
Estimating the Time Reproduction Number in Kupang City Indonesia, 2016–2020, and Assessing the Effects of Vaccination and Different Wolbachia Strains on Dengue Transmission Dynamics

 ,
,
Abstract
:1. Introduction
2. Formulation of Mathematical Model
2.1. Modeling Framework for Mosquito Population
2.2. Modeling Framework for Host-Vector Involving Wolbachia and Vaccination
3. Basic Reproduction Number
4. Parameter Estimation
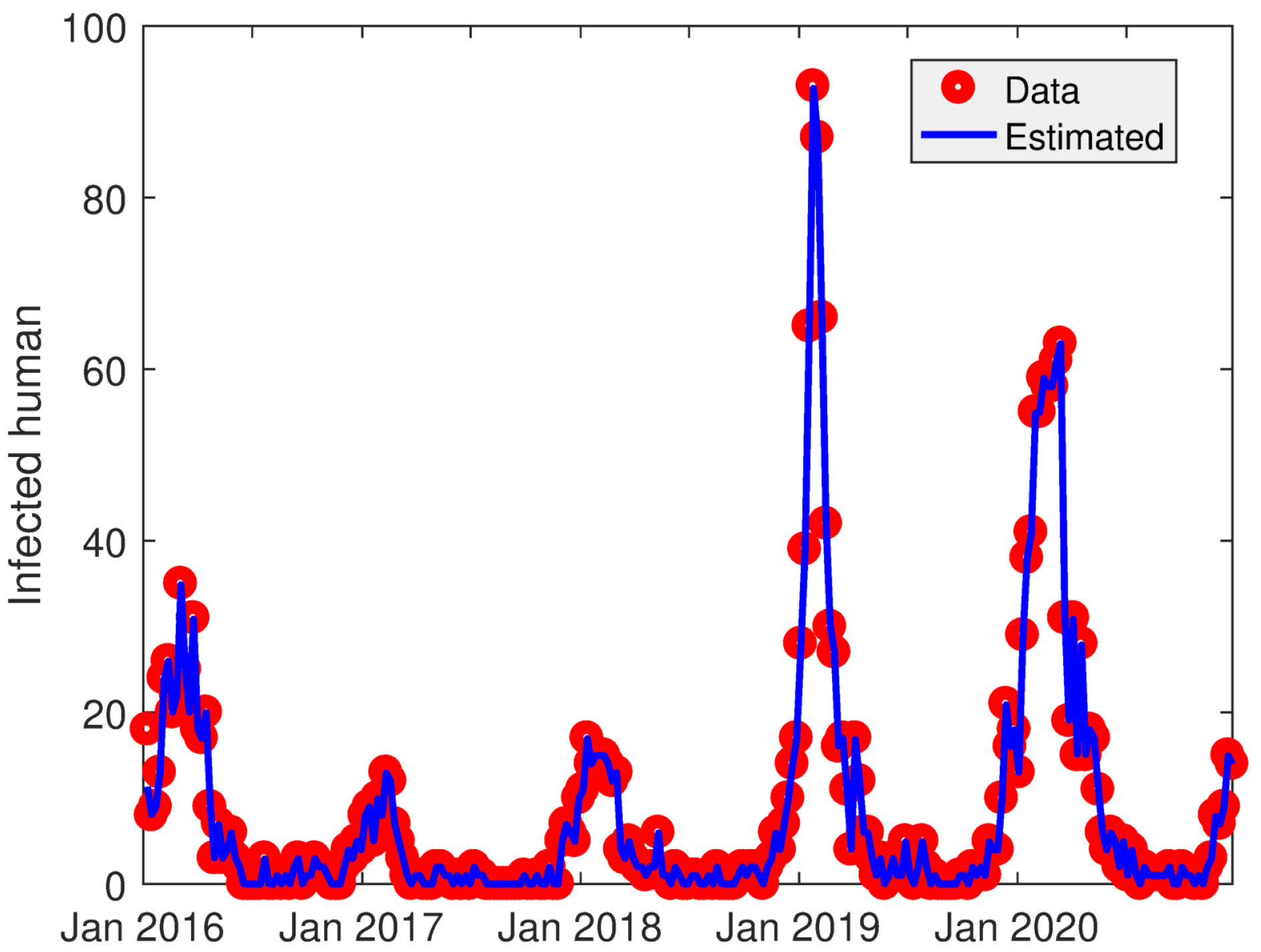
4.1. Data
4.2. Mathematical Model for Parameter Estimation
