Seed extract | 21–50% | A,B | Adult females | Mortalities of 68.17% and 31.28%, respectively | - | [60] |
| Umbelliferae | Angelica tschimganica (Korov.) B. Tikhom. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Umbelliferae | Conium maculatum L. | Seed extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Umbelliferae | Dorema microcarpum Korov. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Umbelliferae | Ferula foetida (Bunge) Regel. | Root extract | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Umbelliferae | Ferula foetidissima Regel. & Schmahl. | Root extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Umbelliferae | Ferula inciso-serrata M. Pimen. & Baranova | Aerial part and root extracts | 1% | G | Adult females | Mortality between 0 and 20% | - | [99] |
| Umbelliferae | Heracleum dissectum Ledeb. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50% | - | [99] |
| Umbelliferae | Mediasia macrophylla (Regel. & Schmahl.) M. Pimen. | Aerial part extract | 1% | G | Adult females | Mortality between 50 and 80% | - | [99] |
| Umbelliferae | Prangos lipskyi Korov | Root extract | 1% | G | Adult females | Mortality between 80 and 100% | - | [99] |
| Urticaceae | Urtica pilulifera L. | Essential oil | - | D | Larvae and adults | Mortalities of 49% (larvae) and 46% (adults) | - | [88] |
| Valerianaceae | Valeriana officinalis L. | Aerial part extract | 1% | G | Adult females | Mortality between 20 and 50%. | - | [99] |
| Verbenaceae | Lantana camara L. | Essential oil | - | D | Larvae and adults | Mortalities of 2% (larvae) and 1% (adults) | - | [88] |
| Verbenaceae | Lippia origanoides H.B.K. | Essential oil | 25.1 μg/mL | J | Adult females | LD50 | carvacrol (48.31%), p-cymene (9.11%), thymol (8.78%) | [135] |
| Verbenaceae | Lippia sidoides Cham. | Essential oil | 0.01 µL/L (extract). 0.001 µL/L (thymol). 3.02 µL/L (p-cymene). 0.08 µL/L (β-caryophyllene) and 0.036 µL/L (carvacrol) | J | Adult females | LD50 | Thymol, p-cymene, β-caryophyllene and carvacrol | [117] |
| Zingiberaceae | Elettaria cardamomum (L.) Maton | Essential oil | 19 × 10−3 µL/mL of air | J | Adults | 87% of mortality | - | [42] |
| Zingiberaceae | Zingiber officinale Rosc. | Essential oil | 0.1% | G | Adult females | 11.9% of mortality | - | [112] |
| Zygophyllaceae | Balanites aegyptiaca (L.) Delile | Leaf extract | 5000 ppm | G | Adults | More than 80% of mortality | - | [71] |
| Zygophyllaceae | Balanites aegyptiaca (L.) Delile | Leaf extract | 2500 ppm | G | Adults | Mortality between 61 and 80% | - | [71] |
References
- Kumari, S.; Chauhan, U.; Kumari, A.; Nadda, G. Comparative toxicities of novel and conventional acaricides against different stages of Tetranychus urticae Koch (Acarina: Tetranychidae). J. Saudi Soc. Agric. Sci. 2017, 16, 191–196. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Landeros, J.; Ail, C.E.; Cerna, E.; Ochoa, Y.; Guevara, L.; Aguirre, L.A. Susceptibility and resistance mechanisms of Tetranychus urticae (Acariformes: Tetranychidae) in greenhouse roses. Rev. Colomb. Entomol. 2010, 36, 5–9. [Google Scholar]
- Hoy, M.A. Agricultural Acarology: Introduction to Integrated Mite Management; CRC Press: Boca Ratón, FL, USA, 2011. [Google Scholar]
- Kumral, N.A.; Çobanoğlu, S.; Yalcin, C. Acaricidal, repellent and oviposition deterrent activities of Datura stramonium L. against adult Tetranychus urticae (Koch). J. Pest Sci. 2009, 83, 173–180. [Google Scholar] [CrossRef]
- Breeuwer, J.A.J.; Jacobs, G. Wolbachia: Intracellular manipulators of mite reproduction. Exp. Appl. Acarol. 1996, 20, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Meena, N.K.; Rampal; Barman, D.; Medhi, R.P. Biology and seasonal abundance of the two-spotted spider mite, Tetranychus urticae, on orchids and rose. Phytoparasitica 2013, 41, 597–609. [Google Scholar] [CrossRef]
- Tehri, K. A review on reproductive strategies in two spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). J. Entomol. Zool. Stud. 2014, 2, 35–39. [Google Scholar]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef]
- Reis, P.R.; Silva, E.A.; Zacarias, M.S. Controle biológico de ácaros em cultivos protegidos. Inf. Agropecuário 2005, 26, 58–68. [Google Scholar]
- Mohankumar, S.; Balakrishnan, N.; Samiyappan, R. Biotechnological and molecular approaches in the management of non-insect pests of crop plants. In Integrated Pest Management; Elsevier: Amsterdam, The Netherlands, 2014; pp. 337–369. [Google Scholar]
- İnak, E.; Alpkent, Y.N.; Çobanoğlu, S.; Dermauw, W.; Van Leeuwen, T. Resistance incidence and presence of resistance mutations in populations of Tetranychus urticae from vegetable crops in Turkey. Exp. Appl. Acarol. 2019, 78, 343–360. [Google Scholar] [CrossRef]
