
Expression of Pannexin 1 in the Human Kidney during Embryonal, Early Fetal and Postnatal Development and Its Prognostic Significance in Diabetic Nephropathy


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Procurement and Processing
2.2. Immunohistochemistry Procedure
2.3. Data Acquisition and Analysis
2.4. Statistical Analysis
3. Results
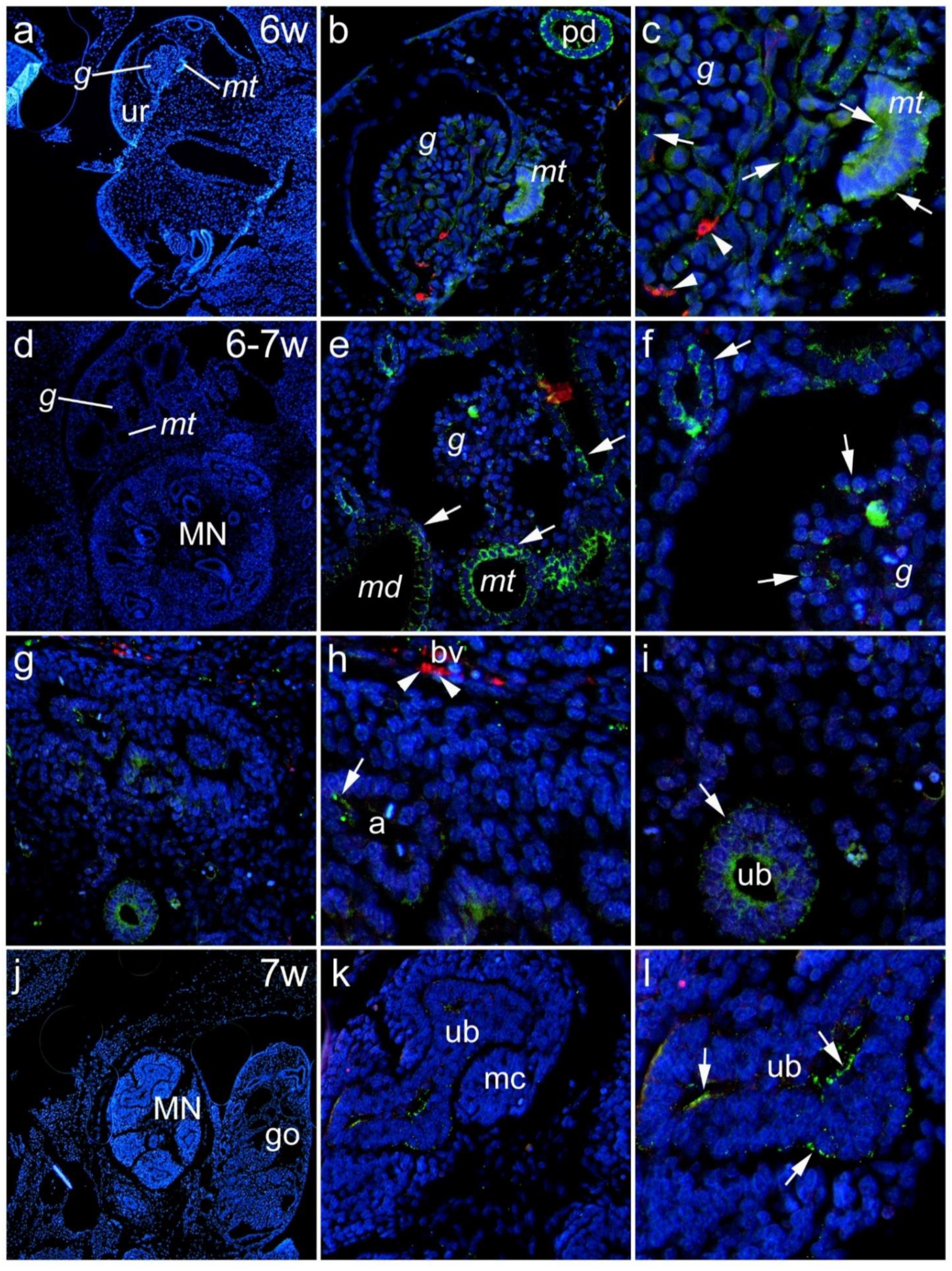
3.1. Renal Expression of Panx1 in the Human Kidney during Embryonic and Early Fetal Development
3.1.1. Sixth to Seventh Week of Development
3.1.2. Eighth to Tenth Week of Development
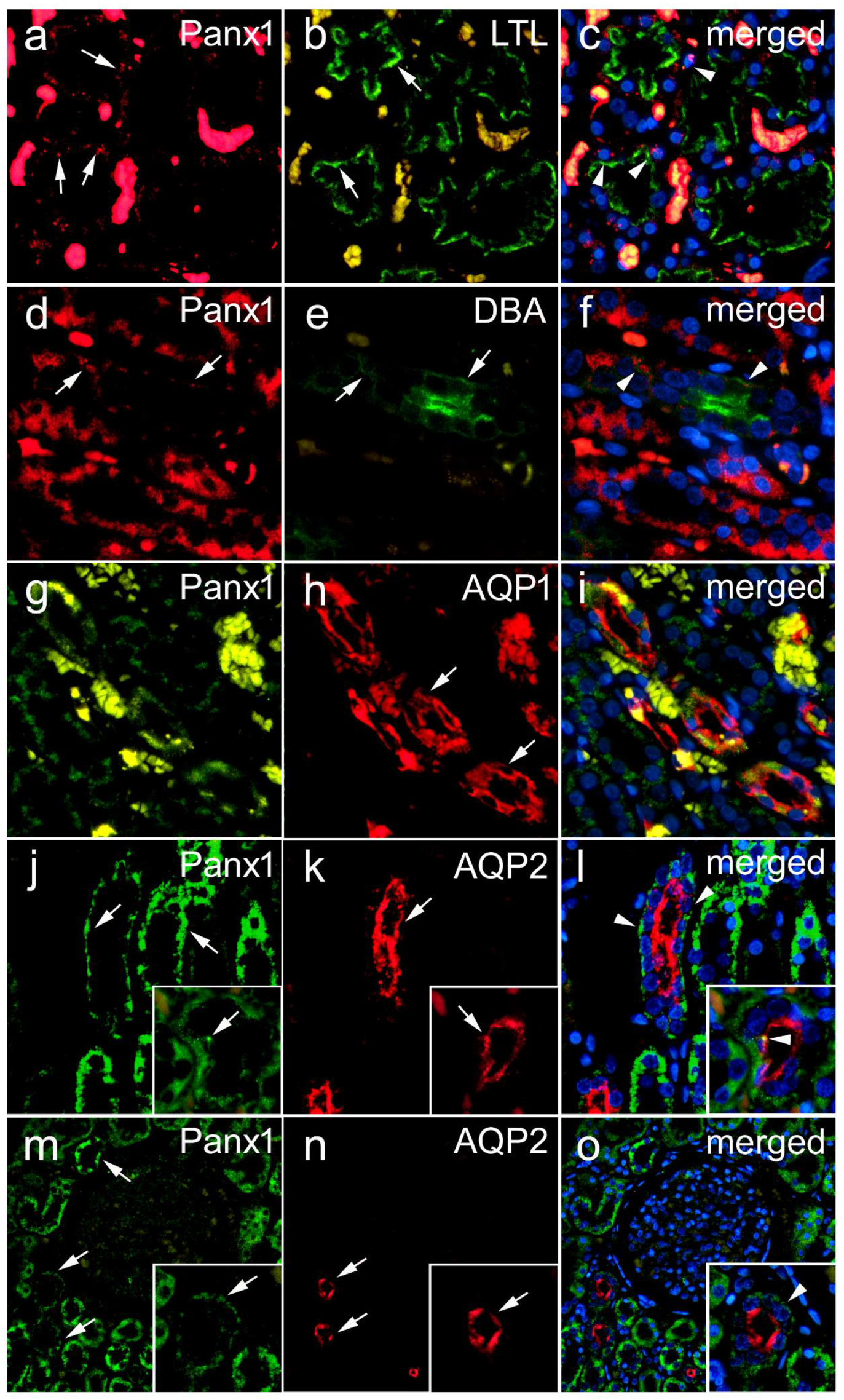
3.1.3. Lotus Tetragonolobus Lectin Binding during Embryonic and Early Fetal Development and Its Colocalization of Panx1 Expression
3.2. Renal Expression of Panx1 in Postnatal Human Kidney
3.3. Expression of Panx1 in Diabetic and Nondiabetic Renal Specimens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bao, L.; Locovei, S.; Dahl, G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbe, M.T.; Monyer, H.; Bruzzone, R. Cell-cell communication beyond connexins: The pannexin channels. Physiology 2006, 21, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Schappe, M.S.; Desai, B.N.; Bayliss, D.A. Revisiting multimodal activation and channel properties of Pannexin 1. J. Gen. Physiol. 2018, 150, 19–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo Yanguas, S.; Willebrords, J.; Johnstone, S.R.; Maes, M.; Decrock, E.; De Bock, M.; Leybaert, L.; Cogliati, B.; Vinken, M. Pannexin1 as mediator of inflammation and cell death. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 51–61. [Google Scholar] [CrossRef]
- Hanner, F.; Lam, L.; Nguyen, M.T.; Yu, A.; Peti-Peterdi, J. Intrarenal localization of the plasma membrane ATP channel pannexin1. Am. J. Physiol. Ren. Physiol. 2012, 303, F1454–F1459. [Google Scholar] [CrossRef] [Green Version]
