Eustress with H2O2 Facilitates Plant Growth by Improving Tolerance to Salt Stress in Two Wheat Cultivars

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
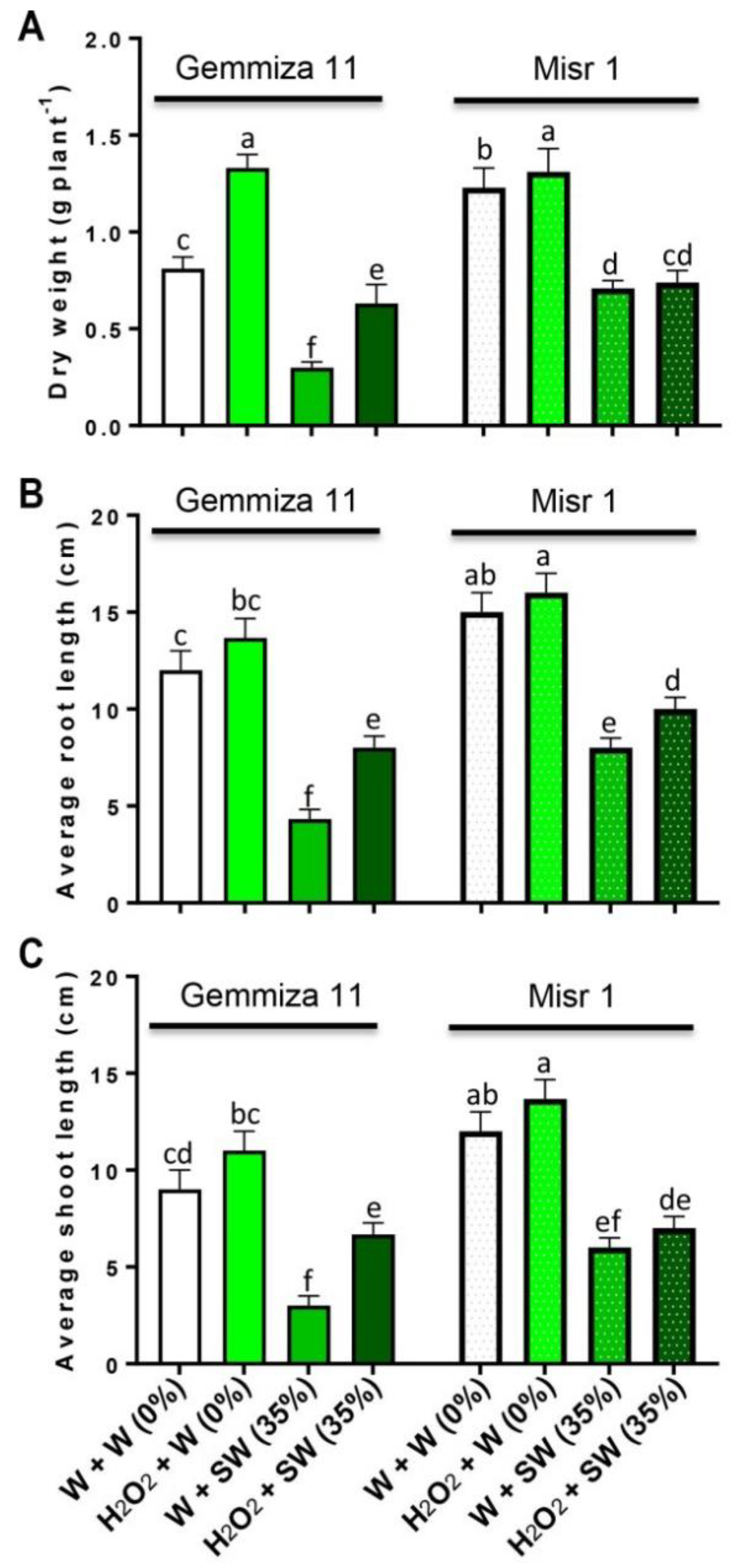
2.1. Seed Eustress with H2O2 Ameliorates Growth and Biomass Production in Wheat Plants
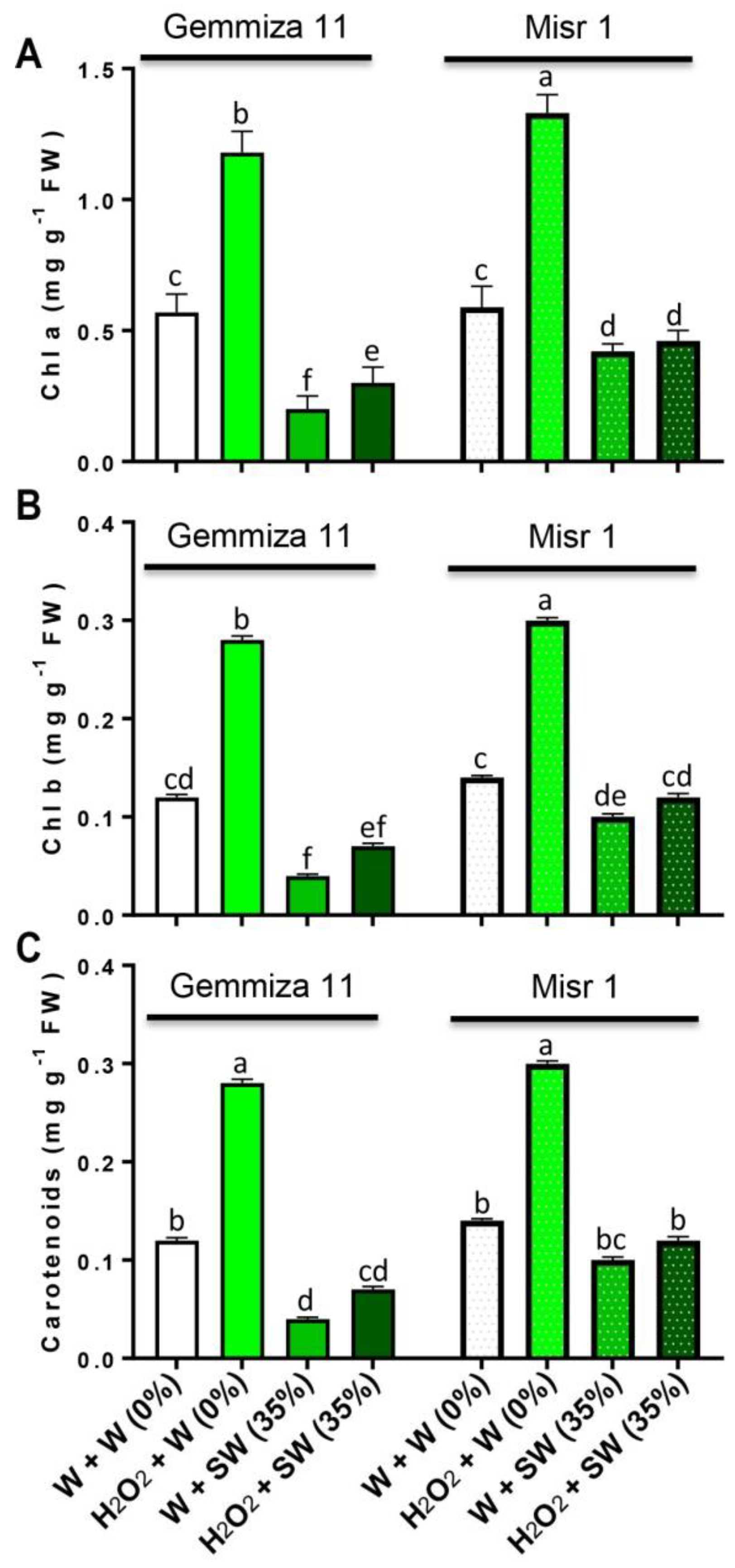
2.2. Seed Eustress with H2O2 Safeguards Photosynthetic Pigments from Seawater Stress
2.3. Seed Eustress with H2O2 Modulates the Content of Osmoprotectant (Proline) in Wheat Plants
2.4. Seed Eustress with H2O2 Regulates the Mineral Uptake in Seawater-Exposed Wheat Plants
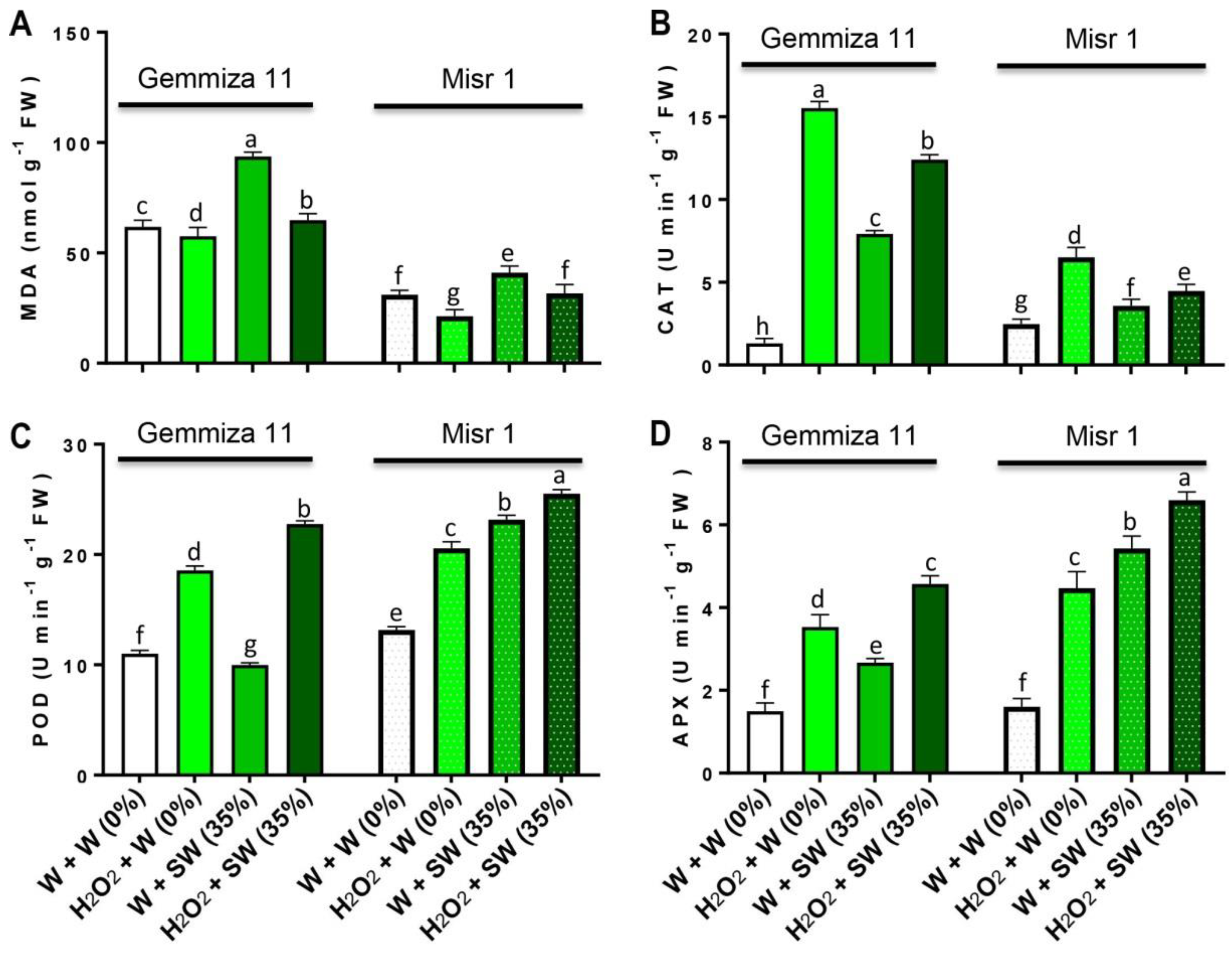
2.5. Seed Eustress with H2O2 Lessens Lipid Peroxidation through the Enhancement of Antioxidant Enzymes Activity in Wheat Plants
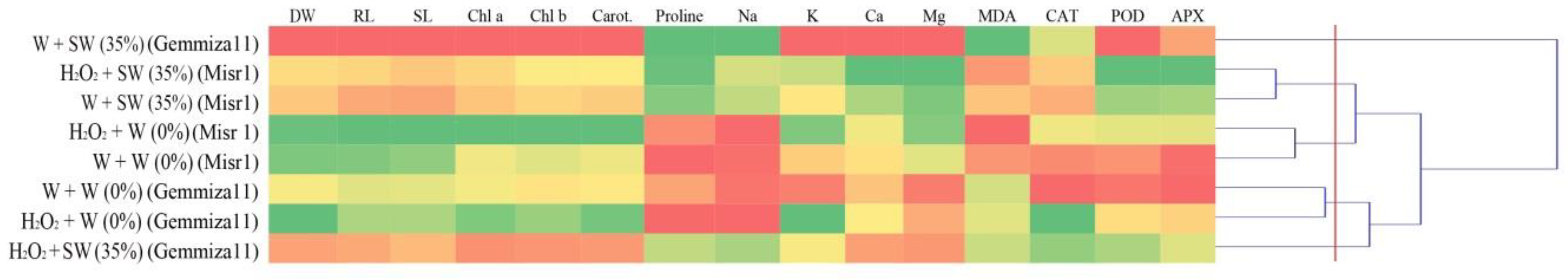
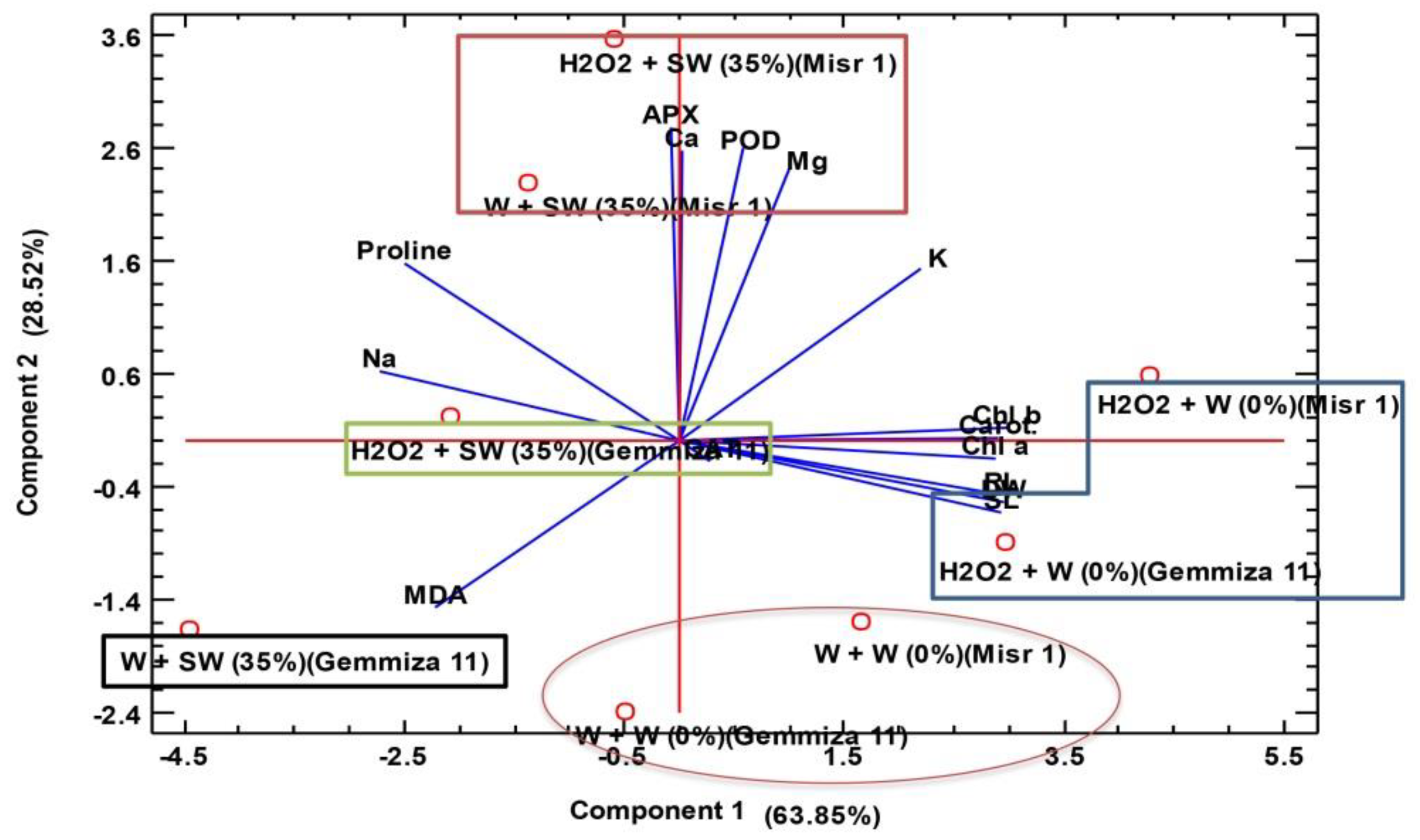
2.6. Hierarchical Clustering and Principle Component Analysis (PCA) Analysis
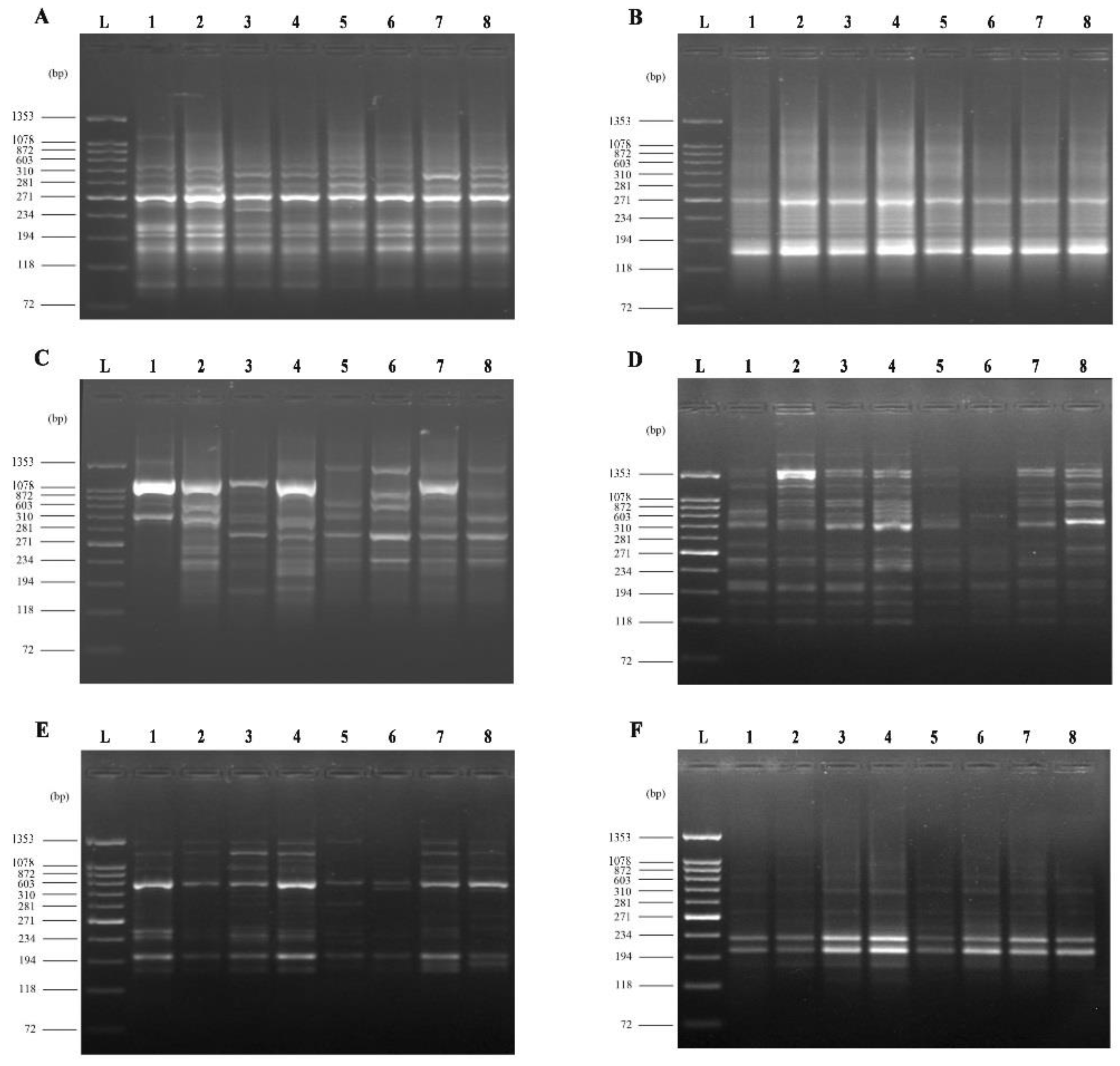
2.7. Molecular Analysis of Treatment
3. Discussion
4. Materials and Methods
4.1. Seed Selection and Treatments
4.2. Determination of Carotenoids and Chlorophyll Contents
4.3. Determination of Proline and Malondialdehyde Contents
4.4. Determination of Mineral Contents
4.5. Determination of Antioxidant Enzyme Activities
4.6. DNA Extraction and ISSR-PCR Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kordrostami, M.; Rabiei, B.; Kumleh, H. Association analysis, genetic diversity and haplotyping of rice plants under salt stress using SSR markers linked to SalTol and morpho-physiological characteristics. Plant Syst. Evol. 2016, 302, 871–890. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.; Srivastava, A.K.; Abdel El-sadek, M.S.; Kordrostami, M.; Tran, L.S.P. Titanium dioxide nanoparticles improve growth and enhance tolerance of broad bean plants under saline soil conditions. Land Degrad. Dev. 2018, 29, 1065–1073. [Google Scholar] [CrossRef]
- Oyiga, B.C.; Sharma, R.C.; Baum, M.; Ogbonnaya, F.C.; Léon, J.; Ballvora, A. Allelic variations and differential expressions detected at quantitative trait loci for salt stress tolerance in wheat. Plant Cell Environ. 2018, 41, 919–935. [Google Scholar] [CrossRef] [PubMed]
- Ning, T. Effects of seawater stress on seed germination and growth of pepper. J. Nanjing Xiaozhuang Univ. 2017, 6, 019. [Google Scholar]