- Demaeght, P.; Osborne, E.J.; Odman-Naresh, J.; Grbić, M.; Nauen, R.; Merzendorfer, H.; Clark, R.M.; Van Leeuwen, T. High resolution genetic mapping uncovers chitin synthase-1 as the target-site of the structurally diverse mite growth inhibitors clofentezine, hexythiazox and etoxazole in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 51, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.B.S.; Andrade, F.H.N.; Rodrigues, A.R.S.; Siqueira, H.A.A.; Gondim, M.G.C. Resistance in field populations of Tetranychus urticae to acaricides and characterization of the inheritance of abamectin resistance. Crop Prot. 2015, 67, 77–83. [Google Scholar] [CrossRef]
- Snyder, M.J.; Glendinning, J.I. Causal connection between detoxification enzyme activity and consumption of a toxic plant compound. J. Comp. Physiol. A 1996, 179, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, N.; Nauen, R. Biochemical markers linked to abamectin resistance in Tetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 2002, 72, 111–121. [Google Scholar] [CrossRef]
- Pavlidi, N.; Tseliou, V.; Riga, M.; Nauen, R.; Van Leeuwen, T.; Labrou, N.E.; Vontas, J. Functional characterization of glutathione S-transferases associated with insecticide resistance in Tetranychus urticae. Pestic. Biochem. Physiol. 2015, 121, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Pavlidi, N.; Khalighi, M.; Myridakis, A.; Dermauw, W.; Wybouw, N.; Tsakireli, D.; Stephanou, E.G.; Labrou, N.E.; Vontas, J.; Van Leeuwen, T. A glutathione-S-transferase (TuGSTd05) associated with acaricide resistance in Tetranychus urticae directly metabolizes the complex II inhibitor cyflumetofen. Insect Biochem. Mol. Biol. 2017, 80, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Merzendorfer, H. ABC transporters and their role in protecting insects from pesticides and their metabolites. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–72. [Google Scholar]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbic, M.; Clark, R.M.; Feyereisen, R.; Van Leeuwen, T. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Natl. Acad. Sci. 2012, 110, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Dermauw, W.; Grbic, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2012, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Demaeght, P.; Dermauw, W.; Tsakireli, D.; Khajehali, J.; Nauen, R.; Tirry, L.; Vontas, J.; Lümmen, P.; Van Leeuwen, T. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae: CYP392E10 metabolizes spirodiclofen, but not its corresponding enol. Insect Biochem. Mol. Biol. 2013, 43, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Choi, J.Y.; Je, Y.H.; Lee, S.H. The overexpression of acetylcholinesterase compensates for the reduced catalytic activity caused by resistance-conferring mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2012, 42, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Flood, J.; Day, R. Managing risks from pests in global commodity networks–policy perspectives. Food Secur. 2016, 8, 89–101. [Google Scholar] [CrossRef]
- Pimentel, D. Environmental and economic costs of the application of pesticides primarily in the united states. Environ. Dev. Sustain. 2005, 7, 229–252. [Google Scholar] [CrossRef]
- Price, J.F.; Legard, D.E.; Chandler, C.K. Two-spotted spider mite resistance to abamectin miticide on strawberry and strategies for resistance management. Acta Hortic. 2002, 683–685. [Google Scholar] [CrossRef]
- García-Marí, F.; Gonzalez-Zamora, J.E. Biological control of Tetranychus urticae (Acari: Tetranychidae) with naturally occurring predators in strawberry plantings in Valencia, Spain. Exp. Appl. Acarol. 1999, 23, 487–495. [Google Scholar] [CrossRef]
- Leite, L.G.; Smith, L.; Moraes, G.J.; Roberts, D.W. In vitro production of hyphal bodies of the mite pathogenic fungus Neozygites floridana. Mycologia 2000, 92, 201–207. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A review of the major biological approaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Monsreal-Ceballos, R.J.; Ruiz-Sánchez, E.; Sánchez-Borja, M.; Ballina-Gómez, H.S.; González-Moreno, A.; Reyes-Ramírez, A. Effects of commercial botanical insecticides in Tamarixia radiata, an ectoparasitoid of Diaphorina citri. Ecosistemas y Recur. Agropecu. 2017, 4, 589. [Google Scholar] [CrossRef]
- Duso, C.; Malagnini, V.; Pozzebon, A.; Castagnoli, M.; Liguori, M.; Simoni, S. Comparative toxicity of botanical and reduced-risk insecticides to Mediterranean populations of Tetranychus urticae and Phytoseiulus persimilis (Acari Tetranychidae, Phytoseiidae). Biol. Control 2008, 47, 16–21. [Google Scholar] [CrossRef]
- Spollen, K.M.; Isman, M.B. Acute and sublethal effects of a Neem insecticide on the commercial biological control agents Phytoseiulus persimilis and Amblyseius cucumeris (Acari: Phytoseiidae) and Aphidoletes aphidimyza (Diptera: Cecidomyiidae). J. Econ. Entomol. 1996, 89, 1379–1386. [Google Scholar] [CrossRef]
