
SWATH-Based Comprehensive Determination of the Localization of Apical and Basolateral Membrane Proteins Using Mouse Liver as a Model Tissue


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Separation of Basolateral (Sinusoidal) and Apical (Canalicular) Plasma Membranes of Mouse Liver by Density-Gradient Ultracentrifugation
2.3. Sample Preparation for SWATH-Based Quantitative Proteomics
2.4. LC-MS/MS Measurement for SWATH-Based Quantitative Proteomics
2.5. Data Analysis for SWATH-Based Quantitative Proteomics
3. Results
3.1. Validation of the SWATH-Based Comprehensive Determinations of Basolateral and Apical Plasma Membrane Localizations
3.2. SWATH-Based Comprehensive Determinations of Basolateral and Apical Plasma Membrane Localizations in Mouse Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debacker, A.J.; Voutila, J.; Catley, M.; Blakey, D.; Habib, N. Delivery of Oligonucleotides to the Liver with GalNAc: From Research to Registered Therapeutic Drug. Mol. Ther. 2020, 28, 1759–1771. [Google Scholar] [CrossRef] [PubMed]
- Lohade, A.A.; Jain, R.R.; Iyer, K.; Roy, S.K.; Shimpi, H.H.; Pawar, Y.; Rajan, M.G.; Menon, M.D. A Novel Folate-Targeted Nanoliposomal System of Doxorubicin for Cancer Targeting. AAPS PharmSciTech 2016, 17, 1298–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, L.C.; Navarro, P.; Tate, S.; Rost, H.; Selevsek, N.; Reiter, L.; Bonner, R.; Aebersold, R. Targeted data extraction of the MS/MS spectra generated by data-independent acquisition: A new concept for consistent and accurate proteome analysis. Mol. Cell. Proteom. 2012, 11, O111.016717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, Y.; Goto, R.; Takeuchi, H.; Luczak, M.; Usui, T.; Tachikawa, M.; Terasaki, T. Abundant Expression of OCT2, MATE1, OAT1, OAT3, PEPT2, BCRP, MDR1, and xCT Transporters in Blood-Arachnoid Barrier of Pig and Polarized Localizations at CSF- and Blood-Facing Plasma Membranes. Drug Metab. Dispos. 2020, 48, 135–145. [Google Scholar] [CrossRef]
- Kamiie, J.; Ohtsuki, S.; Iwase, R.; Ohmine, K.; Katsukura, Y.; Yanai, K.; Sekine, Y.; Uchida, Y.; Ito, S.; Terasaki, T. Quantitative atlas of membrane transporter proteins: Development and application of a highly sensitive simultaneous LC/MS/MS method combined with novel in-silico peptide selection criteria. Pharm. Res. 2008, 25, 1469–1483. [Google Scholar] [CrossRef]
- Uchida, Y.; Higuchi, T.; Shirota, M.; Kagami, S.; Saigusa, D.; Koshiba, S.; Yasuda, J.; Tamiya, G.; Kuriyama, S.; Kinoshita, K.; et al. Identification and Validation of Combination Plasma Biomarker of Afamin, Fibronectin and Sex Hormone-Binding Globulin to Predict Pre-eclampsia. Biol. Pharm. Bull. 2021, 44, 804–815. [Google Scholar] [CrossRef]
- Uchida, Y.; Sasaki, H.; Terasaki, T. Establishment and validation of highly accurate formalin-fixed paraffin-embedded quantitative proteomics by heat-compatible pressure cycling technology using phase-transfer surfactant and SWATH-MS. Sci. Rep. 2020, 10, 11271. [Google Scholar] [CrossRef]
