
Tandem Repeat Diversity in Two Closely Related Hamster Species—The Chinese Hamster (Cricetulus griseus) and Striped Hamster (Cricetulus barabensis)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Assemblies
2.2. Tandem Repeat Search
2.3. TR Nomenclature and Estimation of Their Genome Abundance
2.4. Probe Design
2.5. Animals
2.6. Statement of Ethics
2.7. Metaphase Chromosome Spreads
2.8. FISH
2.9. Microscopy and Image Acquisition
3. Results
3.1. Tandem Repeats Common for Assemblies
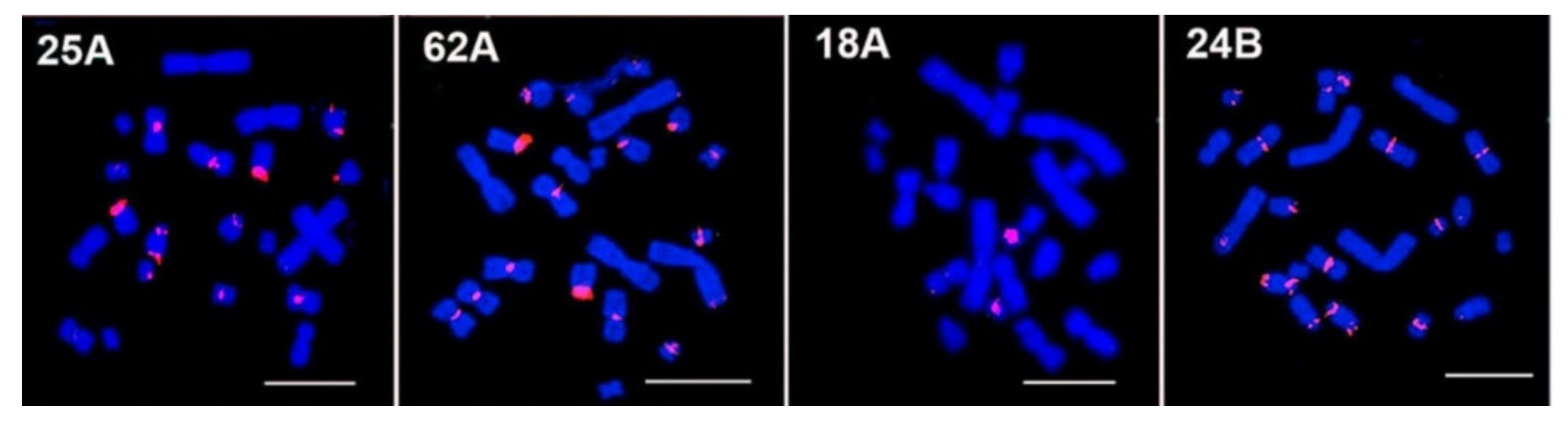
3.2. FISH Mapping on Chinese and Striped Hamsters’ Chromosomes
3.2.1. Chinese Hamster C. griseus
3.2.2. Striped Hamster C. barabensis
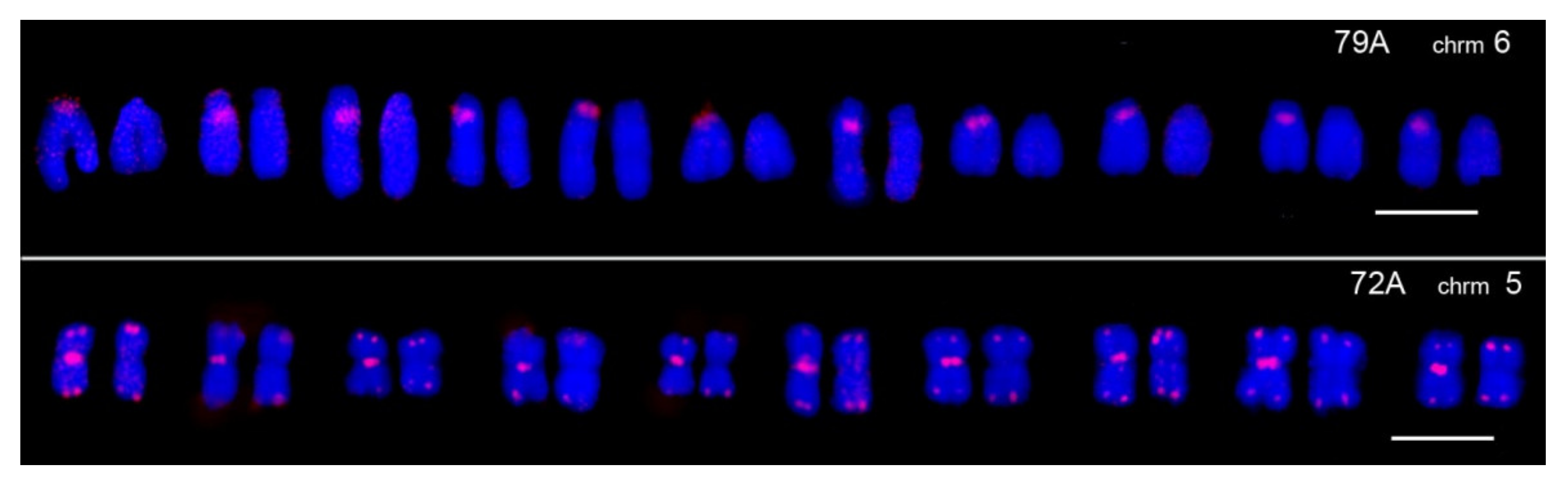
3.3. TR Heteromorphism on Homologous Chromosomes of Striped Hamsters
4. Discussion
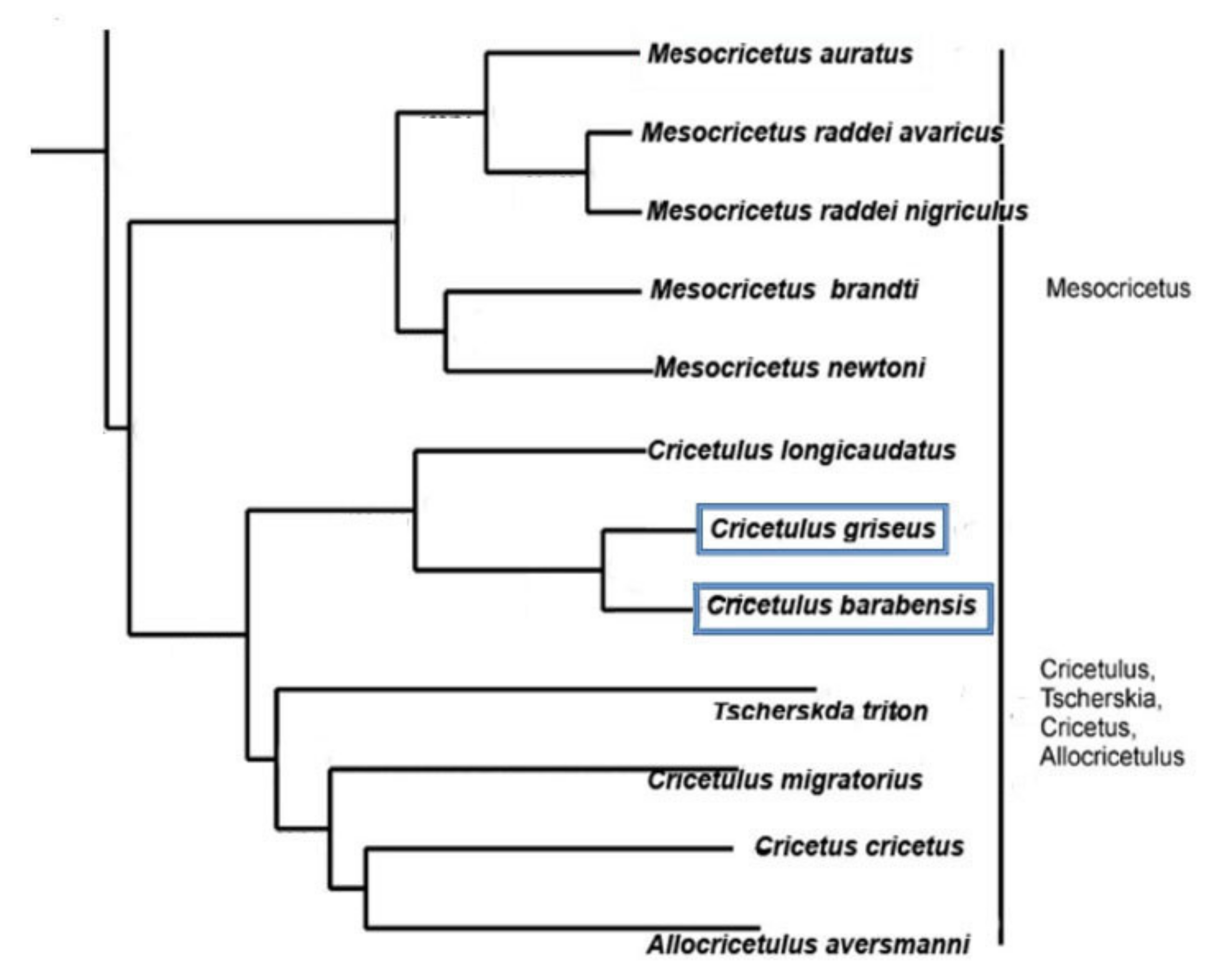
4.1. Species
4.2. Probes
4.3. Library Hypothesis
4.4. CHO (Chinese Hamster Ovary) Cell Lines
4.5. Interspecies Hybrids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miga, K.H.; Koren, S.; Rhie, A.; Vollger, M.R.; Gershman, A.; Bzikadze, A.; Brooks, S.; Howe, E.; Porubsky, D.; Logsdon, G.A.; et al. Telomere-to-Telomere Assembly of a Complete Human X Chromosome. Nature 2020, 585, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Komissarov, A.S.; Gavrilova, E.V.; Demin, S.J.; Ishov, A.M.; Podgornaya, O.I. Tandemly Repeated DNA Families in the Mouse Genome. BMC Genom. 2011, 12, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ali, M.; Zhou, Q. Establishment and Evolution of Heterochromatin. Ann. N. Y. Acad. Sci. 2020, 1476, 59–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbert, P.B.; Henikoff, S. What Makes a Centromere? Exp. Cell Res. 2020, 389, 111895. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, I.; Podgornaya, O.; Ferguson-Smith, M.A. High-Resolution Organization of Mouse Centromeric and Pericentromeric DNA. Cytogenet. Genome Res. 2006, 112, 248–255. [Google Scholar] [CrossRef]
- Alkan, C.; Cardone, M.F.; Catacchio, C.R.; Antonacci, F.; O’Brien, S.J.; Ryder, O.A.; Purgato, S.; Zoli, M.; Della Valle, G.; Eichler, E.E.; et al. Genome-Wide Characterization of Centromeric Satellites from Multiple Mammalian Genomes. Genome Res. 2011, 21, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Ruby, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative Analysis of Tandem Repeats from Hundreds of Species Reveals Unique Insights into Centromere Evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef]
- Valeri, M.P.; Dias, G.B.; do Espírito Santo, A.A.; Moreira, C.N.; Yonenaga-Yassuda, Y.; Sommer, I.B.; Kuhn, G.C.S.; Svartman, M. First Description of a Satellite DNA in Manatees’ Centromeric Regions. Front. Genet. 2021, 12, 694866. [Google Scholar] [CrossRef]