- Bernardi, D.; Botton, M.; da Cunha, U.S.; Bernardi, O.; Malausa, T.; Garcia, M.S.; Nava, D.E. Effects of azadirachtin on Tetranychus urticae (Acari: Tetranychidae) and its compatibility with predatory mites (Acari: Phytoseiidae) on strawberry. Pest Manag. Sci. 2013, 69, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Brito, H.M.; Gondim, M.G.C.G., Jr.; de Oliveira, J.V.; da Câmara, C.A.G. Toxicidade de formulações de nim (Azadirachta indica A. Juss.) ao ácaro-rajado e a Euseius alatus De Leon e Phytoseiulus macropilis (Banks) (Acari: Phytoseiidae). Neotrop. Entomol. 2006, 35, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Yanar, D. Side effects of different doses of azadirachtin on predatory mite Metaseiulus occidentalis (Nesbitt) (acari: Phytoseiidae) under laboratory conditions. Appl. Ecol. Environ. Res. 2019, 17, 3433–3440. [Google Scholar] [CrossRef]
- El-Sharabasy, H.M. Acaricidal activities of Artemisia judaica L. extracts against Tetranychus urticae Koch and its predator Phytoseiulus persimilis Athias-Henriot (Tetranychidae: Phytoseiidae). J. Biopestic. 2010, 3, 514–519. [Google Scholar]
- Lima, D.B.; Melo, J.W.S.; Guedes, N.M.P.; Gontijo, L.M.; Guedes, R.N.C.; Gondim, M.G.C., Jr. Bioinsecticide-predator interactions: Azadirachtin behavioral and reproductive impairment of the coconut mite predator Neoseiulus baraki. PLoS ONE 2015, 10, e0118343. [Google Scholar] [CrossRef]
- Chiasson, H.; Bostanian, N.J.; Vincent, C. Acaricidal properties of a Chenopodium-Based Botanical. J. Econ. Entomol. 2004, 97, 1373–1377. [Google Scholar] [CrossRef]
- Choi, W.-I.; Lee, S.-G.; Park, H.-M.; Ahn, Y.-J. Toxicity of plant essential oils to Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae). J. Econ. Entomol. 2004, 97, 553–558. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Lognay, G.; Heuskin, S.; Mailleux, A.C.; Hance, T. Chemical composition and acaricidal properties of Deverra scoparia essential oil (Araliales: Apiaceae) and blends of its major constituents against Tetranychus urticae (Acari: Tetranychidae). J. Econ. Entomol. 2011, 104, 1220–1228. [Google Scholar] [CrossRef]
- Pontes, W.J.T.; de Oliveira, J.C.S.; da Câmara, C.A.G.; Júnior, M.G.C.G.; de Oliveira, J.V.; Schwartz, M.O.E. Atividade acaricida dos óleos essencias de folhas e frutos de Xylopia sericea sobre o ácaro rajado (Tetranychus urticae Koch). Quim. Nova 2007, 30, 838. [Google Scholar] [CrossRef]
- Chiasson, H.; Bélanger, A.; Bostanian, N.; Vincent, C.; Poliquin, A. Acaricidal properties of Artemisia absinthium and Tanacetum vulgare (Asteraceae) essential oils obtained by three methods of extraction. J. Econ. Entomol. 2001, 94, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.; Grissa, K.L.; Mailleux, A.C.; Heuskin, S.; Lognay, G.; Hance, T. Acaricidal activities of Santolina africana and Hertia cheirifolia essential oils against the two-spotted spider mite (Tetranychus urticae). Pest Manag. Sci. 2012, 68, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Rassem, H.H.A.; Nour, A.H.; Yunus, R.M. Techniques for extraction of essential oils from plants: A review. Aust. J. Basic Appl. Sci. 2016, 10, 117–127. [Google Scholar]
- Kabir, K.H.; Chapman, R.B.; Penman, D.R. Miticide bioassays with spider mites (Acari: Tetranychidae): Effect of test method, exposure period and mortality criterion on the precision of response estimates. Exp. Appl. Acarol. 1993, 17, 695–708. [Google Scholar] [CrossRef]
- Walker, W.F.; Boswell, A.L.; Smith, F.F. Resistance of spider mites to acaricides: Comparison of slide dip and leaf dip methods. J. Econ. Entomol. 1973, 66, 549–550. [Google Scholar] [CrossRef]
- Voss, G. Ein neues akarizid-austestungsverfahren für spinnmilben. Anz. Schädlingskd 1961, 34, 76–77. [Google Scholar] [CrossRef]
- Dittrich, V. A comparative study of toxicological test methods on a population of the two-spotted spider mite (Tetranychus telarius). J. Econ. Entomol. 1962, 55, 644–648. [Google Scholar] [CrossRef]
- Shi, G.L.; Zhao, L.L.; Liu, S.Q.; Cao, H.; Clarke, S.R.; Sun, J.H. Acaricidal activities of extracts of Kochia scoparia against Tetranychus urticae, Tetranychus cinnabarinus, and Tetranychus viennensis (Acari: Tetranychidae). J. Econ. Entomol. 2006, 99, 858–863. [Google Scholar] [CrossRef]
- Laborda, R.; Manzano, I.; Gamón, M.; Gavidia, I.; Pérez-Bermúdez, P.; Boluda, R. Effects of Rosmarinus officinalis and Salvia officinalis essential oils on Tetranychus urticae Koch (Acari: Tetranychidae). Ind. Crops Prod. 2013, 48, 106–110. [Google Scholar] [CrossRef]
- Akyazi, R.; Soysal, M.; Hassan, E. Toxic and repellent effects of Prunus laurocerasus L. (Rosaceae) extracts against Tetranychus urticae Koch (Acari: Tetranychidae). Türk. Entomol. Derg. 2015, 39, 367–380. [Google Scholar] [CrossRef]
- Mahmoud, N.F.; Badawy, M.E.I.; Marei, A.E.-S.M.; Abdelgaleil, S.A.M. Acaricidal and antiacetylcholinesterase activities of essential oils from six plants growing in Egypt. Int. J. Acarol. 2019, 45, 245–251. [Google Scholar] [CrossRef]