- Meier, P.J.; Sztul, E.S.; Reuben, A.; Boyer, J.L. Structural and functional polarity of canalicular and basolateral plasma membrane vesicles isolated in high yield from rat liver. J. Cell Biol. 1984, 98, 991–1000. [Google Scholar] [CrossRef]
- Uchida, Y.; Tachikawa, M.; Obuchi, W.; Hoshi, Y.; Tomioka, Y.; Ohtsuki, S.; Terasaki, T. A study protocol for quantitative targeted absolute proteomics (QTAP) by LC-MS/MS: Application for inter-strain differences in protein expression levels of transporters, receptors, claudin-5, and marker proteins at the blood-brain barrier in ddY, FVB, and C57BL/6J mice. Fluids Barriers CNS 2013, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Sumiya, T.; Tachikawa, M.; Yamakawa, T.; Murata, S.; Yagi, Y.; Sato, K.; Stephan, A.; Ito, K.; Ohtsuki, S.; et al. Involvement of Claudin-11 in Disruption of Blood-Brain, -Spinal Cord, and -Arachnoid Barriers in Multiple Sclerosis. Mol. Neurobiol. 2019, 56, 2039–2056. [Google Scholar] [CrossRef] [PubMed]
- Elkjaer, M.; Vajda, Z.; Nejsum, L.N.; Kwon, T.; Jensen, U.B.; Amiry-Moghaddam, M.; Frokiaer, J.; Nielsen, S. Immunolocalization of AQP9 in liver, epididymis, testis, spleen, and brain. Biochem. Biophys. Res. Commun. 2000, 276, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Sakar, Y.; Nazaret, C.; Letteron, P.; Ait Omar, A.; Avenati, M.; Viollet, B.; Ducroc, R.; Bado, A. Positive regulatory control loop between gut leptin and intestinal GLUT2/GLUT5 transporters links to hepatic metabolic functions in rodents. PLoS ONE 2009, 4, e7935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, C.D.; Aleksunes, L.M. Xenobiotic, bile acid, and cholesterol transporters: Function and regulation. Pharmacol. Rev. 2010, 62, 1–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quondamatteo, F.; Krick, W.; Hagos, Y.; Kruger, M.H.; Neubauer-Saile, K.; Herken, R.; Ramadori, G.; Burckhardt, G.; Burckhardt, B.C. Localization of the sulfate/anion exchanger in the rat liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1075–G1081. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, S.; Tachikawa, M.; Akanuma, S.; Fujinawa, J.; Hosoya, K. Involvement of gamma-aminobutyric acid transporter 2 in the hepatic uptake of taurine in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G291–G297. [Google Scholar] [CrossRef] [Green Version]
- Kirat, D.; Inoue, H.; Iwano, H.; Yokota, H.; Taniyama, H.; Kato, S. Monocarboxylate transporter 1 (MCT1) in the liver of pre-ruminant and adult bovines. Vet. J. 2007, 173, 124–130. [Google Scholar] [CrossRef]
- Garcia, C.K.; Brown, M.S.; Pathak, R.K.; Goldstein, J.L. cDNA cloning of MCT2, a second monocarboxylate transporter expressed in different cells than MCT1. J. Biol. Chem. 1995, 270, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- Preitner, F.; Bonny, O.; Laverriere, A.; Rotman, S.; Firsov, D.; Da Costa, A.; Metref, S.; Thorens, B. Glut9 is a major regulator of urate homeostasis and its genetic inactivation induces hyperuricosuria and urate nephropathy. Proc. Natl. Acad. Sci. USA 2009, 106, 15501–15506. [Google Scholar] [CrossRef] [Green Version]