- Ostromyshenskii, D.I.; Kuznetsova, L.S.; Komissarov, A.S.; Kartavtseva, I.V.; Podgornaya, L. Tandem repeats in rodents genome and their mapping. Tsitologiia 2015, 57, 102–110. [Google Scholar] [CrossRef]
- Fátyol, K.; Cserpán, I.; Praznovszky, T.; Keresö, J.; Hadlaczky, G. Cloning and Molecular Characterization of a Novel Chromosome Specific Centromere Sequence of Chinese Hamster. Nucleic Acids Res. 1994, 22, 3728–3736. [Google Scholar] [CrossRef] [Green Version]
- Faravelli, M.; Moralli, D.; Bertoni, L.; Attolini, C.; Chernova, O.; Raimondi, E.; Giulotto, E. Two Extended Arrays of a Satellite DNA Sequence at the Centromere and at the Short-Arm Telomere of Chinese Hamster Chromosome 5. Cytogenet. Cell Genet. 1998, 83, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, J.R.; Moore, M.M.; Hozier, J.C. High Resolution G-Banded Chromosomes of the Mouse. Chromosoma 1987, 95, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pathak, S.; Hsu, T.C. High Resolution G-Banding Patterns of Syrian Hamster Chromosomes. Cytogenet. Cell Genet. 1982, 33, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S.A.; Perelman, P.L.; Serdukova, N.A.; Trifonov, V.A.; Biltueva, L.S.; Wang, J.; Li, T.; Nie, W.; O’Brien, P.C.M.; Volobouev, V.T.; et al. Reciprocal Chromosome Painting between Three Laboratory Rodent Species. Mamm. Genome 2006, 17, 1183–1192. [Google Scholar] [CrossRef]
- Miheev, D.Y.; Podgornaya, O.I.; Ostromyshenskii, D.I. Large tandem repeats of mesocricetus a uratus in silico and in situ. Tsitologiia 2015, 57, 95–101. [Google Scholar]
- Derouazi, M.; Martinet, D.; Besuchet Schmutz, N.; Flaction, R.; Wicht, M.; Bertschinger, M.; Hacker, D.L.; Beckmann, J.S.; Wurm, F.M. Genetic Characterization of CHO Production Host DG44 and Derivative Recombinant Cell Lines. Biochem. Biophys. Res. Commun. 2006, 340, 1069–1077. [Google Scholar] [CrossRef]
- Wurm, F.M. CHO Quasispecies—Implications for Manufacturing Processes. Processes 2013, 1, 296–311. [Google Scholar] [CrossRef]
- Stefanova, V.N.; Yartseva, N.M.; Petrov, A.V. Comparative cytogenetic analysis of monolayer and suspension Chinese Hamster Ovary Cell Lines CHO(dhfr-). Tsitologiia 2015, 57, 491–498. [Google Scholar]
- Turilova, V.; Goryachaya, T.; Yakovleva, T. Chinese Hamster Ovary Cell Line DXB-11: Chromosomal Instability and Karyotype Heterogeneity. Mol. Cytogenet. 2021, 14, 11. [Google Scholar] [CrossRef]
- Vakurin, A.; Kartavtseva, I.; Korablev, V.; Pavlenko, M. Some Features of Cytogenetic Differentiation of Striped Hamsters, Cricetulus barabensis and Cricetulus pseudogriseus (Rodentia, Cricetidae). Biol. Bull. 2014, 41, 822–831. [Google Scholar] [CrossRef]