- Laborda, R.; Manzano, I.; Gamon, M.; Gavidia, I.; Boluda, R.; Perez-Bermudez, P. Spike lavender essential oil reduces the survival rate and fecundity of two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). J. Agric. Sci. Technol. 2018, 20, 1013–1023. [Google Scholar]
- Eldoksch, H.A.; Ayad, F.A.; El-Sebae, A.-K.H. Acaricidal activity of plant extracts and their main terpenoids on the two-spotted spider mite Tetranychus Urticae (Acari: Tetranychidae). Alexandria Sci. Exch. J. 2009, 30, 344–349. [Google Scholar]
- Helle, W.; Overmeer, W. Toxicological test methods. In Spider Mites. Their Biology, Natural Enemies and Control.; Helle, W., Sabelis, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1, pp. 391–395. [Google Scholar]
- Kabir, K.H.; Chapman, R.B. Operational and biological factors influencing responses of spider mites (acari: Tetranychidae) to propargite by using the petri dish-potter tower method. J. Econ. Entomol. 1997, 90, 272–277. [Google Scholar] [CrossRef]
- Yanar, D.; Kadıoğlu, I.; Gökçe, A. Acaricidal effects of different plant parts extracts on two-spotted spider mite (Tetranychus urticae Koch). Afr. J. Biotechnol. 2011, 10, 11745–11750. [Google Scholar]
- Numa, S.; Rodríguez, L.; Rodríguez, D.; Coy-Barrera, E. Susceptibility of Tetranychus urticae Koch to an ethanol extract of Cnidoscolus aconitifolius leaves under laboratory conditions. Springerplus 2015, 4, 338. [Google Scholar] [CrossRef]
- Potter, C. An improved laboratory apparatus for applying direct sprays and surface films, with data on the electrostatic charge on atomized spray fluids. Ann. Appl. Biol. 1952, 39, 1–28. [Google Scholar] [CrossRef]
- Bostanian, N.J.; Beudjekian, S.; McGregor, E.; Racette, G. A modified excised leaf disc method to estimate the toxicity of slow- and fast-acting reduced-risk acaricides to mites. J. Econ. Entomol. 2009, 102, 2084–2089. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Jalali-Sendi, J.; Razmjou, J. Toxicity and phytochemical profile of essential oil from Iranian Achillea mellifolium L. against Tetranychus urticae Koch (Acari: Tetranychidae). Toxin Rev. 2016, 35, 24–28. [Google Scholar] [CrossRef]
- Pavela, R. Acaricidal properties of extracts and major furanochromenes from the seeds of Ammi visnaga Linn. against Tetranychus urticae Koch. Ind. Crops Prod. 2015, 67, 108–113. [Google Scholar] [CrossRef]
- Pavela, R. Extract from the roots of Saponaria officinalis as a potential acaricide against Tetranychus urticae. J. Pest Sci. 2017, 90, 683–692. [Google Scholar] [CrossRef]
- da Camara, C.A.G.; Akhtar, Y.; Isman, M.B.; Seffrin, R.C.; Born, F.S. Repellent activity of essential oils from two species of Citrus against Tetranychus urticae in the laboratory and greenhouse. Crop Prot. 2015, 74, 110–115. [Google Scholar] [CrossRef]
- Han, J.; Choi, B.R.; Lee, S.G.; Il Kim, S.; Ahn, Y.J. Toxicity of plant essential oils to acaricide-susceptible and -resistant Tetranychus urticae (Acari: Tetranychidae) and Neoseiulus californicus (Acari: Phytoseiidae). J. Econ. Entomol. 2010, 103, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Barua, C.C.; Talukdar, A.; Begum, S.A.; Lahon, L.C.; Sarma, D.K.; Pathak, D.C.; Borah, P. Antinociceptive activity of methanolic extract of leaves of Achyranthes aspera Linn. (Amaranthaceae) in animal models of nociception. Indian J. Exp. Biol. 2010, 48, 817–821. [Google Scholar] [PubMed]
- Monzote, L.; Stamberg, W.; Staniek, K.; Gille, L. Toxic effects of carvacrol, caryophyllene oxide, and ascaridole from essential oil of Chenopodium ambrosioides on mitochondria. Toxicol. Appl. Pharmacol. 2009, 240, 337–347. [Google Scholar] [CrossRef]
- Hiremath, I.G.; Ahn, Y.J.; Kim, S.I.; Choi, B.R.; Cho, J.R. Insecticidal and acaricidal activities of african plant extracts against the brown planthopper and two-spotted spider mite. Korean J. Appl. Entomol. 1995, 34, 200–205. [Google Scholar]
- Harder, M.J.; Tello, V.E.; Giliomee, J.H. The Acaricidal effect of ethanolic extracts of Chenopodium quinoa Willd. on Tetranychus urticae Koch (Acari: Tetranychidae). Afr. Entomol. 2016, 24, 50–60. [Google Scholar] [CrossRef]
- Renard-Nozaki, J.; Kim, T.; Imakura, Y.; Kihara, M.; Kobayashi, S. Effect of alkaloids isolated from Amaryllidaceae on Herpes simplex virus. Res. Virol. 1989, 140, 115–128. [Google Scholar] [CrossRef]
- Weniger, B.; Italiano, L.; Beck, J.-P.; Bastida, J.; Bergoñon, S.; Codina, C.; Lobstein, A.; Anton, R. Cytotoxic activity of Amaryllidaceae alkaloids. Planta Med. 1995, 61, 77–79. [Google Scholar] [CrossRef]
- Ho, S.H.; Koh, L.; Ma, Y.; Huang, Y.; Sim, K.Y. The oil of garlic, Allium sativum L. (Amaryllidaceae), as a potential grain protectant against Tribolium castaneum (Herbst) and Sitophilus zeamais Motsch. Postharvest Biol. Technol. 1996, 9, 41–48. [Google Scholar] [CrossRef]