- Benkoel, L.; Benoliel, A.M.; Brisse, J.; Sastre, B.; Bongrand, P.; Chamlian, A. Immunocytochemical study of NA+ K(+)-ATPase alpha 1 and beta 1 subunits in human and rat normal hepatocytes using confocal microscopy. Cell Mol. Biol. 1995, 41, 499–504. [Google Scholar]
- Matsuzaka, Y.; Hayashi, H.; Kusuhara, H. Impaired Hepatic Uptake by Organic Anion-Transporting Polypeptides Is Associated with Hyperbilirubinemia and Hypercholanemia in Atp11c Mutant Mice. Mol. Pharmacol. 2015, 88, 1085–1092. [Google Scholar] [CrossRef] [Green Version]
- Macias, R.I.; Hierro, C.; de Juan, S.C.; Jimenez, F.; Gonzalez-San Martin, F.; Marin, J.J. Hepatic expression of sodium-dependent vitamin C transporters: Ontogeny, subtissular distribution and effect of chronic liver diseases. Br. J. Nutr. 2011, 106, 1814–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.; Villegas, C.J.; Jiang, J.X. Differential regulation of amino acid transporter SNAT3 by insulin in hepatocytes. J. Biol. Chem. 2005, 280, 26055–26062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, M.; Nakanishi, T.; Shibue, Y.; Kobayashi, D.; Moseley, R.H.; Shirasaka, Y.; Tamai, I. Hepatic uptake of gamma-butyrobetaine, a precursor of carnitine biosynthesis, in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G681–G686. [Google Scholar] [CrossRef] [PubMed]
- Oyamada, M.; Mori, M. Immunohistochemical demonstration of tubulin and actin in rat hepatocytes in situ using a perfusion extraction-fixation procedure. J. Histochem. Cytochem. 1985, 33, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, S.H.; Roche, P.A.; Anderson, H.A.; van Ijzendoorn, S.C.; Zhang, M.; Mostov, K.E.; Weimbs, T. Targeting of SNAP-23 and SNAP-25 in polarized epithelial cells. J. Biol. Chem. 1998, 273, 3422–3430. [Google Scholar] [CrossRef] [Green Version]
- Braiterman, L.T.; Heffernan, S.; Nyasae, L.; Johns, D.; See, A.P.; Yutzy, R.; McNickle, A.; Herman, M.; Sharma, A.; Naik, U.P.; et al. JAM-A is both essential and inhibitory to development of hepatic polarity in WIF-B cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G576–G588. [Google Scholar] [CrossRef] [Green Version]
- Stamatoglou, S.C.; Sullivan, K.H.; Johansson, S.; Bayley, P.M.; Burdett, I.D.; Hughes, R.C. Localization of two fibronectin-binding glycoproteins in rat liver and primary hepatocytes. Co-distribution in vitro of integrin (alpha 5 beta 1) and non-integrin (AGp110) receptors in cell-substratum adhesion sites. J. Cell Sci. 1990, 97, 595–606. [Google Scholar] [CrossRef]
- Chaubey, P.M.; Hofstetter, L.; Roschitzki, B.; Stieger, B. Proteomic Analysis of the Rat Canalicular Membrane Reveals Expression of a Complex System of P4-ATPases in Liver. PLoS ONE 2016, 11, e0158033. [Google Scholar] [CrossRef] [Green Version]
- Treyer, A.; Musch, A. Hepatocyte polarity. Compr. Physiol. 2013, 3, 243–287. [Google Scholar] [CrossRef] [Green Version]