4.3. Discrete Time Stochastic Augmented Model
4.4. Extended Kalman Filter
4.5. Estimation of Reproduction Number
5. Sensitivity Analysis
6. Numerical Simulations in the Absence and Presence of Wolbachia
6.1. Numerical Simulations in the Absence of Vaccination
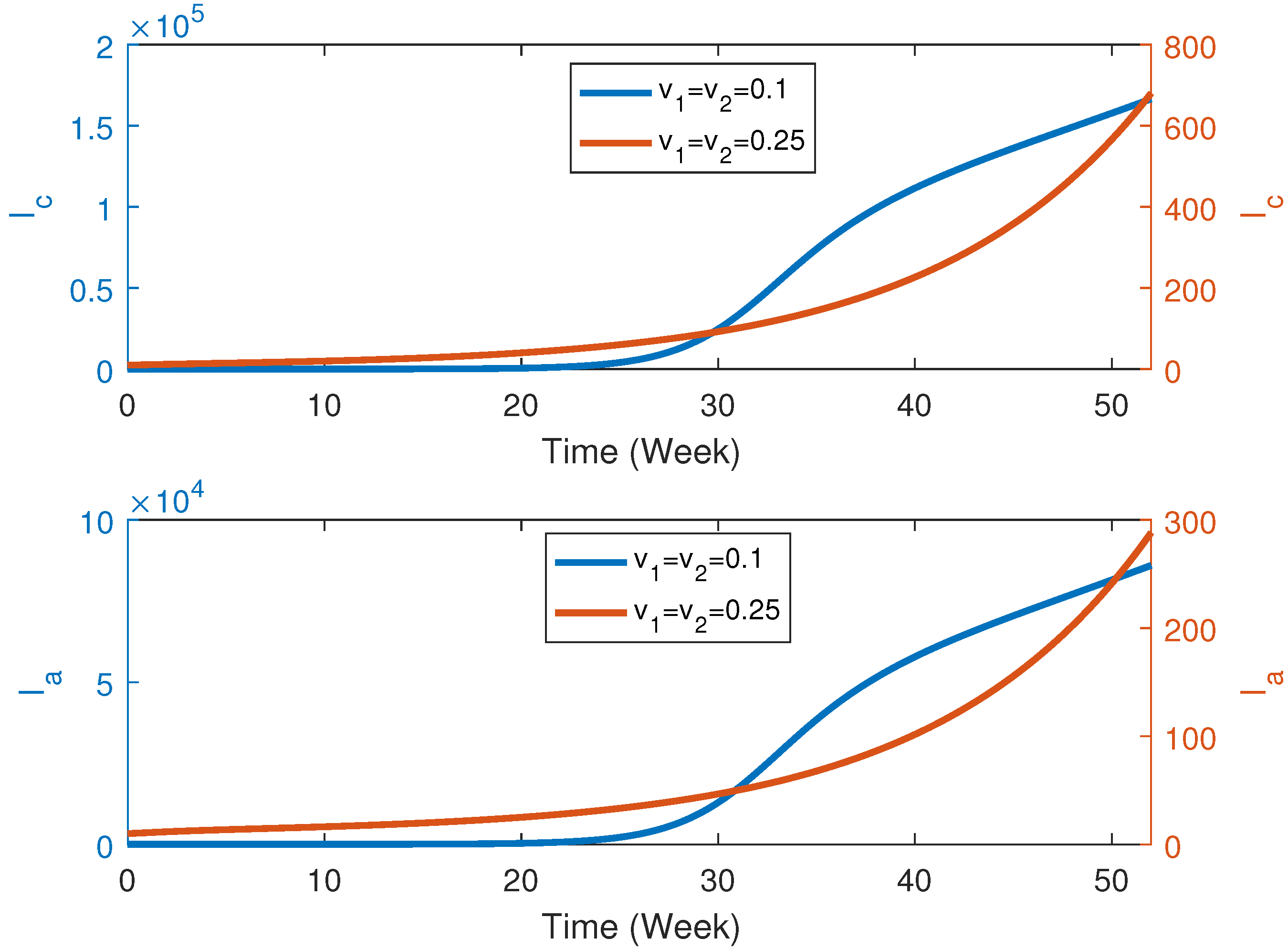
6.2. Numerical Simulation in the Presence of Vaccination
6.3. Numerical Solution for Dengue Transmission Dynamics with Vaccination with Wolbachia
7. Optimal Control Approach
7.1. The Existence and Characterization of Optimal Control
- A1.