- Negrão, S.; Schmöckel, S.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.; Mostofa, M.G.; Rahman, M.M.; Abdel-Farid, I.B.; Tran, L.S.P. Extracts from yeast and carrot roots enhance maize performance under seawater-induced salt stress by altering physio-biochemical characteristics of stressed plants. J. Plant Growth Regul. 2019. [Google Scholar] [CrossRef]
- Schmitt, F.J.; Allakhverdiev, S.I. Reactive Oxygen Species: Signaling between Hierarchical Levels in Plants; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Schieber, M.; Chandel, N. Ros function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Kundu, P.; Gill, R.; Ahlawat, S.; Anjum, N.A.; Sharma, K.K.; Ansari, A.A.; Hasanuzzaman, M.; Ramakrishna, A.; Chauhan, N.; Tuteja, N. Targeting the redox regulatory mechanisms for abiotic stress tolerance in crops. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Elsevier: Amsterdam, The Netherlands, 2018; pp. 151–220. [Google Scholar]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945. [Google Scholar] [CrossRef] [Green Version]
- Hung, S.; Yu, C.; Lin, C.H. Hydrogen peroxide functions as a stress signal in plants. Bot. Bull. Acad. Sin. 2005, 46, 1–10. [Google Scholar]
- Ellouzi, H.; Sghayar, S.; Abdelly, C. H2O2 seed priming improves tolerance to salinity; drought and their combined effect more than mannitol in Cakile maritima when compared to Eutrema salsugineum. J. Plant Physiol. 2017, 210, 38–50. [Google Scholar] [CrossRef]
- Zandalinas, S.; Mittler, R. Ros-induced ROS release in plant and animal cells. Free Radic. Biol. Med. 2018, 122, 21–27. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Akbar, A.; Askari, S.H.; Iqbal, M.; Rasheed, R.; Hussain, I. Recent advances in abiotic stress tolerance of plants through chemical priming: An overview. In Advances in Seed Priming; Springer: Berlin/Heidelberg, Germany, 2018; pp. 51–79. [Google Scholar]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Garrido-Cardenas, J.A.; Mesa-Valle, C.; Manzano-Agugliaro, F. Trends in plant research using molecular markers. Planta 2017, 1–15. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ng, W.L.; Lamasudin, D.U.; Mohamed, R. Inter-simple sequence repeat markers reveal genetic relatedness between natural Aquilaria populations in peninsular Malaysia. Chiang Mai J. Sci. 2018, 45, 1307–1317. [Google Scholar]
- Ng, W.; Tan, S. Inter-simple sequence repeat (ISSR) markers: Are we doing it right. ASM Sci. J. 2015, 9, 30–39. [Google Scholar]
- Vijayan, K. Inter simple sequence repeat (ISSR) polymorphism and its application in mulberry genome analysis. Int. J. Ind. Entomol. 2005, 10, 79–86. [Google Scholar]