- Bon, C.; Hofer, T.; Bousquet-Melou, A.; Davies, M.R.; Krippendorff, B.F. Capacity limits of asialoglycoprotein receptor-mediated liver targeting. MAbs 2017, 9, 1360–1369. [Google Scholar] [CrossRef] [Green Version]
- Kubo, Y.; Ohtsuki, S.; Uchida, Y.; Terasaki, T. Quantitative Determination of Luminal and Abluminal Membrane Distributions of Transporters in Porcine Brain Capillaries by Plasma Membrane Fractionation and Quantitative Targeted Proteomics. J. Pharm. Sci. 2015, 104, 3060–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morre, D.J.; Hammond, T. Isolation of renal brush borders. Curr. Protoc. Cell Biol. 2007, Chapter 3. 3.26.1–3.26.14. [Google Scholar] [CrossRef] [PubMed]
- Pinches, S.A.; Gribble, S.M.; Beechey, R.B.; Ellis, A.; Shaw, J.M.; Shirazi-Beechey, S.P. Preparation and characterization of basolateral membrane vesicles from pig and human colonocytes: The mechanism of glucose transport. Biochem J. 1993, 294, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilden, S.A.; Johns, C.A.; Guggino, W.B.; Madias, N.E. Techniques for isolation of brush-border and basolateral membrane vesicles from dog kidney cortex. Biochim. Biophys. Acta 1989, 983, 77–81. [Google Scholar] [CrossRef]
- Gustin, M.C.; Goodman, D.B. Isolation of brush-border membrane from the rabbit descending colon epithelium. Partial characterization of a unique K+-activated ATPase. J. Biol. Chem. 1981, 256, 10651–10656. [Google Scholar] [CrossRef]



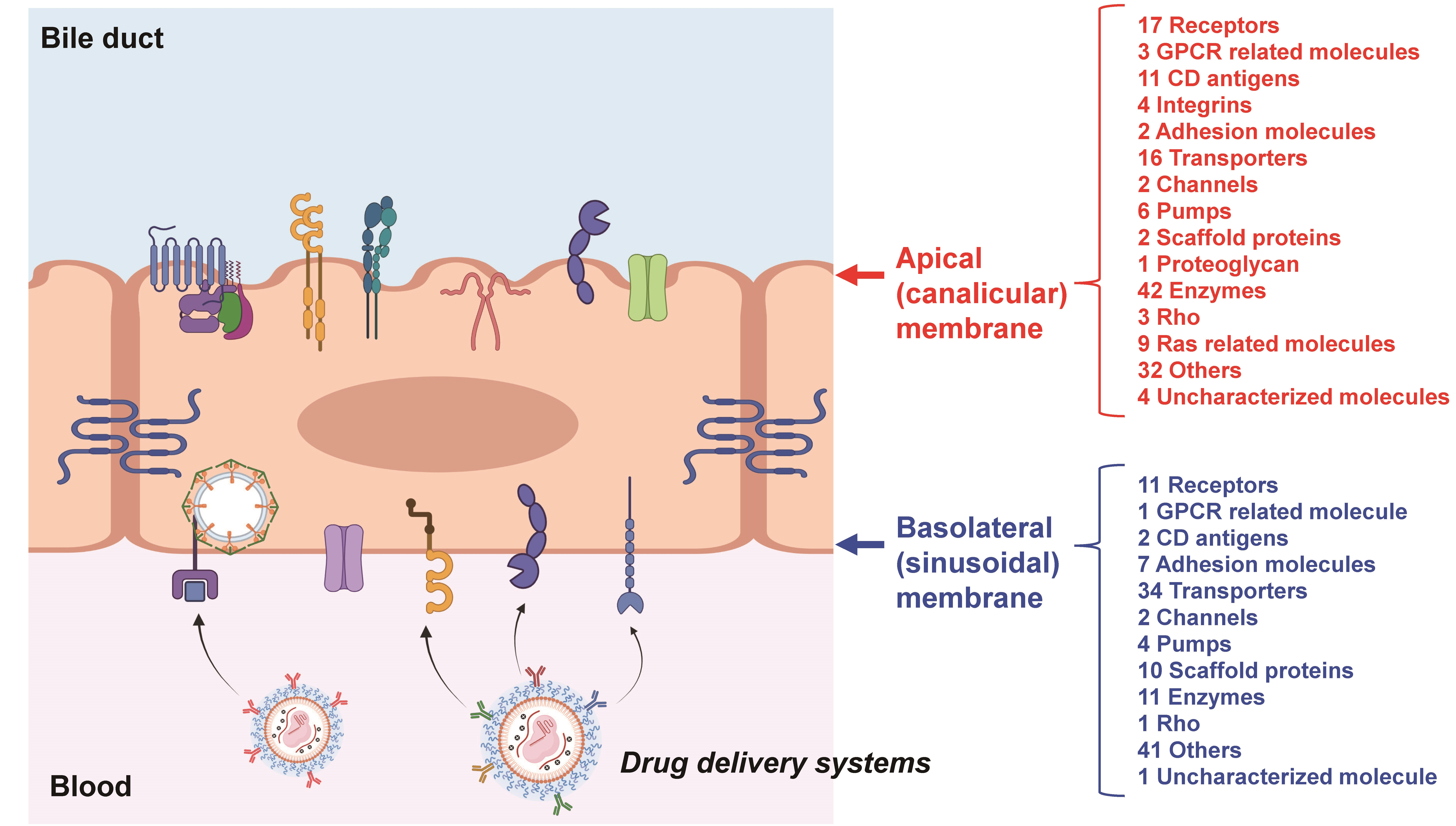{kind=link}
{kind=link}
{kind=link}
| Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Llocalized? (Based on Uniprot) | Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Localized? (Based on Uniprot) | Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Localized? (Based on Uniprot) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Receptors | Channels | Rho | |||||||||
| Folr2 | Q05685 | 0.367 ± 0.010 | Yes | Tmem63a | Q91YT8 | 0.490 ± 0.013 | Yes | Cdc42 | P60766 | 0.501 ± 0.004 | Yes |
| Rpsa | P14206 | 0.407 ± 0.001 | Yes | Aqp1 | Q02013 | 0.694 ± 0.004 | Yes | Rhog | P84096 | 0.526 ± 0.005 | Yes |
| Stra6l | Q9DBN1 | 0.483 ± 0.002 | Yes | Rhob | P62746 | 0.643 ± 0.011 | Yes | ||||
| Fcgr2 | P08101 | 0.483 ± 0.004 | Yes | Pumps | |||||||
| Ptprc | P06800 | 0.496 ± 0.003 | Yes | Atp6v1e1 | P50518 | 0.383 ± 0.010 | Yes | Ras related molecules | |||
| Calcrl | Q9R1W5 | 0.503 ± 0.006 | Yes | Atp6v1b2 | P62814 | 0.513 ± 0.004 | Yes | Rab1A | P62821 | 0.460 ± 0.003 | Potentially |
| Fcer1g | P20491 | 0.518 ± 0.009 | Yes | Tcirg1 | Q9JHF5 | 0.574 ± 0.001 | Potentially | Rab9a | Q9R0M6 | 0.463 ± 0.005 | Yes |
| Npr1 | P18293 | 0.518 ± 0.010 | Potentially | Atp6v1d | P57746 | 0.592 ± 0.008 | Potentially | Rab35 | Q6PHN9 | 0.479 ± 0.002 | Yes |
| P2rx4 | Q9JJX6 | 0.552 ± 0.002 | Potentially | Atp2b4 | Q6Q477 | 0.611 ± 0.006 | Potentially | Rab5a | Q9CQD1 | 0.620 ± 0.008 | Yes |
| Mrc1 | Q61830 | 0.554 ± 0.000 | Yes | Atp6v0a2 | P15920 | 0.690 ± 0.008 | Yes | Rras | P10833 | 0.648 ± 0.004 | Yes |
| Rack1 | P68040 | 0.588 ± 0.002 | Yes | Rab5c | P35278 | 0.653 ± 0.013 | Yes | ||||