- Lohman, A.W.; Billaud, M.; Straub, A.C.; Johnstone, S.R.; Best, A.K.; Lee, M.; Barr, K.; Penuela, S.; Laird, D.W.; Isakson, B.E. Expression of pannexin isoforms in the systemic murine arterial network. J. Vasc. Res. 2012, 49, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.; Perry, H.M.; Medina, C.B.; Huang, L.; Yao, J.; Bajwa, A.; Lorenz, U.M.; Rosin, D.L.; Ravichandran, K.S.; Isakson, B.E.; et al. Epithelial and Endothelial Pannexin1 Channels Mediate AKI. J. Am. Soc. Nephrol. JASN 2018, 29, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, Q.; Hong, J.; Ritter, J.K.; Li, P.L. Inhibition of pannexin-1 channel activity by adiponectin in podocytes: Role of acid ceramidase activation. Biochim. Et Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1246–1256. [Google Scholar] [CrossRef]
- DeLalio, L.J.; Masati, E.; Mendu, S.; Ruddiman, C.A.; Yang, Y.; Johnstone, S.R.; Milstein, J.A.; Keller, T.C.S.t.; Weaver, R.B.; Guagliardo, N.A.; et al. Pannexin 1 channels in renin-expressing cells influence renin secretion and blood pressure homeostasis. Kidney Int. 2020, 98, 630–644. [Google Scholar] [CrossRef]
- Yin, F.; Zheng, P.Q.; Zhao, L.Q.; Wang, Y.Z.; Miao, N.J.; Zhou, Z.L.; Cheng, Q.; Chen, P.P.; Xie, H.Y.; Li, J.Y.; et al. Caspase-11 promotes NLRP3 inflammasome activation via the cleavage of pannexin1 in acute kidney disease. Acta Pharmacol. Sin. 2022, 43, 86–95. [Google Scholar] [CrossRef]
- Moore, K.L.; Persaud, T.V.N.; Torchia, M.G. The Developing Human: Clinically Oriented Embryology; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Sorokin, L.; Ekblom, P. Development of tubular and glomerular cells of the kidney. Kidney Int. 1992, 41, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, M.; Nakayama, K.; Terada, Y.; Hoshi, S.; Watanabe, T. Cell cycle regulation and differentiation in the human podocyte lineage. Am. J. Pathol. 1998, 153, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Kosovic, I.; Filipovic, N.; Benzon, B.; Vukojevic, K.; Saraga, M.; Glavina Durdov, M.; Bocina, I.; Saraga-Babic, M. Spatio-temporal patterning of different connexins in developing and postnatal human kidneys and in nephrotic syndrome of the Finnish type (CNF). Sci. Rep. 2020, 10, 8756. [Google Scholar] [CrossRef] [PubMed]
- Kosovic, I.; Filipovic, N.; Benzon, B.; Bocina, I.; Glavina Durdov, M.; Vukojevic, K.; Saraga, M.; Saraga-Babic, M. Connexin Signaling in the Juxtaglomerular Apparatus (JGA) of Developing, Postnatal Healthy and Nephrotic Human Kidneys. Int. J. Mol. Sci. 2020, 21, 8349. [Google Scholar] [CrossRef]