- Khatab, I.A.; Samah, M.A. Development of agronomical and molecular genetic markers associated with salt stress tolerance in some barley genotypes. Curr. Res. J. Biol. Sci. 2013, 5, 198–204. [Google Scholar] [CrossRef]
- Khalil, R.M. Molecular and biochemical markers associated with salt tolerance in some sorghum genotypes. World Appl. Sci. J. 2013, 22, 459–469. [Google Scholar] [CrossRef]
- Markad, N.R.; Kale, A.A.; Pawar, B.D.; Jadhav, A.S.; Patil, S.C. Molecular characterization of sugarcane (Saccharum officinarum l.) genotypes in relation to salt tolerance. Bioscan 2014, 9, 1785–1788. [Google Scholar]
- Yaycili, O.; Sen, A.; Alikamanoglu, S. Induced of salt tolerance wheat (Triticum aestivum L.) mutants with gamma radiation and determining molecular analysis by ISSR. Procedia Environ. Sci. 2015, 196. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Azzarello, E.; Huang, Y.; Pandolfi, C.; Su, N.; Wu, Q.; Cai, S.; Bazihizina, N.; Wang, L. Na+ extrusion from the cytosol and tissue-specific Na+ sequestration in roots confer differential salt stress tolerance between durum and bread wheat. J. Exp. Bot. 2018, 18, 3987–4001. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Hussain, M.; Ahmad, S.; Hussain, S.; Lal, R.; Ul-Allah, S.; Nawaz, A. Rice in saline soils: Physiology, biochemistry, genetics, and management. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2018; Volume 148, pp. 231–287. [Google Scholar]
- Abass, S.M.; Mohamed, H.I. Alleviation of adverse effects of drought stress on common bean (Phaseolus vulgaris L.) by exogenous application of hydrogen peroxide. Bangladesh J. Bot. 2011, 40, 75–83. [Google Scholar] [CrossRef]
- Kasim, W.A.; Nessem, A.A.; Gaber, A. Alleviation of drought stress in Vicia Faba by seed priming with ascorbic acid or extracts of garlic and carrot. Egypt. J. Bot. 2017, 57, 45–49. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Michailidi, E.; Tzortzakis, N. Physiological and biochemical responses of Lavandula angustifolia to salinity under mineral foliar application. Front. Plant Sci. 2018, 9, 489. [Google Scholar] [CrossRef]
- Çavusoglu, K.; Kabar, K. Effects of hydrogen peroxide on the germination and early seedling growth of barley under nacl and high temperature stresses. Eurasian J. Biosci. 2010, 4. [Google Scholar] [CrossRef]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Medeiros, J.V.R.; Gomes-Filho, E. Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize plants. J. Plant Physiol. 2005, 162, 1114–1122. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Yamamoto, K.; Tawaratsumida, T.; Yuasa, T.; Iwaya-Inoue, M. Hydrogen peroxide scavenging regulates germination ability during wheat (Triticum aestivum L.) seed maturation. Plant Signal. Behav. 2008, 3, 183–188. [Google Scholar] [CrossRef]