| Il6st | Q00560 | 0.607 ± 0.007 | Yes | Scaffold proteins | Rab18 | P35293 | 0.660 ± 0.008 | Yes | |||
| Stab1 | Q8R4Y4 | 0.628 ± 0.001 | Potentially | Cav1 | P49817 | 0.584 ± 0.006 | Yes | Rab5b | P61021 | 0.708 ± 0.012 | Yes |
| Stab2 | Q8R4U0 | 0.634 ± 0.001 | Yes | Msn | P26041 | 0.732 ± 0.003 | Yes | Rab8a | P55258 | 0.733 ± 0.010 | Yes |
| Fcgrt | Q61559 | 0.655 ± 0.003 | Yes | ||||||||
| Pigr | O70570 | 0.715 ± 0.001 | Yes | Proteoglycan | Others | ||||||
| Prlr | Q08501 | 0.746 ± 0.006 | Potentially | Sdc4 | O35988 | 0.340 ± 0.005 | Potentially | Susd2 | Q9DBX3 | 0.437 ± 0.005 | Yes |
| Mreg | Q6NVG5 | 0.437 ± 0.007 | Yes | ||||||||
| GPCR related molecules | Enzymes | Meak7 | Q8K0P3 | 0.458 ± 0.004 | Potentially | ||||||
| Gnai2 | P08752 | 0.449 ± 0.002 | Yes | Dpep1 | P31428 | 0.273 ± 0.006 | Yes | Lamtor1 | Q9CQ22 | 0.470 ± 0.002 | Yes |
| Gpr155 | A2AWR3 | 0.527 ± 0.003 | Potentially | Bst1 | Q64277 | 0.290 ± 0.002 | Yes | Ehd3 | Q9QXY6 | 0.475 ± 0.004 | Yes |
| Gpr182 | P43142 | 0.715 ± 0.010 | Yes | Dpp4 | P28843 | 0.301 ± 0.000 | Yes | Lmbrd1 | Q8K0B2 | 0.523 ± 0.008 | Yes |
| Cd38 | P56528 | 0.334 ± 0.001 | Potentially | Gpc4 | P51655 | 0.523 ± 0.012 | Yes | ||||
| CD antigens | Entpd8 | Q8K0L2 | 0.386 ± 0.004 | Yes | Epb41l2 | O70318 | 0.568 ± 0.003 | Yes | |||
| Lamp1 | P11438 | 0.356 ± 0.001 | Yes | Entpd1 | P55772 | 0.400 ± 0.011 | Potentially | Dysf | Q9ESD7 | 0.569 ± 0.001 | Yes |
| Eng | Q63961 | 0.378 ± 0.002 | Yes | Anpep | P97449 | 0.406 ± 0.001 | Yes | Myof | Q69ZN7 | 0.584 ± 0.006 | Yes |
| Cd44 | P15379 | 0.405 ± 0.017 | Yes | Enpp3 | Q6DYE8 | 0.411 ± 0.009 | Yes | Sidt2 | Q8CIF6 | 0.586 ± 0.004 | Yes |
| Lamp2 | P17047 | 0.410 ± 0.001 | Yes | Nt5e | Q61503 | 0.414 ± 0.002 | Yes | Raet1d | Q9JI58 | 0.587 ± 0.014 | Yes |
| Pecam1 | Q08481 | 0.433 ± 0.005 | Yes | Cemip2 | Q5FWI3 | 0.438 ± 0.002 | Yes | Clec4g | Q8BNX1 | 0.590 ± 0.002 | Yes |
| Cd1d1 | P11609 | 0.511 ± 0.005 | Yes | Got2 | P05202 | 0.444 ± 0.002 | Yes | Irgm1 | Q60766 | 0.599 ± 0.002 | Yes |
| Bst2 | Q8R2Q8 | 0.563 ± 0.002 | Yes | Abhd17b | Q7M759 | 0.450 ± 0.020 | Yes | Stx8 | O88983 | 0.604 ± 0.003 | Potentially |
| Cd36 | Q08857 | 0.626 ± 0.006 | Yes | Alpl | P09242 | 0.454 ± 0.004 | Yes | Fam234b | Q8BYI8 | 0.640 ± 0.018 | Potentially |
| Cd47 | Q61735 | 0.686 ± 0.003 | Yes | Nos3 | P70313 | 0.475 ± 0.007 | Yes | Atraid | Q6PGD0 | 0.657 ± 0.034 | Yes |
| Cd59a | O55186 | 0.709 ± 0.013 | Yes | Enpp4 | Q8BTJ4 | 0.490 ± 0.005 | Yes | Pttg1ip | Q8R143 | 0.657 ± 0.009 | Yes |
| Cd68 | P31996 | 0.760 ± 0.025 | Yes | P4hb | P09103 | 0.510 ± 0.002 | Yes | Gdi2 | Q61598 | 0.661 ± 0.010 | Potentially |
| Plpp1 | Q61469 | 0.514 ± 0.013 | Yes | Hfe | P70387 | 0.678 ± 0.014 | Yes | ||||
| Integrins | Pip4p1 | Q3TWL2 | 0.519 ± 0.004 | Yes | Napa | Q9DB05 | 0.697 ± 0.003 | Yes | |||
| Itga9 | B8JK39 | 0.399 ± 0.014 | Potentially | Pdia6 | Q922R8 | 0.533 ± 0.001 | Yes | Clec2d | Q91V08 | 0.704 ± 0.008 | Yes |
