
Spatial Computing in Modular Spiking Neural Networks with a Robotic Embodiment
 , , and
, , and 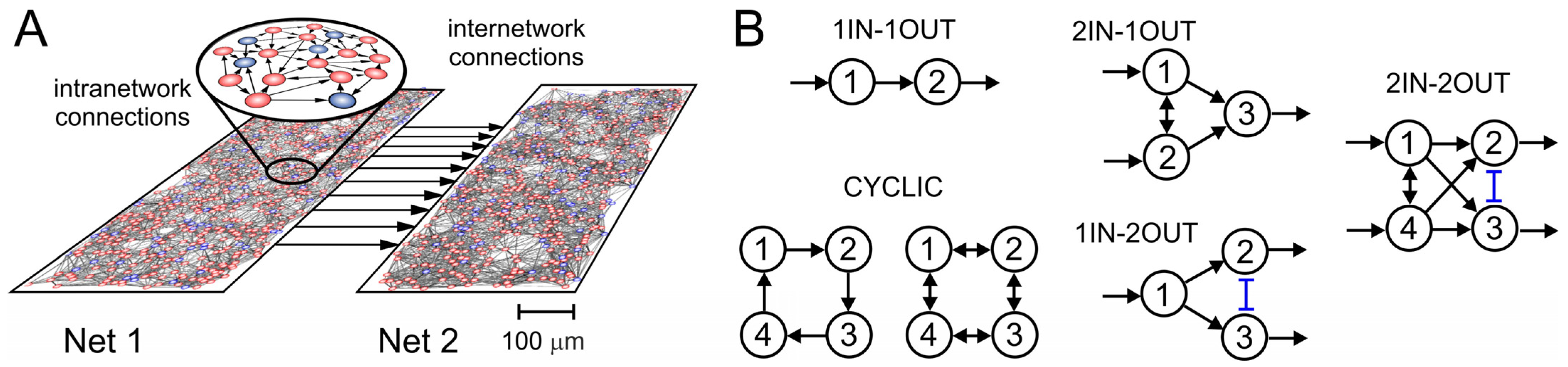{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Local learning mechanisms (e.g., STDP) potentiates the shortest neural pathways and depresses alternative longer pathways at the global network scale.
2. Models and Methods
3. Results
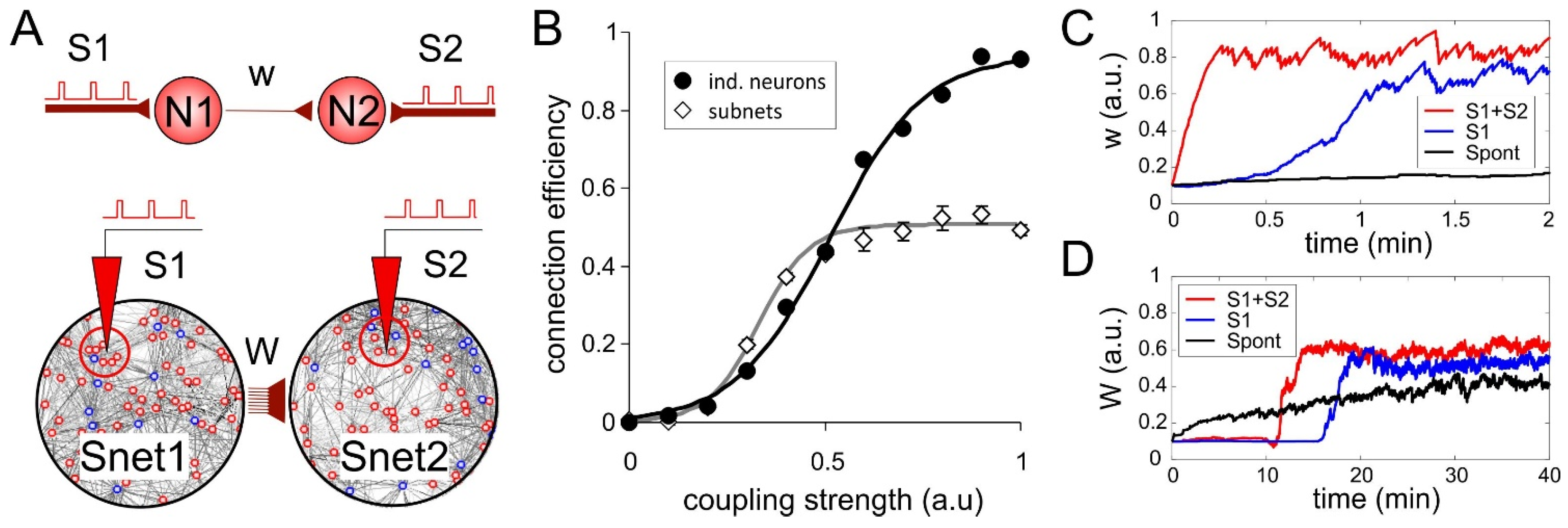
3.1. Self-Reinforcing of Internetwork Couplings in Neural Circuits
3.2. The Shortest Pathway Rule
- Spontaneous (without stimuli) potentiation of connections (Figure 2C,D);
- Potentiation of connections when only the conditional stimulus is applied (no association, Figure 2C,D);
- The lack of a mechanism for depressing an association when it becomes irrelevant (e.g., when the conditional stimulus is not supported by an unconditional one).