- Sayed, S.; Gadallah, M. Hydrogen peroxide supplementation alleviates the deleterious effects of cadmium on photosynthetic pigments and oxidative stress and improves growth, yield and pods quality of pea (Pisum sativum l.) plants. Acta Physiol. Plant. 2019, 41, 113. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rasheed, R.; Hussain, I.; Iqbal, M.; Haider, M.Z.; Parveen, S.; Sajid, M.A. Hydrogen peroxide modulates antioxidant system and nutrient relation in maize (Zea mays L.) under water-deficit conditions. Arch. Agron. Soil Sci. 2015, 61, 507–523. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Becker, V.I.; Goessling, J.W.; Duarte, B.; Caçador, I.; Liu, F.; Rosenqvist, E.; Jacobsen, S.E. Combined effects of soil salinity and high temperature on photosynthesis and growth of quinoa plants (Chenopodium quinoa). Funct. Plant Biol. 2017, 44, 665–678. [Google Scholar] [CrossRef]
- Gomes, M.A.D.C.; Pestana, I.A.; Santa-Catarina, C.; Hauser-Davis, R.A.; Suzuki, M.S. Salinity effects on photosynthetic pigments, proline, biomass and nitric oxide in Salvinia auriculata aubl. Acta Limnol. Bras. 2017, 29. [Google Scholar] [CrossRef]
- Minguez-Mosquera, M.; Jaren-Galan, M.; Garrido-Fernandez, J. Lipoxygenase activity during pepper ripening and processing of paprika. Phytochemistry 1993, 32, 1103–1108. [Google Scholar] [CrossRef]
- Ziaf, K.; Amjad, M.; Pervez, M.A.; Iqbal, Q.; Rajwana, I.A.; Ayyub, M. Evaluation of different growth and physiological traits as indices of salt tolerance in hot pepper (Capsicum annuum l.). Pak. J. Bot. 2009, 41, 1797–1809. [Google Scholar]
- Forlani, G.; Bertazzini, M.; Cagnano, G. Stress-driven increase in proline levels, and not proline levels themselves, correlates with the ability to withstand excess salt in a group of 17 italian rice genotypes. Plant Biol. 2018, 336–342. [Google Scholar] [CrossRef]
- He, L.; Gao, Z. Pretreatment of seed with h2o2 enhances drought tolerance of wheat (Triticum aestivum L.) seedlings. Afr. J. Biotechnol. 2009, 8. [Google Scholar] [CrossRef]