| Itga1 | Q3V3R4 | 0.622 ± 0.002 | Potentially | Ece1 | Q4PZA2 | 0.536 ± 0.002 | Yes | Tmed1 | Q3V009 | 0.717 ± 0.033 | Yes |
| Itgal | P24063 | 0.648 ± 0.025 | Yes | Adam23 | Q9R1V7 | 0.552 ± 0.039 | Yes | Mal2 | Q8BI08 | 0.724 ± 0.002 | Yes |
| Itgav | P43406 | 0.707 ± 0.003 | Yes | Naalad2 | Q9CZR2 | 0.557 ± 0.008 | Yes | Kct2 | Q8K201 | 0.730 ± 0.011 | Potentially |
| Enpep | P16406 | 0.561 ± 0.001 | Yes | Hsp90aa1 | P07901 | 0.746 ± 0.008 | Yes | ||||
| Adhesion molecules | Kars1 | Q99MN1 | 0.575 ± 0.013 | Yes | Itfg1 | Q99KW9 | 0.750 ± 0.019 | Potentially | |||
| Esam | Q925F2 | 0.615 ± 0.030 | Yes | Mgll | O35678 | 0.590 ± 0.005 | Potentially | Hpcal1 | P62748 | 0.750 ± 0.017 | Potentially |
| Icam2 | P35330 | 0.701 ± 0.010 | Potentially | Ggt6 | Q6PDE7 | 0.610 ± 0.011 | Potentially | Lrrc57 | Q9D1G5 | 0.750 ± 0.018 | Potentially |
| Ctsb | P10605 | 0.629 ± 0.003 | Yes | Tmem123 | Q91Z22 | 0.756 ± 0.008 | Potentially | ||||
| Transporters | Akr1a1 | Q9JII6 | 0.632 ± 0.007 | Yes | Plin2 | P43883 | 0.760 ± 0.005 | Potentially | |||
| Slc46a3 | Q9DC26 | 0.390 ± 0.004 | Potentially | FRRS1 | Q8K385 | 0.640 ± 0.008 | Potentially | Rp2 | Q9EPK2 | 0.765 ± 0.004 | Yes |
| Slc39a4 | Q78IQ7 | 0.414 ± 0.015 | Yes | Cpd | O89001 | 0.649 ± 0.011 | Yes | ||||
| Slc44a2 | Q8BY89 | 0.449 ± 0.009 | Potentially | B4galt1 | P15535 | 0.650 ± 0.005 | Yes | Uncharacterized molecules | |||
| Abcb4 | P21440 | 0.464 ± 0.001 | Yes | Hpd | P49429 | 0.668 ± 0.001 | Potentially | Tmem59 | Q9QY73 | 0.291 ± 0.009 | Yes |
| Abcg2 | Q7TMS5 | 0.483 ± 0.001 | Yes | C1galt1 | Q9JJ06 | 0.675 ± 0.009 | Potentially | Tmem176a | Q9DCS1 | 0.363 ± 0.009 | Potentially |
| Abcb11 | Q9QY30 | 0.492 ± 0.000 | Yes | Adam10 | O35598 | 0.683 ± 0.003 | Yes | Tm9sf4 | Q8BH24 | 0.578 ± 0.002 | Potentially |
| Abcg8 | Q9DBM0 | 0.520 ± 0.007 | Yes | Eno1 | P17182 | 0.684 ± 0.003 | Yes | Tm7sf3 | Q9CRG1 | 0.662 ± 0.012 | Yes |
| Slc2a8 | Q9JIF3 | 0.524 ± 0.017 | Yes | Lnpep | Q8C129 | 0.690 ± 0.005 | Yes | ||||
| Slc46a1 | Q6PEM8 | 0.530 ± 0.005 | Yes | Park7 | Q99LX0 | 0.702 ± 0.006 | Yes | ||||
| Abcb6 | Q9DC29 | 0.532 ± 0.001 | Yes | Pi4k2b | Q8CBQ5 | 0.723 ± 0.023 | Potentially | ||||
| Abcc2 | Q8VI47 | 0.564 ± 0.001 | Yes | Pik3r4 | Q8VD65 | 0.740 ± 0.033 | Potentially | ||||
| Slc38a7 | Q8BWH0 | 0.591 ± 0.006 | Potentially | Nedd4 | P46935 | 0.741 ± 0.005 | Yes | ||||
| Abcg5 | Q99PE8 | 0.596 ± 0.011 | Yes | Cnp | P16330 | 0.754 ± 0.012 | Potentially | ||||
| Slc12a9 | Q99MR3 | 0.635 ± 0.007 | Yes | Tgm2 | P21981 | 0.765 ± 0.019 | Yes | ||||
| Slc10a5 | Q5PT54 | 0.704 ± 0.038 | Potentially | ||||||||
| Slc30a10 | Q3UVU3 | 0.750 ± 0.027 | Yes | ||||||||
| Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Localized? (Based on Uniprot) | Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Localized? (Based on Uniprot) | Protein Name | Uniprot Accession Number | B/A Ratio (Mean ± SEM) | Plasma Membrane Localized? (Based on Uniprot) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Scarb1 | Q61009 | 2.78 ± 0.02 | Yes | Slc4a1 | P04919 | 5.73 ± 0.06 | Yes | Arhgef12 | Q8R4H2 | 1.43 ± 0.05 | Potentially |