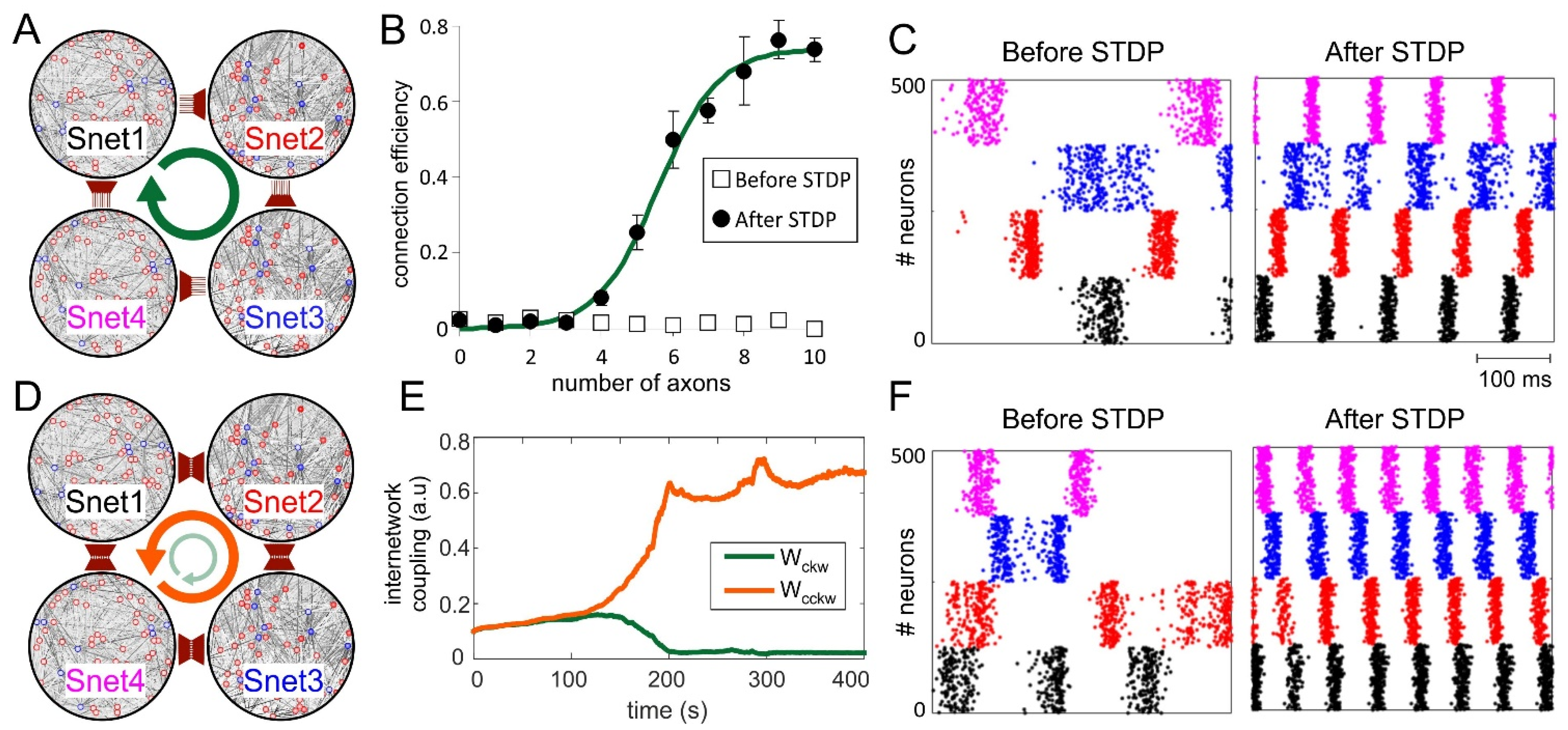
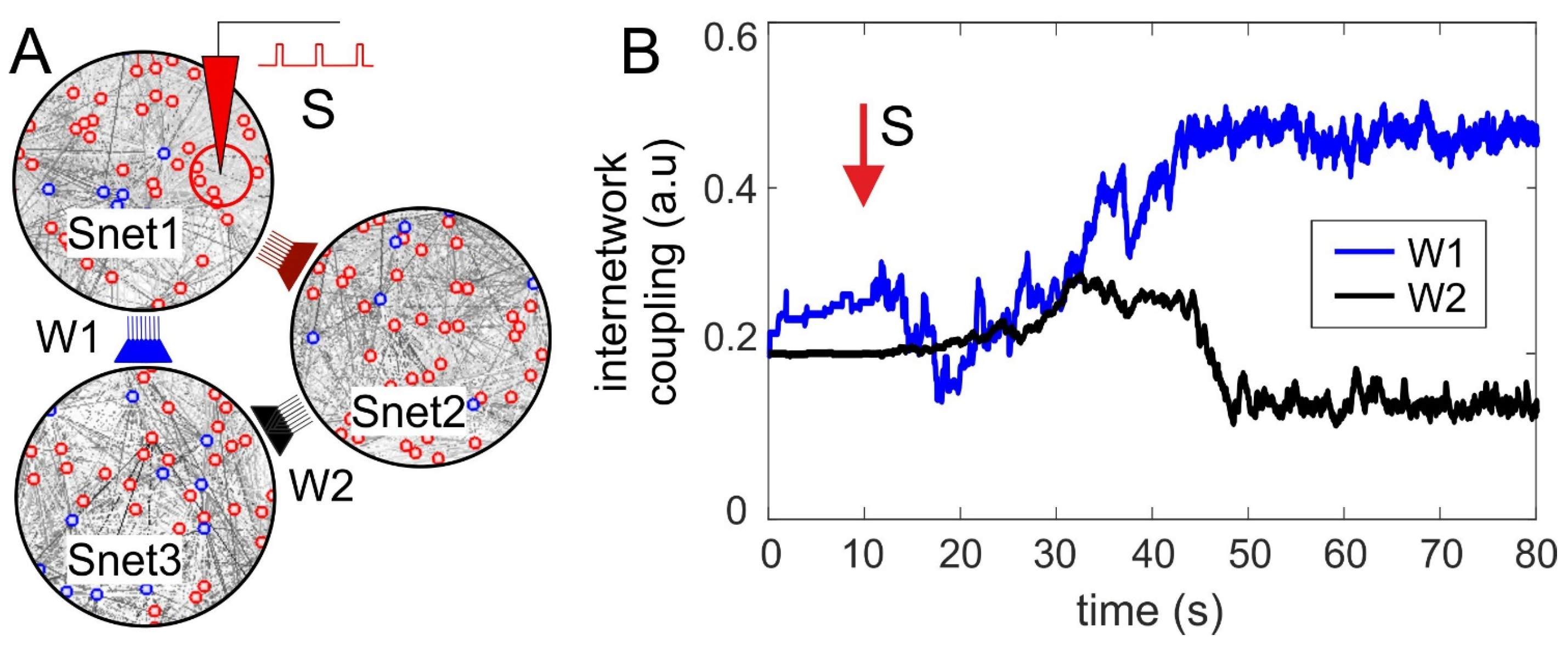
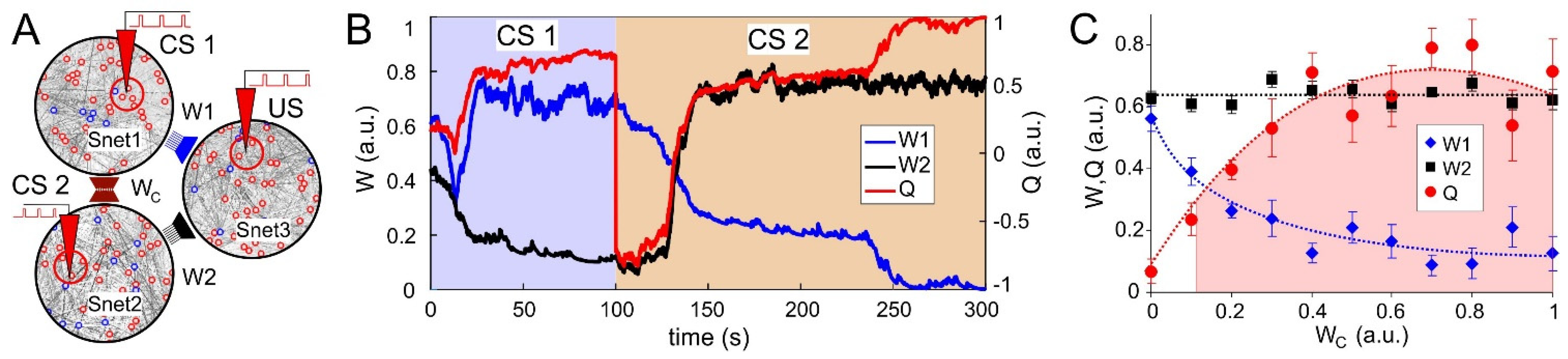
3.3. Synaptic Competition in SNNs
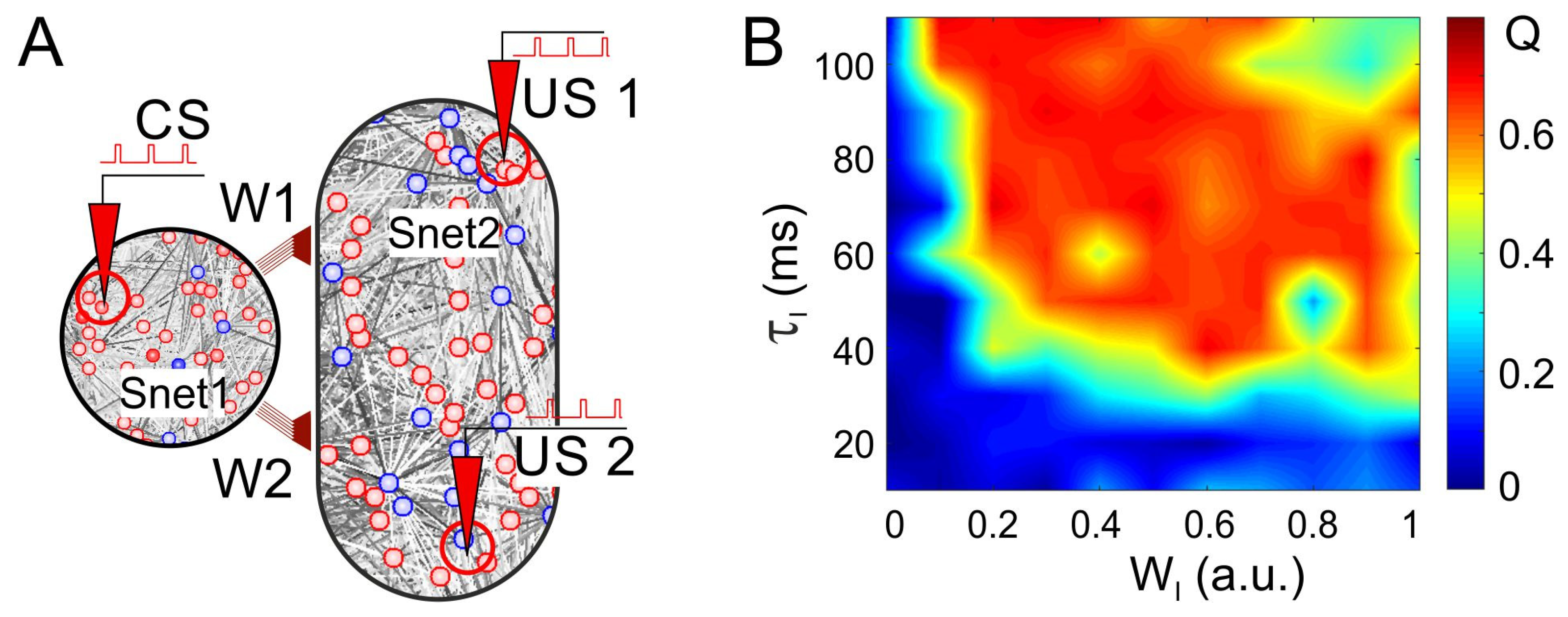
3.4. Neuronal Competition of SNN Outputs
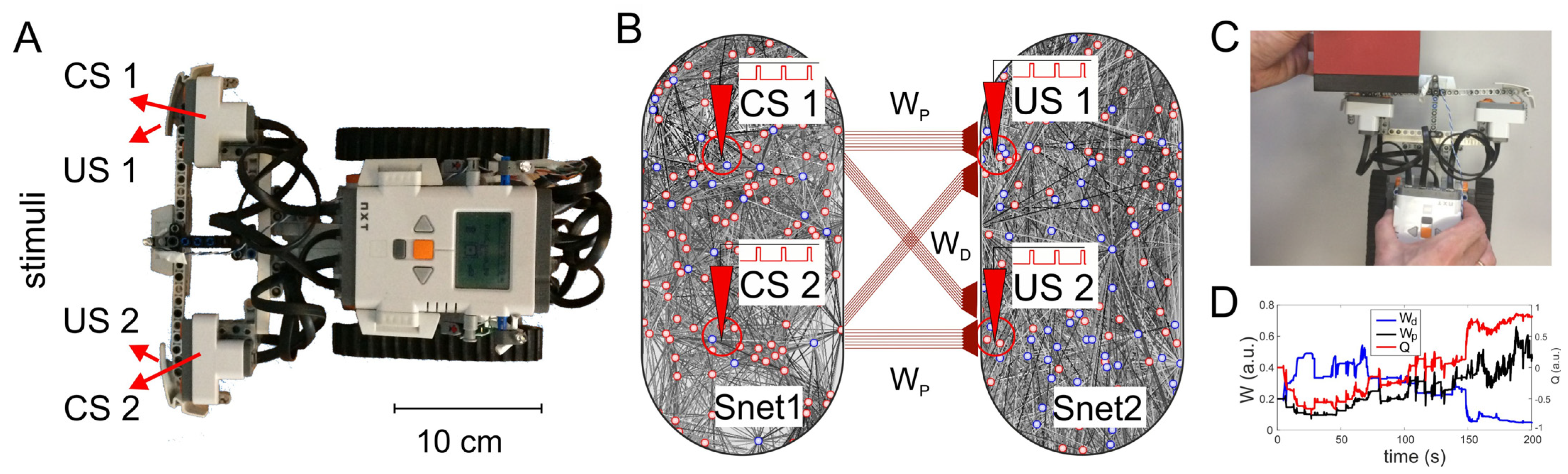
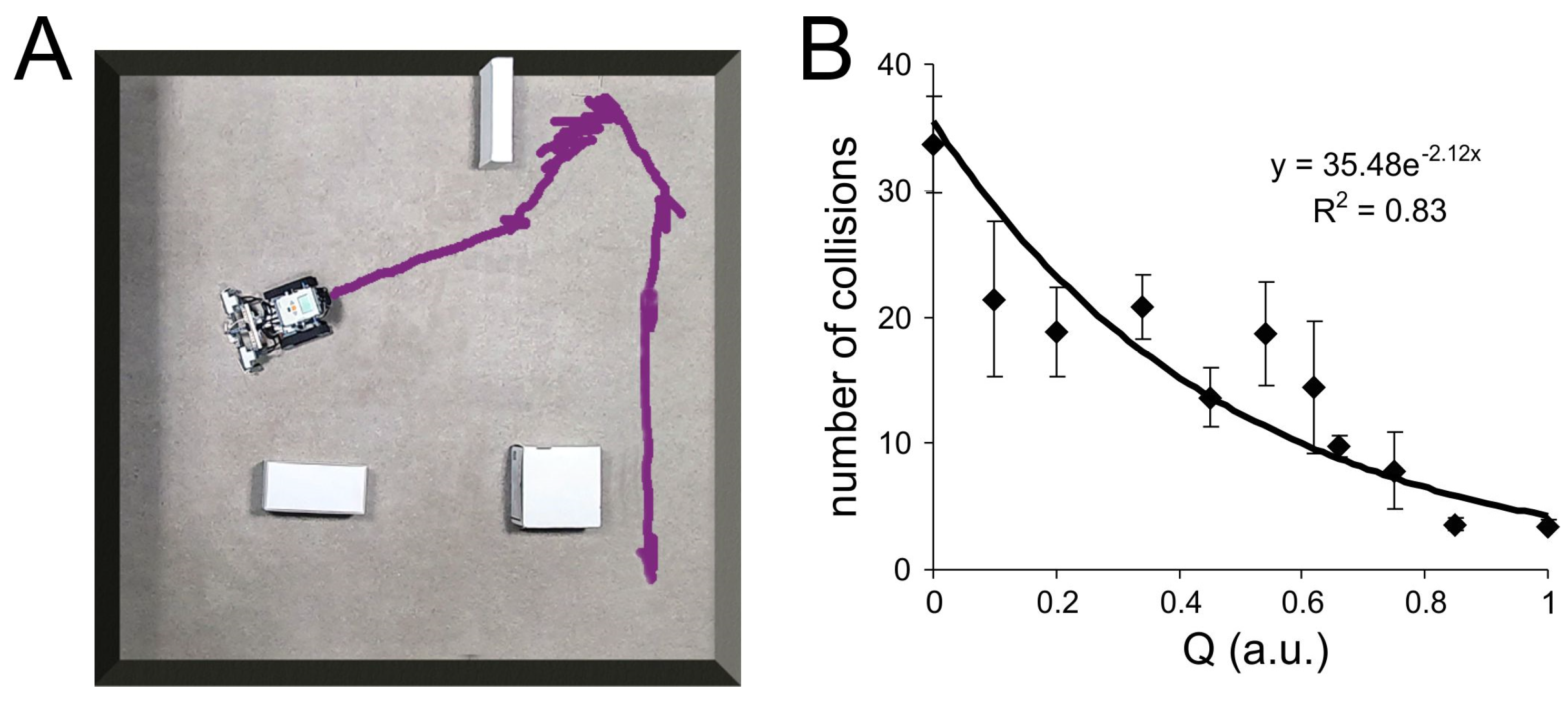
3.5. Robotic Embodiment of Associative Learning
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Potter, S.M.; Fraser, S.E.; Pine, J. Animat in a petri dish: Cultured neural networks for studying neural computation. In Proceedings of the 4th Joint Symposium on Neural Computation, San Diego, CA, USA, 17 May 1997; pp. 167–174. [Google Scholar]
- Pamies, D.; Hartung, T.; Hogberg, H.T. Biological and medical applications of a brain-on-a-chip. Exp. Biol. Med. 2014, 239, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.A.; Wilson, S.W. From animals to animats: Proceedings of the First International Conference on Simulation of Adaptive Behavior; MIT Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Reger, B.D.; Fleming, K.M.; Sanguineti, V.; Alford, S.; Mussa-Ivaldi, F.A. Connecting brains to robots: An artificial body for studying the computational properties of neural tissues. Artif. Life 2000, 6, 307–324. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, B.C. Building a brain on a chip. In Proceedings of the 30th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Vancouver, BC, Canada, 20–25 August 2008; pp. 1604–1606. [Google Scholar]
- Brofiga, M.; Pisano, M.; Raiteri, R.; Massobrio, P. On the road to the brain-on-a-chip: A review on strategies, methods, and applications. J. Neural Eng. 2021, 18, 41005. [Google Scholar] [CrossRef]
- Forro, C.; Caron, D.; Angotzi, G.N.; Gallo, V.; Berdondini, L.; Santoro, F.; Palazzolo, G.; Panuccio, G. Electrophysiology read-out tools for brain-on-chip biotechnology. Micromachines 2021, 12, 124. [Google Scholar] [CrossRef]