- The solution of the model with non-negative initial conditions and the associated control function in is non-empty.
- A2.
- The control set is convex and closed.
- A3.
- The dengue model can be expressed as a linear function of , with time and state dependent coefficients.
- A4.
- There exists constant , and such that the integrand in (12) is convex and satisfy
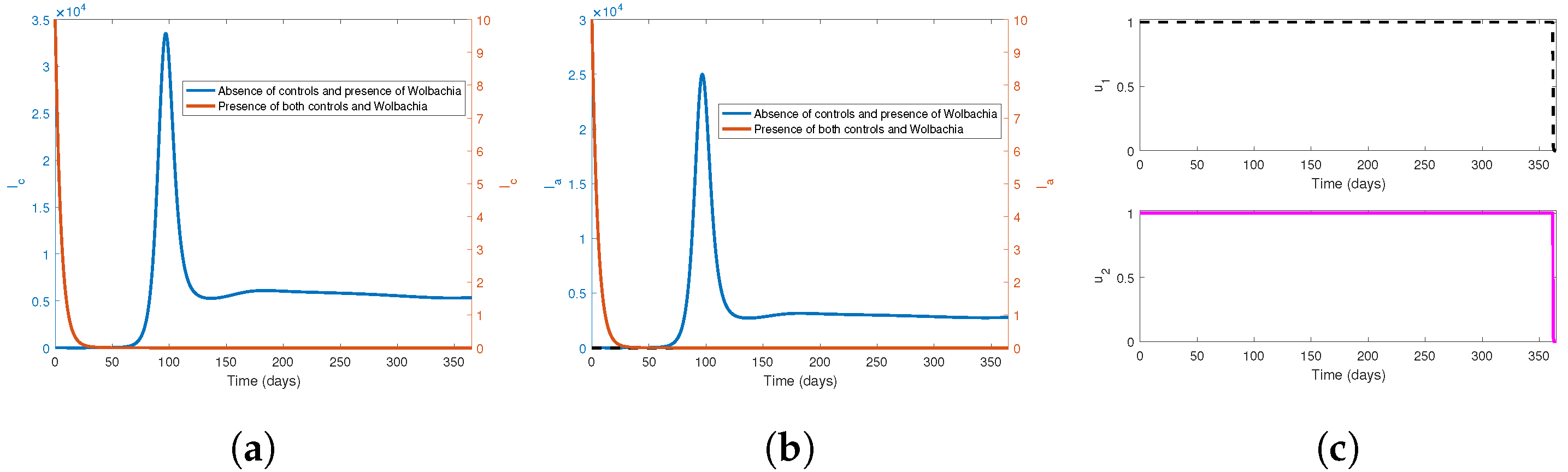
7.2. Numerical Simulations of Optimal Control
8. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cattarino, L.; Rodriguez-Barraquer, I.; Imai, N.; Cummings, D.A.T.; Ferguson, N.M. Mapping global variation in dengue transmission intensity. Sci. Transl. Med. 2020, 12, eaax4144. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The Impact of the Newly Licensed Dengue Vaccine in Endemic Countries. PLoS Negl. Trop. Dis. 2016, 10, e0005179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The risks behind Dengvaxia recommendation. Lancet Infect. Dis. 2016, 16, 882–883. [Google Scholar] [CrossRef]
- Ndii, M.Z.; Mage, A.R.; Messakh, J.J.; Djahi, B.S. Optimal vaccination strategy for dengue transmission in Kupang city, Indonesia. Heliyon 2020, 6, e05345. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Salcuni, P.; Pompa, M.G.; Declich, S.; Rizzo, C. Estimated Imported Infections of Chikungunya and Dengue in Italy, 2008 to 2011. J. Travel Med. 2012, 19, 294–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorigatti, I.; McCormack, C.; Nedjati-Gilani, G.; Ferguson, N.M. Using Wolbachia for Dengue Control: Insights from Modelling. Trends Parasitol. 2018, 34, 102–113. [Google Scholar] [CrossRef]
- Biswal, S.; Reynales, H.; Saez-Llorens, X.; Lopez, P.; Borja-Tabora, C.; Kosalaraksa, P.; Sirivichayakul, C.; Watanaveeradej, V.; Rivera, L.; Espinoza, F.; et al. Efficacy of a Tetravalent Dengue Vaccine in Healthy Children and Adolescents. N. Engl. J. Med. 2019, 381, 2009–2019. [Google Scholar] [CrossRef]
- Biswal, S.; Borja-Tabora, C.; Martinez Vargas, L.; Velásquez, H.; Theresa Alera, M.; Sierra, V.; Johana Rodriguez-Arenales, E.; Yu, D.; Wickramasinghe, V.P.; Duarte Moreira, E.; et al. Efficacy of a tetravalent dengue vaccine in healthy children aged 4–16 years: A randomised, placebo-controlled, phase 3 trial. Lancet 2020, 395, 1423–1433. [Google Scholar] [CrossRef]
- Villar, L.; Dayan, G.H.; Arredondo-García, J.L.; Rivera, D.M.; Cunha, R.; Deseda, C.; Reynales, H.; Costa, M.S.; Morales-Ramírez, J.O.; Carrasquilla, G.; et al. Efficacy of a Tetravalent Dengue Vaccine in Children in Latin America. N. Engl. J. Med. 2015, 372, 113–123. [Google Scholar] [CrossRef]
- Pagendam, D.E.; Trewin, B.J.; Snoad, N.; Ritchie, S.A.; Hoffmann, A.A.; Staunton, K.M.; Paton, C.; Beebe, N. Modelling the Wolbachia incompatible insect technique: Strategies for effective mosquito population elimination. BMC Biol. 2020, 18, 161. [Google Scholar] [CrossRef]