| Ptprf | A2A8L5 | 1.98 ± 0.01 | Potentially | Pdzk1 | Q9JIL4 | 5.28 ± 0.01 | Yes | ||||
| Adra1b | P97717 | 1.95 ± 0.03 | Yes | Slc9a3r1 | P70441 | 4.74 ± 0.03 | Yes | Others | |||
| Ptprg | Q05909 | 1.91 ± 0.07 | Potentially | Slc2a2 | P14246 | 3.19 ± 0.02 | Yes | Actn1 | Q7TPR4 | 5.68 ± 0.14 | Yes |
| Lsr | Q99KG5 | 1.90 ± 0.01 | Yes | Abca8a | Q8K442 | 3.09 ± 0.02 | Yes | Utrn | E9Q6R7 | 5.31 ± 0.06 | Yes |
| Ptprd | Q64487 | 1.64 ± 0.08 | Potentially | Abcc6 | Q9R1S7 | 3.02 ± 0.01 | Yes | Stard10 | Q9JMD3 | 4.83 ± 0.39 | Potentially |
| Egfr | Q01279 | 1.62 ± 0.00 | Yes | Slc26a1 | P58735 | 2.93 ± 0.01 | Yes | Tspan4 | Q9DCK3 | 4.72 ± 0.14 | Potentially |
| Insr | P15208 | 1.52 ± 0.01 | Yes | Slc1a2 | P43006 | 2.92 ± 0.04 | Yes | Pacsin3 | Q99JB8 | 4.32 ± 0.04 | Yes |
| Erbb3 | Q61526 | 1.45 ± 0.04 | Potentially | Slco1a1 | Q9QXZ6 | 2.87 ± 0.01 | Yes | Cask | O70589 | 4.27 ± 0.03 | Yes |
| Asgr1 | P34927 | 1.43 ± 0.00 | Potentially | Slc6a6 | O35316 | 2.75 ± 0.03 | Yes | Lima1 | Q9ERG0 | 4.19 ± 0.03 | Yes |
| Asgr2 | P24721 | 1.43 ± 0.00 | Potentially | Slc6a13 | P31649 | 2.75 ± 0.04 | Yes | Sntb1 | Q99L88 | 3.81 ± 0.04 | Yes |
| Slc6a11 | P31650 | 2.75 ± 0.03 | Potentially | Scrib | Q80U72 | 3.65 ± 0.07 | Yes | ||||
| GPCR related molecule | Slc10a1 | O08705 | 2.70 ± 0.01 | Potentially | C2cd2 | E9Q3C1 | 3.57 ± 0.08 | Potentially | |||
| Gna12 | P27600 | 1.66 ± 0.05 | Yes | Slc16a1 | P53986 | 2.69 ± 0.01 | Yes | Dlg1 | Q811D0 | 3.37 ± 0.04 | Yes |
| Abca1 | P41233 | 2.66 ± 0.01 | Yes | Dmd | P11531 | 3.07 ± 0.03 | Yes | ||||
| CD antigens | Slc16a7 | O70451 | 2.63 ± 0.09 | Yes | Farp1 | F8VPU2 | 3.00 ± 0.03 | Yes | |||
| Bsg | P18572 | 2.50 ± 0.01 | Yes | Slc2a9 | Q3T9X0 | 2.50 ± 0.01 | Yes | Ttyh2 | Q3TH73 | 2.76 ± 0.02 | Yes |
| Cd82 | P40237 | 2.37 ± 0.01 | Yes | Slc4a4 | O88343 | 2.42 ± 0.03 | Yes | Fam126b | Q8C729 | 2.70 ± 0.05 | Yes |
| Abca8b | Q8K440 | 2.32 ± 0.01 | Yes | Epb41l5 | Q8BGS1 | 2.61 ± 0.06 | Yes | ||||
| Adhesion molecules | Atp11c | Q9QZW0 | 2.22 ± 0.00 | Yes | Tspan9 | Q8BJU2 | 2.59 ± 0.09 | Potentially | |||
| Ctnnd1 | P30999 | 2.21 ± 0.00 | Yes | Slco1b2 | Q9JJL3 | 2.21 ± 0.01 | Yes | Twf1 | Q91YR1 | 2.58 ± 0.04 | Potentially |
| Ctnnb1 | Q02248 | 2.20 ± 0.01 | Yes | Slc22a1 | O08966 | 2.20 ± 0.04 | Yes | Ttc7a | Q8BGB2 | 2.55 ± 0.04 | Yes |
| Ctnna1 | P26231 | 2.01 ± 0.01 | Yes | Slc29a1 | Q9JIM1 | 2.11 ± 0.01 | Yes | Coro1c | Q9WUM4 | 2.44 ± 0.02 | Yes |
| Cdh2 | P15116 | 1.93 ± 0.02 | Yes | Slc30a1 | Q60738 | 2.04 ± 0.02 | Yes | Sptan1 | P16546 | 2.41 ± 0.00 | Yes |
| Cldnd1 | Q9CQX5 | 1.75 ± 0.06 | Potentially | Slc23a1 | Q9Z2J0 | 2.03 ± 0.05 | Yes | Pals2 | Q9JLB0 | 2.36 ± 0.01 | Potentially |
| Cadm1 | Q8R5M8 | 1.74 ± 0.05 | Yes | Slc39a14 | Q75N73 | 2.00 ± 0.02 | Yes | Serinc5 | Q8BHJ6 | 2.35 ± 0.11 | Yes |
| Cldn3 | Q9Z0G9 | 1.64 ± 0.02 | Yes | Slc12a7 | Q9WVL3 | 1.95 ± 0.02 | Yes | Nckap1 | P28660 | 2.29 ± 0.04 | Yes |