- Maoz, B.M. Brain-on-a-Chip: Characterizing the next generation of advanced in vitro platforms for modeling the central nervous system. APL Bioeng. 2021, 5, 30902. [Google Scholar] [CrossRef]
- Aaser, P.; Knudsen, M.; Huse Ramstad, O.; van de Wijdeven, R.; Nichele, S.; Sandvig, I.; Tufte, G.; Bauer, U.S.; Halaas, Ø.; Hendseth, S.; et al. Towards making a cyborg: A closed-loop reservoir-neuro system. In Proceedings of the European Conference on Artificial Life; Knibbe, C., Beslon, G., Parsons, D.P., Misevic, D., Rouzaud-Cornabas, J., Bredèche, N., Hassas, S., Simonin, O., Soula, H., Eds.; MIT Press: Lyon, France, 2017; Volume 2017, pp. 430–437. [Google Scholar]
- Bakkum, D.J.; Chao, Z.C.; Potter, S.M. Spatio-temporal electrical stimuli shape behavior of an embodied cortical network in a goal-directed learning task. J. Neural Eng. 2008, 5, 310. [Google Scholar] [CrossRef] [Green Version]
- Shahaf, G.; Eytan, D.; Gal, A.; Kermany, E.; Lyakhov, V.; Zrenner, C.; Marom, S. Order-based representation in random networks of cortical neurons. PLoS Comput. Biol. 2008, 4, e1000228. [Google Scholar] [CrossRef] [Green Version]
- Kagan, B.J.; Kitchen, A.C.; Tran, N.T.; Habibollahi, F.; Khajehnejad, M.; Parker, B.J.; Bhat, A.; Rollo, B.; Razi, A.; Friston, K.J. In vitro neurons learn and exhibit sentience when embodied in a simulated game-world. Neuron 2022, 110, 3952–3969.e8. [Google Scholar] [CrossRef]
- Dauth, S.; Maoz, B.M.; Sheehy, S.P.; Hemphill, M.A.; Murty, T.; Macedonia, M.K.; Greer, A.M.; Budnik, B.; Parker, K.K. Neurons derived from different brain regions are inherently different in vitro: A novel multiregional brain-on-a-chip. J. Neurophysiol. 2016, 117, 1320–1341. [Google Scholar] [CrossRef] [PubMed]
- Pimashkin, A.; Gladkov, A.; Mukhina, I.; Kazantsev, V. Adaptive enhancement of learning protocol in hippocampal cultured networks grown on multielectrode arrays. Front. Neural Circuits 2013, 7, 87. [Google Scholar] [CrossRef]