- Ndii, M.Z.; Hickson, R.; Allingham, D.; Mercer, G. Modelling the transmission dynamics of dengue in the presence of Wolbachia. Math. Biosci. 2015, 262, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, N.M.; Hue Kien, D.T.; Clapham, H.; Aguas, R.; Trung, V.T.; Bich Chau, T.N.; Popovici, J.; Ryan, P.A.; O’Neill, S.L.; McGraw, E.A.; et al. Modeling the impact on virus transmission of Wolbachia-mediated blocking of dengue virus infection of Aedes aegypti. Sci. Transl. Med. 2015, 7, 279ra37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beebe, N.W.; Pagendam, D.; Trewin, B.J.; Boomer, A.; Bradford, M.; Ford, A.; Liddington, C.; Bondarenco, A.; Barro, P.J.D.; Gilchrist, J.; et al. Releasing incompatible males drives strong suppression across populations of wild and Wolbachia-carrying Aedes aegypti in Australia. Proc. Natl. Acad. Sci. USA 2021, 118, e2106828118. [Google Scholar] [CrossRef] [PubMed]
- Turley, A.P.; Moreira, L.A.; O’Neill, S.L.; McGraw, E.A. Wolbachia Infection Reduces Blood–Feeding Success in the Dengue Fever Mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 2009, 3, e516. [Google Scholar] [CrossRef]
- Ndii, M.Z. Modelling the Use of Vaccine and Wolbachia on Dengue Transmission Dynamics. Trop. Med. Infect. Dis. 2020, 5, 78. [Google Scholar] [CrossRef]
- Cardona-Salgado, D.; Campo-Duarte, D.E.; Sepulveda-Salcedo, L.S.; Vasilieva, O. Wolbachia-based biocontrol for dengue reduction using dynamic optimization approach. Appl. Math. Modell. 2020, 82, 125–149. [Google Scholar] [CrossRef]
- Zhang, H.; Lui, R. Releasing Wolbachia-infected Aedes aegypti to prevent the spread of dengue virus: A mathematical study. Infect. Dis. Modell. 2020, 5, 142–160. [Google Scholar] [CrossRef]
- Ogunlade, S.T.; Adekunle, A.I.; Meehan, M.T.; Rojas, D.P.; McBryde, E.S. Modeling the potential of wAu-Wolbachia strain invasion in mosquitoes to control Aedes-borne arboviral infections. Sci. Rep. 2020, 10, 16812. [Google Scholar] [CrossRef]
- Aguiar, M.; Anam, V.; Blyuss, K.B.; Estadilla, C.D.S.; Guerrero, B.V.; Knopoff, D.; Kooi, B.W.; Srivastav, A.K.; Steindorf, V.; Stollenwerk, N. Mathematical models for dengue fever epidemiology: A 10-year systematic review. Phys. Life Rev. 2022, 40, 65–92. [Google Scholar] [CrossRef]
- Shim, E. Optimal dengue vaccination strategies of seropositive individuals. Math. Biosci. Eng. 2019, 16, 1171–1189. [Google Scholar] [CrossRef]