- Poplavskaya, N.; Bannikova, A.; Neumann, K.; Pavlenko, M.; Kartavtseva, I.; Bazhenov, Y.; Bogomolov, P.; Abramov, A.; Surov, A.; Lebedev, V. Phylogeographic Structure in the Chromosomally Polymorphic Rodent Cricetulus barabensis Sensu Lato (Mammalia, Cricetidae). J. Zool. Syst. Evol. Res. 2018, 57, 679–694. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Volobouev, V.T.; Perelman, P.L.; Lebedev, V.S.; Serdukova, N.A.; Trifonov, V.A.; Biltueva, L.S.; Nie, W.; O’Brien, P.C.M.; Bulatova, N.S.; et al. Karyotype Evolution and Phylogenetic Relationships of Hamsters (Cricetidae, Muroidea, Rodentia) Inferred from Chromosomal Painting and Banding Comparison. Chromosome Res. 2007, 15, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Radzhabli, S.I.; Krjukova, E.P. Comparative analysis of the differential staining of chromosomes in 2 species of hamster: C. barabensis and C. griseus. Tsitologiia 1973, 15, 1527–1531. [Google Scholar] [PubMed]
- Kral, B.; Radjabli, S.L.; Grafodatsky, A.S.; Orlov, V.N. Comparison of Karyotypes, G-Bands and NORs in Three Cricetulus spp. (Rodentia, Cricetidae). Folia Zool 1984, 33, 85–96. [Google Scholar]
- Poplavskaya, N.S.; Lebedev, V.S.; Bannikova, A.A.; Belokon, M.M.; Belokon, Y.S.; Pavlenko, M.V.; Korablev, V.P.; Kartavtseva, I.V.; Bazhenov, Y.A.; Surov, A.V. Microsatellite loci variation and investigation of gene flow between two karyoforms of Cricetulus barabensis sensu lato (Rodentia, Cricetidae). Genetika 2017, 53, 63–78. [Google Scholar] [CrossRef]
- Paço, A.; Adega, F.; Meštrović, N.; Plohl, M.; Chaves, R. Evolutionary Story of a Satellite DNA from Phodopus sungorus (Rodentia, Cricetidae). Genome Biol. Evol. 2014, 6, 2944–2955. [Google Scholar] [CrossRef]
- Neumann, K.; Michaux, J.; Lebedev, V.; Yigit, N.; Colak, E.; Ivanova, N.; Poltoraus, A.; Surov, A.; Markov, G.; Maak, S.; et al. Molecular Phylogeny of the Cricetinae Subfamily Based on the Mitochondrial Cytochrome b and 12S RRNA Genes and the Nuclear VWF Gene. Mol. Phylogenet. Evol. 2006, 39, 135–148. [Google Scholar] [CrossRef]
- Beridze, T. Satellite DNA; Springer: Berlin/Heidelberg, Germany, 1986; ISBN 978-3-642-70773-5. [Google Scholar]
- Xu, X.; Nagarajan, H.; Lewis, N.E.; Pan, S.; Cai, Z.; Liu, X.; Chen, W.; Xie, M.; Wang, W.; Hammond, S.; et al. The Genomic Sequence of the Chinese Hamster Ovary (CHO)-K1 Cell Line. Nat. Biotechnol. 2011, 29, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Brinkrolf, K.; Rupp, O.; Laux, H.; Kollin, F.; Ernst, W.; Linke, B.; Kofler, R.; Romand, S.; Hesse, F.; Budach, W.E.; et al. Chinese Hamster Genome Sequenced from Sorted Chromosomes. Nat. Biotechnol. 2013, 31, 694–695. [Google Scholar] [CrossRef]