- Pimashkin, A.; Gladkov, A.; Agrba, E.; Mukhina, I.; Kazantsev, V. Selectivity of stimulus induced responses in cultured hippocampal networks on microelectrode arrays. Cogn. Neurodyn. 2016, 10, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.M.; Rhee, S.W.; Tu, C.H.; Cribbs, D.H.; Cotman, C.W.; Jeon, N.L. Microfluidic multicompartment device for neuroscience research. Langmuir 2003, 19, 1551–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibey, R.; Rojo Arias, J.E.; Striebel, J.; Busskamp, V. Microfluidics for Neuronal cell and circuit engineering. Chem. Rev. 2022, 122, 14842–14880. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Alagapan, S.; Franca, E.; Brewer, G.J.; Wheeler, B.C. Propagation of action potential activity in a predefined microtunnel neural network. J. Neural Eng. 2011, 8, 46031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Alagapan, S.; Franca, E.; Leondopulos, S.S.; DeMarse, T.B.; Brewer, G.J.; Wheeler, B.C. An in vitro method to manipulate the direction and functional strength between neural populations. Front. Neural Circuits 2015, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- DeMarse, T.B.; Pan, L.; Alagapan, S.; Brewer, G.J.; Wheeler, B.C. Feed-forward propagation of temporal and rate information between cortical populations during coherent activation in engineered in vitro networks. Front. Neural Circuits 2016, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- le Feber, J.; Postma, W.; de Weerd, E.; Weusthof, M.; Rutten, W.L.C. Barbed channels enhance unidirectional connectivity between neuronal networks cultured on multi electrode arrays. Front. Neurosci. 2015, 9, 412. [Google Scholar] [CrossRef] [Green Version]
- Malishev, E.; Pimashkin, A.; Arseniy, G.; Pigareva, Y.; Bukatin, A.; Kazantsev, V.; Mukhina, I.; Dubina, M. Microfluidic device for unidirectional axon growth. J. Phys. Conf. Ser. 2015, 643, 012025. [Google Scholar] [CrossRef] [Green Version]