- Junsawang, P.; Zuhra, S.; Sabir, Z.; Raja, M.A.Z.; Shoaib, M.; Botmart, T.; Weera, W. Numerical Simulations of Vaccination and Wolbachia on Dengue Transmission Dynamics in the Nonlinear Model. IEEE Access 2022, 10, 31116–31144. [Google Scholar] [CrossRef]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathogens 2018, 14, e1006815. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.D.; McElroy, K.; Day, A.; Higgs, S.; O’Neill, S.L. Impact of Wolbachia on Infection with Chikungunya and Yellow Fever Viruses in the Mosquito Vector Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, 1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The WMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Hussain, M.; Lu, G.; Torres, S.; Edmonds, J.H.; Kay, B.H.; Khromykh, A.A.; Asgari, S. Effect of Wolbachia on Replication of West Nile Virus in a Mosquito Cell Line and Adult Mosquitoes. J. Virol. 2013, 87, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Ndii, M.Z.; Hickson, R.I.; Mercer, G.N. Modelling the introduction of Wolbachia into Aedes aegypti to reduce dengue transmission. ANZIAM J. 2012, 53, 213–227. [Google Scholar]
- Diekmann, O.; Heesterbeek, J.A.P.; Roberts, M.G. The construction of next-generation matrices for compartmental epidemic models. J. R. Soc. Interface 2010, 7, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Hasan, A.; Nasution, Y. A compartmental epidemic model incorporating probable cases to model COVID-19 outbreak in regions with limited testing capacity. ISA Trans. 2021, 124, 157–163. [Google Scholar] [CrossRef]
- Yi, C.; Cohnstaedt, L.W.; Scoglio, C.M. SEIR-SEI-EnKF: A New Model for Estimating and Forecasting Dengue Outbreak Dynamics. IEEE Access 2021, 9, 156758–156767. [Google Scholar] [CrossRef]
- Hasan, A.; Susanto, H.; Tjahjono, V.; Kusdiantara, R.; Putri, E.; Nuraini, N.; Hadisoemarto, P. A new estimation method for COVID-19 time-varying reproduction number using active cases. Sci. Rep. 2022, 12, 6675. [Google Scholar] [CrossRef]
- Ndii, M.Z.; Anggriani, N.; Messakh, J.J.; Djahi, B.S. Estimating the reproduction number and designing the integrated strategies against dengue. Results Phys. 2021, 27, 104473. [Google Scholar] [CrossRef]
- Herdicho, F.F.; Chukwu, W.; Tasman, H. An optimal control of malaria transmission model with mosquito seasonal factor. Results Phys. 2021, 25, 104238. [Google Scholar] [CrossRef]
- Abidemi, A.; Aziz, N.A.B. Optimal control strategies for dengue fever spread in Johor, Malaysia. Comput. Methods Programs Biomed. 2020, 196, 105585. [Google Scholar] [CrossRef] [PubMed]
- Olaniyi, S.; Mukamuri, M.; Okosun, K.; Adepoju, O. Mathematical analysis of a social hierarchy-structured model for malaria transmission dynamics. Results Phys. 2022, 34, 104991. [Google Scholar] [CrossRef]
- Fleming, W.; Rishel, R. Deterministic and Stochastic Optimal Control; Stochastic Modelling and Applied Probability; Springer: New York, NY, USA, 2012. [Google Scholar]