- Abbassy, M.A.; El-Gougary, O.A.; El-Hamady, S.; Sholo, M.A. Insecticidal, acaricidal and synergistic effects of soosan, Pancratium maritimum extracts and constituents. J. Egypt. Soc. Parasitol. 1998, 28, 197–205. [Google Scholar] [PubMed]
- Attia, S.; Grissa, K.L.; Mailleux, A.C.; Lognay, G.; Heuskin, S.; Mayoufi, S.; Hance, T. Effective concentrations of garlic distillate (Allium sativum) for the control of Tetranychus urticae (Tetranychidae). J. Appl. Entomol. 2012, 136, 302–312. [Google Scholar] [CrossRef]
- Geng, S.; Chen, H.; Zhang, J.; Tu, H. Bioactivity of garlic-straw extracts against the spider mites, Tetranychus urticae and T. viennensis. J. Agric. Urban Entomol. 2014, 30, 38–48. [Google Scholar] [CrossRef]
- Rabelo, S.V.; de Sousa Siqueira Quintans, J.; Costa, E.V.; da Silva Almeida, J.R.G.; Júnior, L.J.Q. Annona species (Annonaceae) oils. In Essential Oils in Food Preservation, Flavor and Safety; Elsevier: Amsterdam, The Netherlands, 2016; pp. 221–229. [Google Scholar]
- Ohsawa, K.; Atsuzawa, S.; Mitsui, T.; Yamamoto, I. Isolation and insecticidal activity of three acetogenins from seeds of pond apple, Annona glabra L. J. Pestic. Sci. 1991, 16, 93–96. [Google Scholar] [CrossRef]
- Tunçtürk, M.; Özgökçe, F. Chemical composition of some Apiaceae plants commonly used inherby cheese in Eastern Anatolia. Turkish J. Agric. For. 2015, 39, 55–62. [Google Scholar] [CrossRef]
- Benelli, G.; Flamini, G.; Fiore, G.; Cioni, P.L.; Conti, B. Larvicidal and repellent activity of the essential oil of Coriandrum sativum L. (Apiaceae) fruits against the filariasis vector Aedes albopictus Skuse (Diptera: Culicidae). Parasitol. Res. 2013, 112, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Tsolakis, H.; Ragusa, S. Effects of a mixture of vegetable and essential oils and fatty acid potassium salts on Tetranychus urticae and Phytoseiulus persimilis. Ecotoxicol. Environ. Saf. 2008, 70, 276–282. [Google Scholar] [CrossRef]
- Amizadeh, M.; Hejazi, M.J.; Saryazdi, G.A. Fumigant toxicity of some essential oils on Tetranychus urticae (Acari: Tetranychidae). Int. J. Acarol. 2013, 39, 285–289. [Google Scholar] [CrossRef]
- Kheradmand, K.; Beynaghi, S.; Asgari, S.; Garjan, A.S. Toxicity and repellency effects of three plant essential oils against two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). J. Agr. Sci. Tech. 2015, 17, 1223–1232. [Google Scholar]
- Fatemikia, S.; Abbasipour, H.; Saeedizadeh, A. Phytochemical and acaricidal study of the Galbanum, Ferula gumosa Boiss. (Apiaceae) essential oil against Tetranychus urticae Koch (Tetranychidae). J. Essent. Oil Bear. Plants 2017, 20, 185–195. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Canale, A.; Nicoletti, M.; Petrelli, R.; Cappellacci, L.; Galassi, R.; Maggi, F. Isofuranodiene and germacrone from Smyrnium olusatrum essential oil as acaricides and oviposition inhibitors against Tetranychus urticae: Impact of chemical stabilization of isofuranodiene by interaction with silver triflate. J. Pest Sci. 2017, 90, 693–699. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Ghrabi, Z.G.; Mailleux, A.C.; Lognay, G.; Hance, T. Acaricidal activity of 31 essential oils extracted from plants collected in Tunisia. J. Essent. Oil Res. 2012, 24, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Afify, A.E.-M.M.R.; Ali, F.S.; Turky, A.F. Control of Tetranychus urticae Koch by extracts of three essential oils of chamomile, marjoram and Eucalyptus. Asian Pac. J. Trop. Biomed. 2012, 2, 24–30. [Google Scholar] [CrossRef]
- Aslan, I.; Kordali, S.; Çalmaşur, Ö. Toxicity of the vapours of Artemisia absinthium essential oils to Tetranychus urticae Koch and Bemisia tabasi (Genn.). Fresenius Environ. Bull. 2005, 14, 415–417. [Google Scholar]
- Derbalah, A.S.; Keratrum, A.Y.; El-Dewy, M.E.; El-Shamy, E.H. Efficacy of some insecticides and plant extracts against Tetranychus urticae under laboratory conditions. Egypt. J. Plant Prot. Res. 2013, 1, 47–70. [Google Scholar]
- Pavela, R.; Dall’acqua, S.; Sut, S.; Baldan, V.; Kamte, S.L.N.; Nya, P.C.B.; Cappellacci, L.; Petrelli, R.; Nicoletti, M.; Canale, A.; et al. Oviposition inhibitory activity of the Mexican sunflower Tithonia diversifolia (Asteraceae) polar extracts against the two-spotted spider mite Tetranychus urticae (Tetranychidae). Physiol. Mol. Plant Pathol. 2018, 101, 85–92. [Google Scholar] [CrossRef]
- Sut, S.; Pavela, R.; Kolarčik, V.; Cappellacci, L.; Petrelli, R.; Maggi, F.; Dall’Acqua, S.; Benelli, G. Identification of Onosma visianii roots extract and purified shikonin derivatives as potential acaricidal agents against Tetranychus urticae. Molecules 2017, 22, E1002. [Google Scholar] [CrossRef]
- Carretero, M.E.; López-Pérez, J.L.; Abad, M.J.; Bermejo, P.; Tillet, S.; Israel, A.; Noguera-P, B. Preliminary study of the anti-inflammatory activity of hexane extract and fractions from Bursera simaruba (Linneo) Sarg. (Burseraceae) leaves. J. Ethnopharmacol. 2008, 116, 11–15. [Google Scholar] [CrossRef]