- Gladkov, A.; Pigareva, Y.; Kutyina, D.; Kolpakov, V.; Bukatin, A.; Mukhina, I.; Kazantsev, V.; Pimashkin, A. Design of cultured neuron networks in vitro with predefined connectivity using asymmetric microfluidic channels. Sci. Rep. 2017, 7, 15625. [Google Scholar] [CrossRef] [Green Version]
- Forró, C.; Thompson-Steckel, G.; Weaver, S.; Weydert, S.; Ihle, S.; Dermutz, H.; Aebersold, M.J.; Pilz, R.; Demkó, L.; Vörös, J. Modular microstructure design to build neuronal networks of defined functional connectivity. Biosens. Bioelectron. 2018, 122, 75–87. [Google Scholar] [CrossRef]
- Na, S.; Kang, M.; Bang, S.; Park, D.; Kim, J.; Sim, S.J.; Chang, S.; Jeon, N.L. Microfluidic neural axon diode. Technology 2016, 4, 240–248. [Google Scholar] [CrossRef]
- Dworak, B.J.; Wheeler, B.C. Novel MEA platform with PDMS microtunnels enables the detection of action potential propagation from isolated axons in culture. Lab. Chip 2009, 9, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, S.; Park, S.I.; Choe, Y.; Li, J.; Han, A. A microchip for quantitative analysis of CNS axon growth under localized biomolecular treatments. J. Neurosci. Methods 2014, 221, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Wijdeven, R.; Ramstad, O.H.; Bauer, U.S.; Halaas, Ø.; Sandvig, A.; Sandvig, I. Structuring a multi-nodal neural network in vitro within a novel design microfluidic chip. Biomed. Microdevices 2018, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Furukawa, T.; Asahina, T.; Shimba, K.; Kotani, K.; Jimbo, Y. Coupling of in vitro neocortical-hippocampal coculture bursts induces different spike shythms in individual networks. Front. Neurosci. 2022, 16, 873664. [Google Scholar] [CrossRef] [PubMed]
- Callegari, F.; Brofiga, M.; Poggio, F.; Massobrio, P. Stimulus-evoked activity modulation of in vitro engineered cortical and hippocampal networks. Micromachines 2022, 13, 1212. [Google Scholar] [CrossRef]