- Abdel Latef, A.A. Ameliorative effect of calcium chloride on growth, antioxidant enzymes, protein patterns and some metabolic activities of canola (Brassica napus L.) under seawater stress. J. Plant Nutr. 2011. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Alhmad, M.F.; Hammad, S.A. Foliar application of fresh moringa leaf extract overcomes salt stress in fenugreek (Trigonella foenum-graecum) plants. Egypt. J. Bot. 2011, 157–179. [Google Scholar]
- Wahid, A.; Perveen, M.; Gelani, S.; Basra, S.M. Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J. Plant Physiol. 2007, 164, 283–294. [Google Scholar] [CrossRef]
- Gondim, F.A.; Gomes-Filho, E.; Lacerda, C.F.; Prisco, J.T.; Azevedo Neto, A.D.; Marques, E.C. Pretreatment with h2o2 in maize seeds: Effects on germination and seedling acclimation to salt stress. Braz. J. Plant Physiol. 2010, 22, 103–112. [Google Scholar] [CrossRef]
- Santhy, V.; Meshram, M.; Wakde, R.; Kumari, P.V. Hydrogen peroxide pre-treatment for seed enhancement in cotton (gossypim hirsutum L.). Afr. J. Agric. Res. 2014, 9, 1982–1989. [Google Scholar] [CrossRef]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in nacl-stressed rice (Oryza sativa L.)—differential response in salt-tolerant and sensitive varieties. Plant Sci. 2003, 165, 1411–1418. [Google Scholar] [CrossRef]
- Kumar, V.; Shriram, V.; Nikam, T.; Jawali, N.; Shitole, M.G. Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes differing in salt tolerance. Arch. Agron. Soil Sci. 2009, 55, 379–394. [Google Scholar] [CrossRef]
- Rubio, M.C.; González, E.M.; Minchin, F.R.; Webb, K.J.; Arrese-Igor, C.; Ramos, J.; Becana, M. Effects of water stress on antioxidant enzymes of leaves and nodules of transgenic alfalfa overexpressing superoxide dismutases. Physiol. Plant. 2002, 115, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- Yu, S.; Wang, W.; Wang, B. Recent progress of salinity tolerance research in plants. Russ. J. Genet. 2012, 48, 497–505. [Google Scholar] [CrossRef]
- Saha, S.; Sadle, J.; Van Der Heiden, C.; Sternberg, L. Salinity, groundwater, and water uptake depth of plants in coastal uplands of everglades national park (florida, USA). Ecohydrology 2015, 8, 128–136. [Google Scholar] [CrossRef]
- Bray, C.M.; West, C.E. DNA repair mechanisms in plants: Crucial sensors and effectors for the maintenance of genome integrity. New Phytol. 2005, 168, 511–528. [Google Scholar] [CrossRef]