- Rama, K.S.; Chandrasekar, R.M.; Rani, S.; Pullaiah, T. Bioactive principles and biological properties of essential oils of burseraceae: A review. J. Pharmacogn. Phytochem. 2016, 5, 247–258. [Google Scholar]
- Baratta, M.T.; Dorman, H.J.D.; Deans, S.G.; Figueiredo, A.C.; Barroso, J.G.; Ruberto, G. Antimicrobial and antioxidant properties of some commercial essential oils. Flavour Fragr. J. 1998, 13, 235–244. [Google Scholar] [CrossRef]
- Pontes, W.J.T.; de Oliveira, J.C.S.; da Camara, C.A.G.; Lopes, A.C.H.R.; Gondim, M.G.C.; de Oliveira, J.V.; Schwartz, M.O.E. Composition and acaricidal activity of the resin’s essential oil of Protium bahianum Daly against two spotted spider mite (Tetranychus Urticae). J. Essent. Oil Res. 2007, 19, 379–383. [Google Scholar] [CrossRef]
- Górski, R.; Sobieralski, K.; Siwulski, M. The effect of hemp essential oil on mortality Aulacorthum solani Kalt. and Tetranychus urticae Koch. Ecol. Chem. Eng. S. 2016, 23, 505–511. [Google Scholar] [CrossRef]
- Chermenskaya, T.D.; Stepanycheva, E.A.; Shchenikova, A.V.; Chakaeva, A.S. Insectoacaricidal and deterrent activities of extracts of Kyrgyzstan plants against three agricultural pests. Ind. Crops Prod. 2010, 32, 157–163. [Google Scholar] [CrossRef]
- Le Dang, Q.; Choi, Y.H.; Choi, G.J.; Jang, K.S.; Park, M.S.; Park, N.-J.; Lim, C.H.; Kim, H.; Ngoc, L.H.; Kim, J.-C. Pesticidal activity of ingenane diterpenes isolated from Euphorbia kansui against Nilaparvata lugens and Tetranychus urticae. J. Asia Pac. Entomol. 2010, 13, 51–54. [Google Scholar] [CrossRef]
- Islam, T.; Biswas, M.J.H.; Howlader, M.T.H.; Ullah, M.S. Laboratory evaluation of Beauveria bassiana, some plant oils and insect growth regulators against two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Persian, J. Acarol. 2017, 6, 203–211. [Google Scholar]
- Shah, G.; Shri, R.; Panchal, V.; Sharma, N.; Singh, B.; Mann, A.S. Scientific basis for the therapeutic use of Cymbopogon citratus, stapf (Lemon grass). J. Adv. Pharm. Technol. Res. 2011, 2, 3. [Google Scholar] [CrossRef]
- Rasikari, H.L.; Leach, D.N.; Waterman, P.G.; Spooner-hart, R.N.; Basta, A.H.; Banbury, L.K.; Forster, P.I. Acaricidal and cytotoxic activities of extracts from selected genera of Australian Lamiaceae. J. Econ. Entomol. 2005, 98, 1259–1266. [Google Scholar] [CrossRef]
- Miresmailli, S.; Bradbury, R.; Isman, M.B. Comparative toxicity of Rosmarinus officinalis L. essential oil and blends of its major constituents against Tetranychus urticae Koch (Acari: Tetranychidae) on two different host plants. Pest Manag. Sci. 2006, 62, 366–371. [Google Scholar] [CrossRef]
- Çalmaşur, Ö.; Aslan, İ.; Şahin, F. Insecticidal and acaricidal effect of three Lamiaceae plant essential oils against Tetranychus urticae Koch and Bemisia tabaci Genn. Ind. Crops Prod. 2006, 23, 140–146. [Google Scholar] [CrossRef]
- Carpinella, M.C.; Defago, M.T.; Valladares, G.; Palacios, S.M. Antifeedant and insecticide properties of a limonoid Melia azedarach (Meliaceae) with potential use for pest management. J. Agric. Food Chem. 2003, 51, 369–374. [Google Scholar] [CrossRef]
- Ismail, S. Selectivity and joint action of Melia azedarach L. fruit extracts with certain acaricides to Tetranychus urticae Koch and Stethorus gilvifrons Mulsant. Ann. Agric. Sci. 1997, 35, 605–618. [Google Scholar]
- Afify, A.E.-M.M.R.; El-Beltagi, H.S.; Fayed, S.A.; Shalaby, E.A. Acaricidal activity of different extracts from Syzygium cumini L. Skeels (Pomposia) against Tetranychus urticae Koch. Asian Pac. J. Trop. Biomed. 2011, 1, 359–364. [Google Scholar] [CrossRef]
- Chaveerach, A.; Mokkamul, P.; Sudmoon, R.; Tanee, T. Ethnobotany of the genus Piper (Piperaceae) in Thailand. Ethnobot. Res. Appl. 2006, 4, 223–231. [Google Scholar] [CrossRef]
- Araújo, M.J.C.; Câmara, C.A.G.; Born, F.S.; Moraes, M.M.; Badji, C.A. Acaricidal activity and repellency of essential oil from Piper aduncum and its components against Tetranychus urticae. Exp. Appl. Acarol. 2012, 57, 139–155. [Google Scholar] [CrossRef]
- Tewary, D.K.; Bhardwaj, A.; Shanker, A. Pesticidal activities in five medicinal plants collected from mid hills of western Himalayas. Ind. Crops Prod. 2005, 22, 241–247. [Google Scholar] [CrossRef]
- Roh, H.S.; Lim, E.G.; Kim, J.; Park, C.G. Acaricidal and oviposition deterring effects of santalol identified in sandalwood oil against two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). J. Pest Sci. 2011, 84, 495–501. [Google Scholar] [CrossRef]
- Khambay, B.P.S.; Batty, D.; Cahill, M.; Denholm, I.; Mead-Briggs, M.; Vinall, S.; Niemeyer, H.M.; Simmonds, M.S.J. Isolation, Characterization, and biological activity of naphthoquinones from Calceolaria andina L. J. Agric. Food Chem. 1999, 47, 770–775. [Google Scholar] [CrossRef]
- Latif, Z.; Craven, L.; Hartley, T.G.; Kemp, B.R.; Potter, J.; Rice, M.J.; Waigh, R.D.; Waterman, P.G. An insecticidal quassinoid from the new Australian species Quassia sp. aff. bidwillii. Biochem. Syst. Ecol. 2000, 28, 183–184. [Google Scholar] [CrossRef]