- Lobov, S.A.; Zhuravlev, M.O.; Makarov, V.A.; Kazantsev, V.B. Noise enhanced signaling in STDP driven spiking-neuron network. Math. Model. Nat. Phenom. 2017, 12, 109–124. [Google Scholar] [CrossRef]
- Lobov, S.; Balashova, K.; Makarov, V.A.; Kazantsev, V. Competition of spike-conducting pathways in STDP driven neural networks. In Proceedings of the 5th International Congress on Neurotechnology, Electronics and Informatics (NEUROTECHNIX 2017); SciTePress: Setúbal, Portugalija, 2017; pp. 15–21. [Google Scholar]
- Lobov, S.A.; Mikhaylov, A.N.; Shamshin, M.; Makarov, V.A.; Kazantsev, V.B. Spatial properties of STDP in a self-learning spiking neural network enable controlling a mobile robot. Front. Neurosci. 2020, 14, 88. [Google Scholar] [CrossRef] [PubMed]
- Markram, H.; Gerstner, W.; Sjöström, P.J. A history of spike-timing-dependent plasticity. Front. Synaptic Neurosci. 2011, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markram, H.; Lübke, J.; Frotscher, M.; Sakmann, B. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 1997, 275, 213–215. [Google Scholar] [CrossRef]
- Bi, G.Q.; Poo, M.M. Synaptic modifications in cultured hippocampal neurons: Dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci. 1998, 18, 10464–10472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöström, P.J.; Turrigiano, G.G.; Nelson, S.B. Rate, timing, and cooperativity jointly determine cortical synaptic plasticity. Neuron 2001, 32, 1149–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jäckel, D.; Bakkum, D.J.; Russell, T.L.; Müller, J.; Radivojevic, M.; Frey, U.; Franke, F.; Hierlemann, A. Combination of high-density microelectrode array and patch clamp recordings to enable studies of multisynaptic integration. Sci. Rep. 2017, 7, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigby, M.; Anthonisen, M.; Chua, X.Y.; Kaplan, A.; Fournier, A.E.; Grütter, P. Building an artificial neural network with neurons. AIP Adv. 2019, 9, 075009. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Networks 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Which model to use for cortical spiking neurons? IEEE Trans. Neural Networks 2004, 15, 1063–1070. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting; The MIT Press: Cambridge, MA, USA; London, UK, 2007. [Google Scholar]
- Lobov, S.A.; Chernyshov, A.V.; Krilova, N.P.; Shamshin, M.O.; Kazantsev, V.B. Competitive learning in a spiking neural network: Towards an intelligent pattern classifier. Sensors 2020, 20, 500. [Google Scholar] [CrossRef] [Green Version]
- Makarov, V.A.; Lobov, S.A.; Shchanikov, S.; Mikhaylov, A.; Kazantsev, V.B. Toward reflective spiking neural networks exploiting memristive devices. Front. Comput. Neurosci. 2022, 16, 859874. [Google Scholar] [CrossRef]
- Tsodyks, M.; Pawelzik, K.; Markram, H. Neural networks with dynamic synapses. Neural Comput. 1998, 10, 821–835. [Google Scholar] [CrossRef]
- Morrison, A.; Diesmann, M.; Gerstner, W. Phenomenological models of synaptic plasticity based on spike timing. Biol. Cybern. 2008, 98, 459–478. [Google Scholar] [CrossRef]
- Song, S.; Miller, K.D.; Abbott, L.F. Competitive Hebbian learning through spike-timing-dependent synaptic plasticity. Nat. Neurosci. 2000, 3, 919. [Google Scholar] [CrossRef] [PubMed]
- Lobov, S.A.; Zharinov, A.I.; Semenova, O.; Kazantsev, V.B. Topological classification of population activity in spiking neural network. In Proceedings of SPIE: The Saratov Fall Meeting 2020: Computations and Data Analysis: From Molecular Processes to Brain Functions; Postnov, D.E., Ed.; SPIE: Bellingham, DC, USA, 2021; Volume 11847, pp. 1–6. [Google Scholar]
- Spiking Neurosimulator NeuroNet with a User-Friendly Graphical Interface. Available online: http://spneuro.net/ (accessed on 29 December 2022).