- Saha, J.; Brauer, E.K.; Sengupta, A.; Popescu, S.C.; Gupta, K.; Gupta, B. Polyamines as redox homeostasis regulators during salt stress in plants. Front. Environ. Sci. 2015, 3, 21. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Tazeb, A.; Haileselassie, T.; Tesfaye, K. Molecular characterization of introduced sugarcane genotypes in ethiopia using inter simple sequence repeat (ISSR) molecular markers. Afr. J. Biotechnol. 2017, 16, 434–449. [Google Scholar] [CrossRef]
- Younis, R.A.; Ahmed, M.; El-Menshawy, M.M. Molecular genetic markers associated with salt tolerance in grain sorghum. Arab J. Biotechnol. 2007, 10, 249–258. [Google Scholar]
- Abdi, A.A.; Sofalian, O.; Asghari, A.; Shokrpour, M.; Baqheri, H.; Masoumi, S.Y. Inter-simple sequence repeat (ISSR) markers to study genetic diversity among cotton cultivars in associated with salt tolerance. Not. Sci. Biol. 2012, 4, 120–126. [Google Scholar] [CrossRef]
- Lakshmanan, V.; Reddampalli Venkataramareddy, S.; Neelwarne, B. Molecular analysis of genetic stability in long-term micropropagated shoots of banana using rapd and issr markers. Electron. J. Biotechnol. 2007, 10, 106–113. [Google Scholar] [CrossRef]
- Shalaby, T.A.; El-Banna, A. Molecular and horticultural characteristics of in vitro induced tomato mutants. J. Agric. Sci. 2013, 5, 155. [Google Scholar] [CrossRef]
- Farokhzadeh, S.; Fakheri, B. Marker-assisted selection for disease resistance: Applications in breeding. Int. J. Agric. Crop Sci. 2014, 7, 1392–1405. [Google Scholar] [CrossRef]
- Krupa-Małkiewicz, M.; Bienias, A. BSA and molecular markers screening for salt stress tolerant mutant of petunia obtained in in vitro culture. Ciênc. Rural 2018, 48. [Google Scholar] [CrossRef]
- Mahgoub, H.A.; Sofy, A.R.; Abdel-Azeem, E.A.; Abo-Zahra, M.S. Molecular markers associated with salt-tolerance of different soybean (Glycine max L.) cultivars under salt stress. Int. J. Adv. Res. Biol. Sci 2016, 3, 241–267. [Google Scholar]
- Reddy, C.S.; Babu, A.P.; Swamy, B.M.; Kaladhar, K.; Sarla, N. ISSR markers based on ga and ag repeats reveal genetic relationship among rice varieties tolerant to drought, flood, or salinity. J. Zhejiang Univ. Sci. B 2009, 10, 133–141. [Google Scholar] [CrossRef]
- Kordrostami, M.; Rahimi, M. Molecular markers in plants: Concepts and applications. Genet. 3rd Millenn. 2015, 13, 4024–4031. [Google Scholar]
- Bhutta, W.M.; Hanif, M. Identification of rapd markers linked to salinity tolerance in wheat. Afr. J. Biotechnol. 2013, 12. [Google Scholar] [CrossRef]
- Masojć, P.; Lebiecka, K.; Milczarski, P.; Wiśniewska, M.; Łań, A.; Owsianicki, R. Three classes of loci controlling preharvest sprouting in rye (Secale cereale L.) discerned by means of bidirectional selective genotyping (bsg). Euphytica 2009, 170, 123. [Google Scholar] [CrossRef]