- Snyder, J.C.; Guo, Z.; Thacker, R.; Goodman, J.P.; Pyrek, J.S. 2,3-dihydrofarnesoic acid, a unique terpene from trichomes of Lycopersicon hirsutum, repels spider mites. J. Chem. Ecol. 1993, 19, 2981–2997. [Google Scholar] [CrossRef]
- Antonious, G.F.; Meyer, J.E.; Snyder, J.C. Toxicity and repellency of hot pepper extracts to spider mite, Tetranychus urticae Koch. J. Environ. Sci. Heal. Part B 2006, 41, 1383–1391. [Google Scholar] [CrossRef]
- Cavalcanti, S.C.H.; Dos, S.; Niculau, E.; Blank, A.F.; Câmara, C.A.G.; Araújo, I.N.; Alves, P.B. Composition and acaricidal activity of Lippia sidoides essential oil against two-spotted spider mite (Tetranychus urticae Koch). Bioresour. Technol. 2010, 101, 829–832. [Google Scholar] [CrossRef]
- Lee, S.; Tsao, R.; Peterson, C.; Coats, J.R. Insecticidal activity of monoterpenoids to western corn rootworm (Coleoptera: Chrysomelidae), two-spotted spider mite (Acari: Tetranychidae), and house fly (Diptera: Muscidae). J. Econ. Entomol. 1997, 90, 883–892. [Google Scholar] [CrossRef]
- Martínez-Villar, E.; Sáenz-De-Cabezón, F.J.; Moreno-Grijalba, F.; Marco, V.; Pérez-Moreno, I. Effects of azadirachtin on the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2005, 35, 215–222. [Google Scholar] [CrossRef]
- Han, J.; Kim, S.-I.; Choi, B.-R.; Lee, S.-G.; Ahn, Y.-J. Fumigant toxicity of lemon eucalyptus oil constituents to acaricide-susceptible and acaricide-resistant Tetranychus urticae. Pest Manag. Sci. 2011, 67, 1583–1588. [Google Scholar] [CrossRef]
- Akhtar, Y.; Isman, M.B.; Lee, C.-H.; Lee, S.-G.; Lee, H.-S. Toxicity of quinones against two-spotted spider mite and three species of aphids in laboratory and greenhouse conditions. Ind. Crops Prod. 2012, 37, 536–541. [Google Scholar] [CrossRef]
- Marčić, D.; Međo, I. Acaricidal activity and sublethal effects of an oxymatrine-based biopesticide on two-spotted spider mite (Acari: Tetranychidae). Exp. Appl. Acarol. 2014, 64, 375–391. [Google Scholar] [CrossRef]
- Marčić, D.; Međo, I. Sublethal effects of azadirachtin-A (NeemAzal-T/S) on Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol. 2015, 30, 25. [Google Scholar]
- Numa, S.; Rodríguez-Coy, L.; Rodríguez, D.; Coy-Barrera, E. Laboratory screening of six botanicals for acaricidal activity against two-spotted spider mite, Tetranychus urticae (Koch). Biopestic. Int. 2017, 13, 13–20. [Google Scholar]
- Mostafa, M.E.; Alshamy, M.M.; Abdelmonem, A.; Abdel-Mogib, M. Acaricidal activity of Chrozophora oblongifolia on the two spotted spider mite, Tetranychus urticae Koch. J. Entomol. Nematol. 2017, 9, 23–28. [Google Scholar]
- Numa, S.; Rodríguez-Coy, L.; Rodríguez, D.; Coy-Barrera, E. Examination of the acaricidal effect of a set of colombian native plants-derived extracts against Tetranychus urticae Koch under laboratory conditions. J. Biopestic. 2018, 11, 30–37. [Google Scholar]
- Reddy, S.G.E.; Dolma, S.K. Acaricidal activities of essential oils against two-spotted spider mite, Tetranychus urticae Koch. Toxin Rev. 2017, 37, 62–66. [Google Scholar] [CrossRef]
- Erdogan, P.; Yilmaz, B.S. Acaricidal activity of extracts of Juglans regia L. on Tetranychus urticae Koch (Acari: Tetranychidae). J. Food Sci. Eng. 2017, 7, 202–208. [Google Scholar]
- Chrysargyris, A.; Laoutari, S.; Litskas, V.D.; Stavrinides, M.C.; Tzortzakis, N. Effects of water stress on lavender and sage biomass production, essential oil composition and biocidal properties against Tetranychus urticae (Koch). Sci. Hortic. Amst. 2016, 213, 96–103. [Google Scholar] [CrossRef]
- Pavela, R.; Stepanycheva, E.; Shchenikova, A.; Chermenskaya, T.; Petrova, M. Essential oils as prospective fumigants against Tetranychus urticae Koch. Ind. Crops Prod. 2016, 94, 755–761. [Google Scholar] [CrossRef]
- Zevedo, S.G.; Mar, J.M.; da Silva, L.S.; França, L.P.; Machado, M.B.; Tadei, W.P.; Bezerra, J.D.A.; dos Santos, A.L.; Sanches, E.A.; Sanches, E.A. Bioactivity of Licaria puchury-major essential oil against Aedes aegypti, Tetranychus urticae and Cerataphis lataniae. Rec. Nat. Prod. 2018, 12, 229–238. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Sendi, J.J.; Maroufpoor, M.; Rahimi-Nasrabadi, M. Acaricidal potentials of the terpene-rich essential oils of two Iranian Eucalyptus species against Tetranychus urticae Koch. J. Oleo Sci. 2017, 66, 307–314. [Google Scholar] [CrossRef]
- Wagan, T.A.; Cai, W.; Hua, H. Repellency, toxicity, and anti-oviposition of essential oil of Gardenia jasminoides and its four major chemical components against whiteflies and mites. Sci. Rep. 2018, 8, 9375. [Google Scholar] [CrossRef]
- Numa, S.; Rodríguez-Coy, L.; Rodríguez, D.; Coy-Barrera, E. Effect of acaricidal activity of Solanum nigrum on Tetranychus urticae Koch under laboratory conditions. Afr. J. Biotechnol. 2015, 15, 363–369. [Google Scholar]