- Lewis, N.E.; Liu, X.; Li, Y.; Nagarajan, H.; Yerganian, G.; O’Brien, E.; Bordbar, A.; Roth, A.M.; Rosenbloom, J.; Bian, C.; et al. Genomic Landscapes of Chinese Hamster Ovary Cell Lines as Revealed by the Cricetulus griseus Draft Genome. Nat. Biotechnol. 2013, 31, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem Repeats Finder: A Program to Analyze DNA Sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, C.E.; Hamerton, J.L. A Colchicine, Hypotonic Citrate, Squash Sequence for Mammalian Chromosomes. Stain Technol. 1956, 31, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Kartavtseva, I.; Sheremetyeva, I.; Pavlenko, M. Intraspecies Multiple Chromosomal Variations Including Rare Tandem Fusion in the Russian Far Eastern Endemic Evoron Vole Alexandromys evoronensis (Rodentia, Arvicolinae). Comp. Cytogenet. 2021, 15, 393–411. [Google Scholar] [CrossRef] [PubMed]
- Tagarro, I.; Wiegant, J.; Raap, A.K.; González-Aguilera, J.J.; Fernández-Peralta, A.M. Assignment of Human Satellite 1 DNA as Revealed by Fluorescent in Situ Hybridization with Oligonucleotides. Hum. Genet. 1994, 93, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.; Mohandas, T. Proposed Banding Nomenclature for the Chinese Hamster Chromosomes (Cricetulus griseus). Cytogenet. Cell Genet. 1976, 16, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Gamperl, R.; Vistorin, G.; Rosenkranz, W. A Comparative Analysis of the Karyotypes of Cricetus cricetus and Cricetulus griseus. Chromosoma 1976, 55, 259–265. [Google Scholar] [CrossRef]
- Pertile, M.D.; Graham, A.N.; Choo, K.H.A.; Kalitsis, P. Rapid Evolution of Mouse Y Centromere Repeat DNA Belies Recent Sequence Stability. Genome Res. 2009, 19, 2202–2213. [Google Scholar] [CrossRef] [Green Version]
- Lemskaya, N.A.; Kartavtseva, I.V.; Rubtsova, N.V.; Golenishchev, F.N.; Sheremetyeva, I.N.; Graphodatsky, A.S. Chromosome Polymorphism in Microtus (Alexandromys) mujanensis (Arvicolinae, Rodentia). Cytogenet. Genome Res. 2015, 146, 238–242. [Google Scholar] [CrossRef]
- Lebedev, V.; Lissovsky, A. The Geographical Variability of Cranial Characteristics and Systematics of Striped-Back Hamsters (Cricetulus barabensis, Rodentia, Cricetidae). Zool. Zhurnal 2008, 87, 361–374. [Google Scholar]
- Pavlinov, I.; Lissovsky, A. The Mammals of Russia: A Taxonomic and Geographic Reference; KMK Scientific Press Ltd.: Moscow, Russia, 2012; ISBN 978-5-87317-844-5. [Google Scholar]
- Malygin, V.M.; Startsev, N.V.; Zima, Y. Karyotypes and Distribution of Dwarf Hamsters from the “Barabensis” Group (Rodentia, Criceti- Dae). Mosc. Univ. Biol. Sci. Bull. 1992, 2, 32–39. [Google Scholar]
- Gromov, I.M.; Erbaeva, M.A. The mammals of Russia and adjacent territories. Lagomorphs and Rodents. St. Peterburg Russ. Zool. Inst. RAN 1995, 522. [Google Scholar]