- Sharma, K.; Agrawal, V.; Gupta, S.; Kumar, R.; Prasad, M. ISSR marker-assisted selection of male and female plants in a promising dioecious crop: Jojoba (Simmondsia chinensis). Plant Biotechnol. Rep. 2008, 2, 239–243. [Google Scholar] [CrossRef]
- El-Nahas, A.; El-Shazly, H.; Ahmed, S.M.; Omran, A. Molecular and biochemical markers in some lentil (Lens culinaris medik.) genotypes. Ann. Agric. Sci. 2011, 56, 105–112. [Google Scholar] [CrossRef]
- Maha, A.; Sanaa, I.; Mabrook, Y.; Amira, Y.; Gouda, M. Evaluation of some egyptian bread wheat (Triticum aestivum) cultivars under salinity stress. Alex. Sci. Exch. J. 2017, 38, 260. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents; Portland Press Limited: London, UK, 1983. [Google Scholar]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Abdel Latef, A.; Tran, L.S.P. Impacts of priming with silicon on the growth and tolerance of maize plants to alkaline stress. Front. Plant Sci. 2016, 7, 243. [Google Scholar] [CrossRef]
- Williams, V.; Twine, S. Flame Photometric Method for Sodium, Potassium and Calcium; Springer International Publishing: Cham, Switzerland, 1960; Volume 5, pp. 3–5. [Google Scholar]
- Ahmad, P.; Abdel Latef, A.; Abd Allah, E.; Hashem, A.; Sarwat, M.; Anjum, N.; Gucel, S. Calcium and potassium supplementation enhanced growth, osmolyte secondary metabolite production, and enzymatic antioxidant machinery in cadmium-exposed chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 513. [Google Scholar] [CrossRef]
- Chen, G.X.; Asada, K. Inactivation of ascorbate peroxidase by thiols requires hydrogen peroxide. Plant Cell Physiol. 1992, 33, 117–123. [Google Scholar] [CrossRef]
- Maehly, A.; Chance, B. Methods of Biochemical Analysis; Wiley-Interscience: Hoboken, NJ, USA, 1954. [Google Scholar]
- Aebi, H. [13] Catalase in vitro. In Methods Enzymol; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Adhikari, S.; Saha, S.; Bandyopadhyay, T.K.; Ghosh, P. Efficiency of issr marker for characterization of Cymbopogon germplasms and their suitability in molecular barcoding. Plant Syst. Evol. 2015, 301, 439–450. [Google Scholar] [CrossRef]
- Wicklin, R. SAS/IML Software: Usage and Reference; SAS Institute inc.: Cary, NC, USA, 2010. [Google Scholar]