- Mar, J.M.; Silva, L.S.; Azevedo, S.G.; França, L.P.; Goes, A.F.; dos Santos, A.L.; Bezerra, J.D.A.; Rita de Cássia, S.N.; Machado, M.B.; Sanches, E.A. Lippia origanoides essential oil: An efficient alternative to control Aedes aegypti, Tetranychus urticae and Cerataphis lataniae. Ind. Crops Prod. 2018, 111, 292–297. [Google Scholar] [CrossRef]

| Acaricide | Population | N° | LC50 (mg/L) * | LC95 (mg/L) * | RR50 |
|---|---|---|---|---|---|
| Diafenthiuron | Petrolina II | 426 | 6.6 | 70 | 1 |
| Piracicaba | 484 | 10.7 | 105 | 1.6 | |
| Brejão | 340 | 4053 | 93,708 | 619 | |
| Bonito | 401 | 7732 | 133,440 | 1180 | |
| Milbemectin | Piracicaba | 484 | 0.6 | 8.3 | 1 |
| Petrolina II | 455 | 5.4 | 101 | 9.9 | |
| Bonito | 373 | 357 | 3726 | 650 | |
| Brejão | 380 | 384 | 2386 | 700 | |
| Fenpyroximate | Piracicaba | 315 | 22 | 341 | 1 |
| Petrolina II | 469 | 87 | 1929 | 4 | |
| Bonito | 378 | 3246 | 10,014 | 200 | |
| Brejão | 387 | 4343 | 16.234 | 150 | |
| Clorfenapyr | Piracicaba | 424 | 1.3 | 9.3 | 1 |
| Petrolina II | 481 | 2.8 | 20.1 | 2.2 | |
| Brejão | 477 | 735 | 4157 | 570 | |
| Bonito | 524 | 4652 | 94.598 | 3600 | |
| Spirodiclofen | Piracicaba | 401 | 16.4 | 1590 | 1 |
| Petrolina II | 547 | 37.5 | 370.870 | 2.3 | |
| Bonito | 538 | 6401 | 127.750 | 390 | |
| Brejão | 414 | 6586 | 56.390 | 400 | |
| Fenbutatin oxide | Piracicaba | 465 | 0.83 | 436 | 1 |
| Petrolina II | 538 | 1.72 | 1093 | 2.1 | |
| Bonito | 477 | 293 | 52.892 | 350 | |
| Brejão | 459 | 1705 | 197.990 | 2048 | |
| Propargite | Piracicaba | 472 | 6.5 | 28 | 1 |
| Petrolina II | 397 | 15 | 66 | 2.3 | |
| Bonito | 395 | 291 | 990 | 45 | |
| Brejão | 391 | 622 | 4410 | 96 | |
| Hexythiazox | Piracicaba | 416 | 2938 | 64.871 | 1 |
| Petrolina II | 404 | 4370 | 100.510 | 1.5 | |
| Brejão | 440 | 12,700 | 605.400 | 4.3 | |
| Bonito | 415 | 1384 | 381.630 | 4.7 | |
| Spiromesifen | Piracicaba | 418 | 373 | 18.404 | 1 |
| Petrolina II | 487 | 487 | 17.752 | 1 | |
| Brejão | 467 | 1388 | 42.781 | 3.7 | |
| Bonito | 470 | 3201 | 90.424 | 8.6 | |
| Abamectin | Petrolina II | 613 | 0.0011 | 0.033 | 1 |
| Piracicaba | 714 | 0.0084 | 0.066 | 8 | |
| Petrolina I | 676 | 0.036 | 0.205 | 34.4 | |
| Gravatá | 787 | 0.041 | 1.66 | 39.3 | |
| Goiânia | 584 | 1.79 | 27.9 | 1716 | |
| Brejão | 610 | 118 | 3000 | 113.532 | |
| Bonito | 693 | 326 | 3397 | 295.270 |
| Species | Taxonomy | Kind of Pest | Number of Active Ingredients | Cases of Resistance |
|---|---|---|---|---|
| Tetranychus urticae Koch | Acari: Tetranychidae | Crop | 93 | 389 |
| Plutella xylostella L. | Lepidoptera: Plutellidae | Crop | 81 | 437 |
| Myzus persicae Sulzer | Hemiptera: Aphididae | Crop | 73 | 320 |
| Leptinotarsa decemlineata Say | Coleoptera: Chrysomelidae | Crop | 51 | 188 |
| Musca domestica L. | Diptera: Muscidae | Urban | 53 | 266 |
| Blatella germanica L. | Blattodea: Blatellidae | Urban | 43 | 213 |
| Rhipicephalus microplus Canestrini | Acari: Ixodidae | Cattle | 43 | 158 |
| Helicoverpa armigera Hubner | Lepidoptera: Noctuidae | Crop | 43 | 639 |
| Bemisia tabaci Gennadius | Hemiptera: Aleyrodidae | Crop | 45 | 428 |
| Panonychus ulmi Koch | Acari: Tetranychidae | Crop | 42 | 181 |
| Varroa destructor Anderson y trueman | Acari: Varroidae | Bees parasite | 2 | 10 |
| Ixodes scapularis Say | Acari: Ixodidae | Cattle | 0 | 0 |
| Culex pipiens L. | Diptera: Culicidae | Disease vector | 36 | 161 |
| Culex quinquefasciatus Say | Diptera: Culicidae | Disease vector | 32 | 256 |
| Tribolium castaneum Herbst | Coleoptera: Tenebrionidae | Stored-grain pest | 32 | 113 |
| Aedes egypti egypti L. | Diptera: Culicidae | Disease vector | 24 | 267 |
| Spodoptera frugiperda Smith | Lepidoptera: Noctuidae | Crop | 16 | 25 |
| Pediculus humanus L. | Phthiraptera: Pediculidae | Disease vector | 9 | 59 |
| Anopheles gambiae Giles | Diptera: Culicidae | Disease vector | 3 | 39 |
| Manduca sexta L. | Lepidoptera: Sphingidae | Crop | 3 | 4 |
| Rhodnius prolixus Stal | Hemiptera: Reduviidae | Disease vector | 3 | 3 |
| Anopheles darlingi Root | Diptera: Culicidae | Disease vector | 1 | 2 |
| Linepithema humile Mayr | Hymenoptera: Formicidae | Urban | 2 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón, R.A.; Rodríguez, D.; Coy-Barrera, E. Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites. Plants 2019, 8, 272. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080272
Rincón RA, Rodríguez D, Coy-Barrera E. Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites. Plants. 2019; 8(8):272. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080272
Chicago/Turabian StyleRincón, Ricardo A., Daniel Rodríguez, and Ericsson Coy-Barrera. 2019. "Botanicals Against Tetranychus urticae Koch Under Laboratory Conditions: A Survey of Alternatives for Controlling Pest Mites" Plants 8, no. 8: 272. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080272