- Price, G.W.; Potter, J.A.; Williams, B.M.; Cliff, C.L.; Wall, M.J.; Hills, C.E.; Squires, P.E. Examining Local Cell-to-Cell Signalling in the Kidney Using ATP Biosensing. Methods Mol. Biol. 2021, 2346, 135–149. [Google Scholar] [CrossRef]
- Sun, X.; Huang, K.; Haiming, X.; Lin, Z.; Yang, Y.; Zhang, M.; Liu, P.; Huang, H. Connexin 43 prevents the progression of diabetic renal tubulointerstitial fibrosis by regulating the SIRT1-HIF-1alpha signaling pathway. Clin. Sci. 2020, 134, 1573–1592. [Google Scholar] [CrossRef]
- Price, G.W.; Potter, J.A.; Williams, B.M.; Cliff, C.L.; Squires, P.E.; Hills, C.E. Connexin-mediated cell communication in the kidney: A potential therapeutic target for future intervention of diabetic kidney disease? Joan Mott Prize Lecture. Exp. Physiol. 2020, 105, 219–229. [Google Scholar] [CrossRef]
- Hills, C.; Price, G.W.; Wall, M.J.; Kaufmann, T.J.; Chi-Wai Tang, S.; Yiu, W.H.; Squires, P.E. Transforming Growth Factor Beta 1 Drives a Switch in Connexin Mediated Cell-to-Cell Communication in Tubular Cells of the Diabetic Kidney. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 45, 2369–2388. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Tao, J.; Li, G.; Zheng, D.; Tan, Y.; Li, R.; Tian, L.; Li, Z.; Cheng, H.; Xie, X. Astaxanthin ameliorates experimental diabetes-induced renal oxidative stress and fibronectin by upregulating connexin43 in glomerular mesangial cells and diabetic mice. Eur. J. Pharmacol. 2018, 840, 33–43. [Google Scholar] [CrossRef]
- Li, H.; Wang, F. The role of connexin43 in diabetic microvascular complications. Discov. Med. 2016, 22, 275–280. [Google Scholar]
- Luetic, M.; Vitlov Uljevic, M.; Masek, T.; Benzon, B.; Vukojevic, K.; Filipovic, N. PUFAs supplementation affects the renal expression of pannexin 1 and connexins in diabetic kidney of rats. Histochem. Cell Biol. 2020, 153, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Vitlov Uljevic, M.; Starcevic, K.; Masek, T.; Bocina, I.; Restovic, I.; Kevic, N.; Racetin, A.; Kretzschmar, G.; Grobe, M.; Vukojevic, K.; et al. Dietary DHA/EPA supplementation ameliorates diabetic nephropathy by protecting from distal tubular cell