- Lukes, D. Differential Equations: Classical to Controlled; Number v. 162 in Differential Equations: Classical to Controlled; Academic Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Abidemi, A.; Olaniyi, S.; Adepoju, O.A. An Explicit Note on the Existence Theorem of Optimal Control Problem. J. Phys. Conf. Ser. 2022, 2199, 012021. [Google Scholar] [CrossRef]
- Sepulveda, L.S.; Vasilieva, O. Optimal control approach to dengue reduction and prevention in Cali, Colombia. Math. Methods Appl. Sci. 2016, 39, 5475–5496. [Google Scholar] [CrossRef]
- Khan, A.; Hassan, M.; Imran, M. Estimating the basic reproduction number for single-strain dengue fever epidemics. Infect. Dis. Poverty 2014, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Fatmawati. Dengue infection modeling and its optimal control analysis in East Java, Indonesia. Heliyon 2021, 7, e06023. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Unit |
|---|---|---|---|
| Recruitment rate of human | week | ||
| Natural Death Rate | week | ||
| Progression rate from child to adult | week | ||
| Transmission rate from Non-W to human | week | ||
| Transmission rate from W to human | 0.2098 | week | |
| Progression rate from recovered child to susceptible child | week | ||
| Progression rate from recovered adult to susceptible adult | week | ||
| Recovery rate | 7/5 | week | |
| Reproduction rate of non-W mosquitoes | 8.75 | week | |
| Maturation rate of non-W mosquitoes | 2 | week | |
| Death rate of aquatic non-W mosquitoes | 1/2 | week | |
| Maternal transmission | 0.9 | n/a | |
| Transmission rate from human to Non-W mosquitoes | week | ||
| Reproductive rate of W-mosquitoes | week | ||
| Maturation rate of W-mosquitoes | 2 | week | |
| Loss of Wolbachia infections | week | ||
| Death rate of adult non-W mosquitoes | 1/2 | week | |
| Death rate of adult W mosquitoes | 0.45 | week |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndii, M.Z.; Beay, L.K.; Anggriani, N.; Nukul, K.N.; Djahi, B.S. Estimating the Time Reproduction Number in Kupang City Indonesia, 2016–2020, and Assessing the Effects of Vaccination and Different Wolbachia Strains on Dengue Transmission Dynamics. Mathematics 2022, 10, 2075. https://0-doi-org.brum.beds.ac.uk/10.3390/math10122075
Ndii MZ, Beay LK, Anggriani N, Nukul KN, Djahi BS. Estimating the Time Reproduction Number in Kupang City Indonesia, 2016–2020, and Assessing the Effects of Vaccination and Different Wolbachia Strains on Dengue Transmission Dynamics. Mathematics. 2022; 10(12):2075. https://0-doi-org.brum.beds.ac.uk/10.3390/math10122075
Chicago/Turabian StyleNdii, Meksianis Z., Lazarus Kalvein Beay, Nursanti Anggriani, Karolina N. Nukul, and Bertha S. Djahi. 2022. "Estimating the Time Reproduction Number in Kupang City Indonesia, 2016–2020, and Assessing the Effects of Vaccination and Different Wolbachia Strains on Dengue Transmission Dynamics" Mathematics 10, no. 12: 2075. https://0-doi-org.brum.beds.ac.uk/10.3390/math10122075