- Bradley, R.; Baker, R. A Test of the Genetic Species Concept: Cytochrome-b Sequences and Mammals. J. Mammal. 2001, 82, 960–973. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Perelman, P.L.; Trifonov, V.A.; Graphodatsky, A.S. Chromosomal Evolution in Rodentia. Heredity 2012, 108, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavagna, P.; Stone, G.; Stanyon, R. Black Rat (Rattus rattus) Genomic Variability Characterized by Chromosome Painting. Mamm. Genome 2002, 13, 157–163. [Google Scholar] [CrossRef]
- Yang, F.; O’Brien, P.C.; Ferguson-Smith, M.A. Comparative Chromosome Map of the Laboratory Mouse and Chinese Hamster Defined by Reciprocal Chromosome Painting. Chromosome Res. 2000, 8, 219–227. [Google Scholar] [CrossRef]
- Li, T.; Wang, J.; Su, W.; Yang, F. Chromosomal Mechanisms Underlying the Karyotype Evolution of the Oriental Voles (Muridae, Eothenomys). Cytogenet. Genome Res. 2006, 114, 50–55. [Google Scholar] [CrossRef]
- Telenius, H.; Pelmear, A.H.; Tunnacliffe, A.; Carter, N.P.; Behmel, A.; Ferguson-Smith, M.A.; Nordenskjöld, M.; Pfragner, R.; Ponder, B.A. Cytogenetic Analysis by Chromosome Painting Using DOP-PCR Amplified Flow-Sorted Chromosomes. Genes. Chromosomes Cancer 1992, 4, 257–263. [Google Scholar] [CrossRef]
- Telenius, H.; Carter, N.P.; Bebb, C.E.; Nordenskjöld, M.; Ponder, B.A.; Tunnacliffe, A. Degenerate Oligonucleotide-Primed PCR: General Amplification of Target DNA by a Single Degenerate Primer. Genomics 1992, 13, 718–725. [Google Scholar] [CrossRef]
- Kuznetsova, I.S.; Ostromyshenskii, D.I.; Komissarov, A.S.; Prusov, A.N.; Waisertreiger, I.S.; Gorbunova, A.V.; Trifonov, V.A.; Ferguson-Smith, M.A.; Podgornaya, O.I. LINE-Related Component of Mouse Heterochromatin and Complex Chromocenters’ Composition. Chromosome Res. 2016, 24, 309–323. [Google Scholar] [CrossRef]
- Ostromyshenskiĭ, D.I.; Kuznetsova, I.S.; Golinishchev, F.N.; Malikov, V.G.; Podgornaia, O.I. Satellite DNA as a phylogenetic marker: Case study of three genera of the Murinae subfamily. Tsitologiia 2011, 53, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Meštrović, N.; Mravinac, B. Satellite DNA Evolution. Genome Dyn. 2012, 7, 126–152. [Google Scholar] [CrossRef] [PubMed]
- Kipling, D.; Mitchell, A.R.; Masumoto, H.; Wilson, H.E.; Nicol, L.; Cooke, H.J. CENP-B Binds a Novel Centromeric Sequence in the Asian Mouse Mus Caroli. Mol. Cell. Biol. 1995, 15, 4009–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossant, J.; Croy, B.A.; Clark, D.A.; Chapman, V.M. Interspecific Hybrids and Chimeras in Mice. J. Exp. Zool. 1983, 228, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Spies, J.; Polasek-Sedlackova, H.; Lukas, J.; Somyajit, K. Homologous Recombination as a Fundamental Genome Surveillance Mechanism during DNA Replication. Genes 2021, 12, 1960. [Google Scholar] [CrossRef]
- Kaas, C.S.; Kristensen, C.; Betenbaugh, M.J.; Andersen, M.R. Sequencing the CHO DXB11 Genome Reveals Regional Variations in Genomic Stability and Haploidy. BMC Genom. 2015, 16, 160. [Google Scholar] [CrossRef] [Green Version]