| Slco2b1 | Q8BXB6 | 1.94 ± 0.01 | Yes | Eps15l1 | Q60902 | 2.17 ± 0.05 | Yes | ||||
| Scaffold proteins | Slc38a3 | Q9DCP2 | 1.92 ± 0.01 | Yes | Wasf2 | Q8BH43 | 2.13 ± 0.02 | Yes | |||
| Rdx | P26043 | 2.72 ± 0.02 | Yes | Slc22a5 | Q9Z0E8 | 1.86 ± 0.06 | Yes | Phb | P67778 | 2.11 ± 0.01 | Yes |
| Ap2b1 | Q9DBG3 | 2.05 ± 0.01 | Yes | Slc22a23 | Q3UHH2 | 1.82 ± 0.05 | Potentially | Tmem30a | Q8VEK0 | 2.06 ± 0.01 | Yes |
| Ezr | P26040 | 1.98 ± 0.04 | Yes | Slc44a1 | Q6X893 | 1.82 ± 0.02 | Yes | Fam234a | Q8C0Z1 | 1.86 ± 0.01 | Potentially |
| Picalm | Q7M6Y3 | 1.94 ± 0.02 | Yes | Abcc3 | B2RX12 | 1.71 ± 0.00 | Yes | Numb | Q9QZS3 | 1.82 ± 0.01 | Yes |
| Dnm2 | P39054 | 1.74 ± 0.02 | Potentially | Slc2a1 | P17809 | 1.59 ± 0.05 | Yes | Vapa | Q9WV55 | 1.77 ± 0.03 | Yes |
| Ap2m1 | P84091 | 1.68 ± 0.01 | Yes | Stx4 | P70452 | 1.76 ± 0.01 | Yes | ||||
| Ap2a2 | P17427 | 1.66 ± 0.00 | Yes | Enzymes | Efr3a | Q8BG67 | 1.74 ± 0.01 | Yes | |||
| Ap2s1 | P62743 | 1.65 ± 0.02 | Yes | Cdc42bpb | Q7TT50 | 3.54 ± 0.01 | Yes | Sema4g | Q9WUH7 | 1.72 ± 0.07 | Yes |
| Clint1 | Q99KN9 | 1.62 ± 0.01 | Potentially | Zdhhc5 | Q8VDZ4 | 2.59 ± 0.05 | Yes | Eppk1 | Q8R0W0 | 1.71 ± 0.03 | Yes |
| Ap2a1 | P17426 | 1.60 ± 0.01 | Yes | Tgm1 | Q9JLF6 | 2.43 ± 0.01 | Potentially | Vcl | Q64727 | 1.68 ± 0.01 | Yes |
| Adcy9 | P51830 | 2.16 ± 0.11 | Yes | Plxnb2 | B2RXS4 | 1.62 ± 0.00 | Yes | ||||
| Channels | Pi4ka | E9Q3L2 | 2.07 ± 0.01 | Yes | Lrrn4 | P59383 | 1.61 ± 0.09 | Potentially | |||
| Aqp9 | Q9JJJ3 | 3.72 ± 0.02 | Yes | Atp5f1a | Q03265 | 1.86 ± 0.00 | Yes | Stxbp3 | Q60770 | 1.61 ± 0.01 | Yes |
| Clic4 | Q9QYB1 | 2.94 ± 0.03 | Yes | Steap4 | Q923B6 | 1.76 ± 0.01 | Yes | Ndrg1 | Q62433 | 1.50 ± 0.02 | Yes |
| Ilk | O55222 | 1.75 ± 0.03 | Yes | Eps15 | P42567 | 1.43 ± 0.02 | Yes | ||||
| Pumps | Enpp1 | P06802 | 1.74 ± 0.01 | Yes | |||||||
| Fxyd1 | Q9Z239 | 2.45 ± 0.02 | Yes | Pik3c2a | Q61194 | 1.59 ± 0.07 | Yes | Uncharacterized molecules | |||
| Atp1b1 | P14094 | 2.36 ± 0.00 | Yes | Adam17 | Q9Z0F8 | 1.55 ± 0.04 | Yes | Tmem150a | Q91WN2 | 3.01 ± 0.10 | Yes |
| Atp1a1 | Q8VDN2 | 2.23 ± 0.00 | Yes | ||||||||
| Atp1b3 | P97370 | 1.69 ± 0.01 | Yes | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirano, S.; Goto, R.; Uchida, Y. SWATH-Based Comprehensive Determination of the Localization of Apical and Basolateral Membrane Proteins Using Mouse Liver as a Model Tissue. Biomedicines 2022, 10, 383. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020383
Hirano S, Goto R, Uchida Y. SWATH-Based Comprehensive Determination of the Localization of Apical and Basolateral Membrane Proteins Using Mouse Liver as a Model Tissue. Biomedicines. 2022; 10(2):383. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020383
Chicago/Turabian StyleHirano, Satoshi, Ryohei Goto, and Yasuo Uchida. 2022. "SWATH-Based Comprehensive Determination of the Localization of Apical and Basolateral Membrane Proteins Using Mouse Liver as a Model Tissue" Biomedicines 10, no. 2: 383. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10020383