- Green, S.; Salkind, N. Using SPSS for Windows and Macintosh, Books a La Carte; Pearson: London, UK, 2016. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- StatGraphics, C. Statgraphics centurion xvii. In User Manual; Statgraphics Technologies, Inc.: The Plains, VA, USA, 2014; Volume 17. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Proline | Na+ | K+ | Ca2+ | Mg2+ |
|---|---|---|---|---|---|
| W + W (0%) (Gemmiza 11) | 0.50 d ± 0.05 | 2.92 d ± 0.44 | 8.50 d ± 1.51 | 0.93 d,e ± 0.05 | 0.23 e,f ± 0.03 |
| H2O2 + W (0%) (Gemmiza 11) | 0.17 f ± 0.02 | 2.23 d ± 0.11 | 17.73 a ± 2.26 | 1.16 c,d ± 0.06 | 0.36 d ± 0.02 |
| W + SW (35%) (Gemmiza 11) | 1.80 a ± 0.03 | 36.64 a ± 3.30 | 7.40 d ± 1.05 | 0.53 f ± 0.02 | 0.17 f ± 0.02 |
| H2O2 + SW (35%) (Gemmiza 11) | 1.25 c ± 0.05 | 24.78 b ± 4.60 | 14.00 b,c ± 4.29 | 0.78 e,f ± 0.05 | 0.30 d,e ± 0.05 |
| W + W (0%) (Misr 1) | 0.17 f ± 0.04 | 2.63 d ± 0.08 | 12.37 c ± 0.87 | 1.06 d ± 0.03 | 0.71 c ± 0.06 |
| H2O2 + W (0%) (Misr 1) | 0.38 e ± 0.07 | 2.31 d ± 0.09 | 17.00 a,b ± 1.51 | 1.41 c ± 0.03 | 1.24 b ± 0.05 |
| W + SW (35%) (Misr 1) | 1.59 b ± 0.07 | 20.89 c ± 2.05 | 13.63 b,c ± 0.25 | 2.89 b ± 0.40 | 1.28 b ± 0.03 |
| H2O2 + SW (35%) (Misr 1) | 1.76 a ± 0.03 | 17.48 c ± 0.89 | 15.20 a–c ± 0.96 | 4.53 a ± 0.16 | 1.45 a ± 0.10 |
| Primer Name | Sequence | Motif | OB | PB | P% |
|---|---|---|---|---|---|
| ISSR-1 | 5’-AGAGAGAGAGAGAGAGC-3’ | (AG)8C | 18 | 13 | 72.22 |
| ISSR-2 | 5’-ACACACACACACACACT-3’ | (AC)8T | 11 | 6 | 54.54 |
| ISSR-3 | 5’-ACACACACACACACACG-3’ | (AC)8G | 25 | 23 | 92 |
| ISSR-4 | 5’-CGCGATAGATAGATAGAT-3’ | CGC(GATA)4 | 17 | 14 | 82.35 |
| ISSR-5 | 5’-GACGATAGATAGATAGATA-3’ | GAC(GATA)4 | 19 | 16 | 84.21 |
| ISSR-6 | 5’-GACAGACAGACAGACAAT-3’ | (GACA)4AT | 11 | 9 | 81.81 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel Latef, A.A.H.; Kordrostami, M.; Zakir, A.; Zaki, H.; Saleh, O.M. Eustress with H2O2 Facilitates Plant Growth by Improving Tolerance to Salt Stress in Two Wheat Cultivars. Plants 2019, 8, 303. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8090303
Abdel Latef AAH, Kordrostami M, Zakir A, Zaki H, Saleh OM. Eustress with H2O2 Facilitates Plant Growth by Improving Tolerance to Salt Stress in Two Wheat Cultivars. Plants. 2019; 8(9):303. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8090303
Chicago/Turabian StyleAbdel Latef, Arafat Abdel Hamed, Mojtaba Kordrostami, Ali Zakir, Hoida Zaki, and Osama Moseilhy Saleh. 2019. "Eustress with H2O2 Facilitates Plant Growth by Improving Tolerance to Salt Stress in Two Wheat Cultivars" Plants 8, no. 9: 303. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8090303