- Schwarzacher-Robinson, T.; Kraemer, P.M.; Cram, L.S. Spontaneous in Vitro Neoplastic Evolution of Cultured Chinese Hamster Cells. Nucleolus Organizing Region Activity. Cancer Genet. Cytogenet. 1988, 35, 119–128. [Google Scholar] [CrossRef]
- Poplavskaia, N.S.; Lebedev, V.S.; Bannikova, A.A.; Meshcherskiĭ, I.G.; Surov, A.V. Karyotypes divergence in superspecies complex Cricetulus barabensis sensu lato and their interrelationships in natural contact zones. Zh. Obshch. Biol. 2012, 73, 183–197. [Google Scholar]
- Poplavskaya, N.; Lebedev, V.; Bannikova, A.; Malygin, V.; Surov, A. Interpopulation Karyotype Variability in Hamster of the “Barabensis” Group (Cricetidae, Rodentia) from Central Mongolia and Possible Reasons of Its Origin. Dokl. Biol. Sci. 2011, 439, 225. [Google Scholar] [CrossRef]
- Matveevsky, S.N.; Malygin, V.M.; Lebedev, V.S.; Poplavskaya, N.S.; Surov, A.V.; Kolomiets, O.L. Sporadic Disorders in the Meiotic Prophase I in Cricetulus barabensis Hybrids (Cricetidae, Rodentia) Do Not Lead to Reproductive Isolation between Karyomorphs. Caryologia 2014, 67, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Graphodatsky, A.S.; Sablina, O.V.; Meyer, M.N.; Malikov, V.G.; Isakova, E.A.; Trifonov, V.A.; Polyakov, A.V.; Lushnikova, T.P.; Vorobieva, N.V.; Serdyukova, N.A.; et al. Comparative Cytogenetics of Hamsters of the Genus Calomyscus. Cytogenet. Cell Genet. 2000, 88, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S.A.; Malikov, V.G.; Mahmoudi, A.; Golenishchev, F.N.; Lemskaya, N.A.; Pereira, J.C.; Trifonov, V.A.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Aliabadian, M.; et al. New Data on Comparative Cytogenetics of the Mouse-Like Hamsters (Calomyscus Thomas, 1905) from Iran and Turkmenistan. Genes 2021, 12, 964. [Google Scholar] [CrossRef] [PubMed]
- Mazurok, N.A.; Isaenko, A.A.; Nesterova, T.B.; Zakian, S.M. High-Resolution G-Banding of Chromosones in the Common Vole Microtus arvalis (Rodentia, Arvicolidae). Hereditas 1996, 124, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Mazurok, N.A.; Rubtsova, N.V.; Isaenko, A.A.; Pavlova, M.E.; Slobodyanyuk, S.Y.; Nesterova, T.B.; Zakian, S.M. Comparative Chromosome and Mitochondrial DNA Analyses and Phylogenetic Relationships within Common Voles (Microtus, Arvicolidae). Chromosome Res. 2001, 9, 107–120. [Google Scholar] [CrossRef]
- Basheva, E.; Torgasheva, A.; Golenishchev, F.; Frisman, L.; Borodin, P. Chromosome Synapsis and Recombination in the Hybrids between Chromosome Races of the Common Vole Microtus aravalis: “arvalis” and “obscurus”. Dokl. Biol. Sci. 2014, 456, 206–208. [Google Scholar] [CrossRef]