- Wagenaar, D.A.; Pine, J.; Potter, S.M. An extremely rich repertoire of bursting patterns during the development of cortical cultures. BMC Neurosci. 2006, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Chiappalone, M.; Novellino, A.; Vajda, I.; Vato, A.; Martinoia, S.; van Pelt, J. Burst detection algorithms for the analysis of spatio-temporal patterns in cortical networks of neurons. Neurocomputing 2005, 65–66, 653–662. [Google Scholar] [CrossRef]
- Stegenga, J.; Le Feber, J.; Marani, E.; Rutten, W.L.C. Analysis of cultured neuronal networks using intraburst firing characteristics. IEEE Trans. Biomed. Eng. 2008, 55, 1382–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigareva, Y.; Gladkov, A.; Kolpakov, V.; Mukhina, I.; Bukatin, A.; Kazantsev, V.B.; Pimashkin, A. Experimental platform to study spiking pattern propagation in modular networks in vitro. Brain Sci. 2021, 11, 717. [Google Scholar] [CrossRef]
- Pan, L.; Alagapan, S.; Franca, E.; DeMarse, T.; Brewer, G.J.; Wheeler, B.C. Large extracellular spikes recordable from axons in microtunnels. IEEE Trans. Neural Syst. Rehabil. Eng. 2014, 22, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Geramifard, N.; Lawson, J.; Cogan, S.F.; Black, B.J. A novel 3D helical microelectrode array for in vitro extracellular action potential recording. Micromachines 2022, 13, 1692. [Google Scholar] [CrossRef] [PubMed]
- Keren, H.; Marom, S. Long-range synchrony and emergence of neural reentry. Sci. Rep. 2016, 6, 36837. [Google Scholar] [CrossRef] [Green Version]
- Chua, L.O.; Kang, S.M. Memristive devices and systems. Proc. IEEE 1976, 64, 209–223. [Google Scholar] [CrossRef]
- Strukov, D.B.; Snider, G.S.; Stewart, D.R.; Williams, R.S. The missing memristor found. Nature 2008, 453, 80–83. [Google Scholar] [CrossRef]
- Dias, C.; Castro, D.; Aroso, M.; Ventura, J.; Aguiar, P. A memristor-based neuromodulation device for real-time monitoring and adaptive control of in vitro neuronal populations. ACS Appl. Electron. Mater. 2022, 4, 2380–2387. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylov, A.; Pimashkin, A.; Pigareva, Y.; Gerasimova, S.A.; Lobov, S.; Gryaznov, E.; Talanov, M.; Lavrov, I.; Demin, V.; Erokhin, V.; et al. Neurohybrid memristive CMOS-integrated systems for biosensors and neuroprostetics. Front. Neurosci. 2020, 14, 358. [Google Scholar] [CrossRef] [PubMed]
- Juzekaeva, E.; Nasretdinov, A.; Battistoni, S.; Berzina, T.; Iannotta, S.; Khazipov, R.; Erokhin, V.; Mukhtarov, M. Coupling cortical neurons through electronic memristive synapse. Adv. Mater. Technol. 2019, 4, 1800350. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobov, S.A.; Mikhaylov, A.N.; Berdnikova, E.S.; Makarov, V.A.; Kazantsev, V.B. Spatial Computing in Modular Spiking Neural Networks with a Robotic Embodiment. Mathematics 2023, 11, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/math11010234
Lobov SA, Mikhaylov AN, Berdnikova ES, Makarov VA, Kazantsev VB. Spatial Computing in Modular Spiking Neural Networks with a Robotic Embodiment. Mathematics. 2023; 11(1):234. https://0-doi-org.brum.beds.ac.uk/10.3390/math11010234
Chicago/Turabian StyleLobov, Sergey A., Alexey N. Mikhaylov, Ekaterina S. Berdnikova, Valeri A. Makarov, and Victor B. Kazantsev. 2023. "Spatial Computing in Modular Spiking Neural Networks with a Robotic Embodiment" Mathematics 11, no. 1: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/math11010234