damage. Cell Tissue Res. 2019, 378, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Vukojevic, K.; Raguz, F.; Saraga, M.; Filipovic, N.; Bocina, I.; Kero, D.; Glavina Durdov, M.; Martinovic, V.; Saraga-Babic, M. Glomeruli from patients with nephrin mutations show increased number of ciliated and poorly differentiated podocytes. Acta Histochem. 2018, 120, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, N.; Vukojevic, K.; Bocina, I.; Saraga, M.; Durdov, M.G.; Kablar, B.; Saraga-Babic, M. Immunohistochemical and electronmicroscopic features of mesenchymal-to-epithelial transition in human developing, postnatal and nephrotic podocytes. Histochem. Cell Biol. 2017, 147, 481–495. [Google Scholar] [CrossRef] [PubMed]
- Menzies, R.I.; Tam, F.W.; Unwin, R.J.; Bailey, M.A. Purinergic signaling in kidney disease. Kidney Int. 2017, 91, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, R.L. Evolution, kidney development, and chronic kidney disease. Semin. Cell Dev. Biol. 2019, 91, 119–131. [Google Scholar] [CrossRef]
- Huang, G.; Bao, J.; Shao, X.; Zhou, W.; Wu, B.; Ni, Z.; Wang, L. Inhibiting pannexin-1 alleviates sepsis-induced acute kidney injury via decreasing NLRP3 inflammasome activation and cell apoptosis. Life Sci. 2020, 254, 117791. [Google Scholar] [CrossRef]
- Williams, J.R. The Declaration of Helsinki and public health. Bull. World Health Organ. 2008, 86, 650–652. [Google Scholar] [CrossRef]
- O’Rahilly, R.; Gardner, E. The timing and sequence of events in the development of the human nervous system during the embryonic period proper. Z. Anat. Entwickl. 1971, 134, 1–12. [Google Scholar] [CrossRef]
- Swayne, L.A.; Bennett, S.A. Connexins and pannexins in neuronal development and adult neurogenesis. BMC Cell Biol. 2016, 17 (Suppl. 1), 10. [Google Scholar] [CrossRef] [Green Version]
- Worsdorfer, P.; Wagner, N.; Ergun, S. The role of connexins during early embryonic development: Pluripotent stem cells, gene editing, and artificial embryonic tissues as tools to close the knowledge gap. Histochem. Cell Biol. 2018, 150, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Gap junctional communication in morphogenesis. Prog. Biophys. Mol. Biol. 2007, 94, 186–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carev, D.; Krnic, D.; Saraga, M.; Sapunar, D.; Saraga-Babic, M. Role of mitotic, pro-apoptotic and anti-apoptotic factors in human kidney development. Pediatr. Nephrol. 