- Bikchurina, T.I.; Golenishchev, F.N.; Kizilova, E.A.; Mahmoudi, A.; Borodin, P.M. Reproductive Isolation Between Taxonomically Controversial Forms of the Gray Voles (Microtus, Rodentia; Arvicolinae): Cytological Mechanisms and Taxonomical Implications. Front. Genet. 2021, 12, 653837. [Google Scholar] [CrossRef]
- Bikchurina, T.I.; Tishakova, K.V.; Kizilova, E.A.; Romanenko, S.A.; Serdyukova, N.A.; Torgasheva, A.A.; Borodin, P.M. Chromosome Synapsis and Recombination in Male-Sterile and Female-Fertile Interspecies Hybrids of the Dwarf Hamsters (Phodopus, Cricetidae). Genes 2018, 9, 227. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WGS Project | Assembly Name | Sequencing Technology | Assembly Method | Total Sequence Length | Number of Contigs | Contig N50 |
|---|---|---|---|---|---|---|
| APMK | Cgr1.0 | Illumina GA Iix | ALLPATHS-LG v. 41879 | 2,332,774,290 | 319,219 | 11,899 |
| AFTD CHO | CriGri_1.0 | Illumina GA Iix | SOAPdenovo v. 1.05 | 2,399,786,748 | 265,787 | 39,361 |
| AMDS | C_griseus_v1.0 | Illumina HiSeq | SOAPdenovo v. 2.2 | 2,360,130,144 | 218,862 | 27,129 |
| No. | TR | Sequence |
|---|---|---|
| 1 | 33A | GTGATGTCACCTGAAGGGTCT |
| 2 | 79A | CTAGTTTTCTGTATTACGTTGTATCCG |
| 3 | 25B | TGTCCTTCTCTCCCCAGTGTC |
| 4 | 72A | CCTCCTAAAGACATAACTGAAATCC |
| 5 | 77A | CCTTGCCTTGCCTAAATGAGA |
| 6 | 84A | ACTGGAGAGAAACCCTATGAATACC |
| 7 | 26A | CTAGTGCTCCTGTAAGGAAGCC |
| 8 | 25A | GAAGAACCAGCTAACACTAGGC |
| 9 | 27A | AGGCTGGGACAATGGAGA |
| 10 | 62A | CAGCACTGTGACATCAGAATAGA |
| 11 | 18A | GACAGATGAGAGCTGGGTGA |
| 12 | 24B | TGGTCAGGCCTATACAGAGAG |
| 13 | 13A | GTGCAGAGTGAGAGTGCAGAGAG |
| Assembly | TR Family | Numbers of Families |
|---|---|---|
| AFTD + AMDS + APMK | 6A, 9A, 11A, 13A, 18A, 18B, 19B, 20B, 23A, 24A, 24B, 25A, 25B, 26A, 33A, 46A, 65A, 72A, 77A, 79A, 84A, 84B, 141A, 154A, 272A, 291A, 304A, 669A | 28 |
| AFTD + AMDS | 17B, 21C, 21D, 22A, 23B, 26B, 27B, 30B, 31A, 31B, 32A, 32B, 33B, 36A, 51B, 58A, 60A, 63A, 72B, 94A, 100A, 104A, 170A, 180A, 450A, 1464A | 26 |
| AFTD + APMK | 20A, 62A, 616A | 3 |
| AMDS + APMK | 27A, 87A, 146A | 3 |
| All | 60 | |
| No. | TR Family | Maximum Array Length, bp | GC, % | Amount in Genome, % | Genome Assembly | Chromosome In Silico | Repbase Similarities |
|---|---|---|---|---|---|---|---|
| 1 | 272A | 5953 | 44 | 1.0711 | AMDS | X | B1 |
| 2 | 11A | 13,947 | 50 | 0.9855 | AFTD | 9–10 | ERV2 |
| 3 | 49A | 2075 | 32 | 0.8666 | APMK | 8 | ERV |
| 4 | 767A | 7543 | 42 | 0.66232 | AMDS | NA | ERV2 |
| 5 | 6A | 36,714 | 59 | 0.5978 | AFTD | 5, 6, 8–10 | |
| 6 | 33A | 29,248 | 46 | 0.5119 | AFTD | 5 | SAU1.5 |
| 7 | 79A | 12,802 | 34 | 0.4047 | AFTD | 5, 9–10, 6, X | |
| 8 | 25B | 14,645 | 49 | 0.1788 | AMDS | 9–10 | |
| 9 | 304A | 4935 | 36 | 0.1365 | AFTD | NA | ERV2 |
| 10 | 72A | 40,914 | 39 | 0.1284 | AFTD | 1 | |
| 11 | 77A | 3456 | 40 | 0.0761 | APMK | 5, 2, 8 | |
| 12 | 84A | 2885 | 39 | 0.0715 | APMK | all | Zn-finger |
| 13 | 26A | 28,887 | 45 | 0.0569 | AFTD | X | |
| 14 | 17A | 15,866 | 41 | 0.0340 | APMK | 6 | |
| 15 | 65A | 3391 | 39 | 0.0296 | APMK | X | Tc1 |
| 16 | 25A | 13,526 | 43 | 0.0232 | APMK | 5 | |
| 17 | 27A | 2668 | 46 | 0.0172 | APMK | 6 | |
| 18 | 62A | 8036 | 36 | 0.0076 | APMK | 2 | |
| 19 | 18A | 11,035 | 49 | 0.0074 | AFTD | 6, 2, 9–10 | |
| 20 | 24B | 5004 | 48 | 0.0004 | APMK | 7 | |
| 21 | 13A | 1769 | 48 | 0.0003 | APMK | 3 |
| C. griseus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | X/Y |
| C. barabensis | 1 | 2 | 3 | 5 | 6 | 4 | 7 | 8 | 9 | X/Y | |
| TR | C. griseus | C. barabensis |
|---|---|---|
| 33A | C: 5, 10; | C: 6 (CG5) |
| I: 2 | ||
| 79A | C: 3, 4, 5, 6, 7, 8, 9, 10, X | C: 5, 6, 7, 8, 9 (CG: 4, 5, 8, 9, 10) |
| 25B | C: 3, 4, 5, 6, 7, 9, 10 | C: 6, 9 (CG: 5, 10); |
| I: pX, pY | ||
| 72A | C: 3, 4, 5, 6, 7, 8, 10, X; | C: 5, 6, 7, 9 (CG: 4, 5, 8, 10); |
| T: 1, 2, 6, Y | T: 1–9 (CG10), X | |
| 77A | C: 1, 2, 3, 4, 5, 6, 7, 9, 10; | C: 6, 4, 7, 8, 9 (CG: 5, 6/7, 8, 9, 10); |
| I: qX, pY | I: pX, pY | |
| 84A | C: 4, 5, 6, 7, 9, 10 | C: 6, 9 (CG: 5, 10); |
| I: pX, pY | ||
| 26A | C: 3, 4, 5, 6, 7, 8, 10, X; | C: 5, 6, 7, 9 (CG: 4, 5, 8, 10); |
| T: 1, 2, 3, 4, 6, 8, 10, X, Y | T: 1, 2, 3, 4 (CG: 6/7), X, Y | |
| 25A | C: 3–10, X; T: 1, 2, 6, 7, X | No signal |
| 27A | C: 3, 4, 5, 6, 7, 8, 10, X; | C: 5, 6, 7 (CG: 4, 5, 8); |
| T: 6 | T: 1–9 (CG: −10), X | |
| 62A | C: 3–8, 10; T: 1, 2, 3, 6, X, Y; I: 2 | No signal |
| 18A | C: 4, 5, 10 | No signal |
| 24B | C: 3–8, 10, X; T: 1, 2, 3, 4, 6, 8, X, Y | No signal |
| 13A | C: 4, 5, 10; | C: 6 (CG: 5) |
| I: qX, qY |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, N.G.; Kartavtseva, I.V.; Stefanova, V.N.; Ostromyshenskii, D.I.; Podgornaya, O.I. Tandem Repeat Diversity in Two Closely Related Hamster Species—The Chinese Hamster (Cricetulus griseus) and Striped Hamster (Cricetulus barabensis). Biomedicines 2022, 10, 925. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040925
Ivanova NG, Kartavtseva IV, Stefanova VN, Ostromyshenskii DI, Podgornaya OI. Tandem Repeat Diversity in Two Closely Related Hamster Species—The Chinese Hamster (Cricetulus griseus) and Striped Hamster (Cricetulus barabensis). Biomedicines. 2022; 10(4):925. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040925
Chicago/Turabian StyleIvanova, Nadezhda G., Irina V. Kartavtseva, Vera N. Stefanova, Dmitrii I. Ostromyshenskii, and Olga I. Podgornaya. 2022. "Tandem Repeat Diversity in Two Closely Related Hamster Species—The Chinese Hamster (Cricetulus griseus) and Striped Hamster (Cricetulus barabensis)" Biomedicines 10, no. 4: 925. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040925