2006, 21, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Carev, D.; Saraga, M.; Saraga-Babic, M. Involvement of FGF and BMP family proteins and VEGF in early human kidney development. Histol. Histopathol. 2008, 23, 853–862. [Google Scholar] [CrossRef]
- Carev, D.; Saraga, M.; Saraga-Babic, M. Expression of intermediate filaments, EGF and TGF-alpha in early human kidney development. J. Mol. Histol. 2008, 39, 227–235. [Google Scholar] [CrossRef]
- Hillman, K.A.; Johnson, T.M.; Winyard, P.J.; Burnstock, G.; Unwin, R.J.; Woolf, A.S. P2X(7) receptors are expressed during mouse nephrogenesis and in collecting duct cysts of the cpk/cpk mouse. Exp. Nephrol. 2002, 10, 34–42. [Google Scholar] [CrossRef]
- Poudel, N.; Okusa, M.D. Pannexins in Acute Kidney Injury. Nephron 2019, 143, 158–161. [Google Scholar] [CrossRef]
- Kirby, B.S.; Sparks, M.A.; Lazarowski, E.R.; Lopez Domowicz, D.A.; Zhu, H.; McMahon, T.J. Pannexin 1 channels control the hemodynamic response to hypoxia by regulating O2-sensitive extracellular ATP in blood. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1055–H1065. [Google Scholar] [CrossRef]
- Ravindran, S.; Kuruvilla, V.; Wilbur, K.; Munusamy, S. Nephroprotective Effects of Metformin in Diabetic Nephropathy. J. Cell. Physiol. 2017, 232, 731–742. [Google Scholar] [CrossRef]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Code No. | Host | Dilution | Source | |
|---|---|---|---|---|---|
| Primary | Anti-Pannexin 1/PANX1 | ABN242 | Rabbit | 1:300 | Merck KGaA, Darmstadt, Germany |
| Anti-Nephrin (B-12) | sc-377246 | Mouse | 1:50 | Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA | |
| Anti-Aquaporin 1/AQP1 (B-11) | sc-25287 | Mouse | 1:50 | Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA | |
| Anti-Aquaporin 2/AQP2 (E-2) | sc-515770 | Mouse | 1:50 | Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA | |
| Anti-Renin [7D3-E3] | ab134783 | Mouse | 1:50 | Abcam, Cambridge, UK | |
| Anti-Smooth Muscle Actin | M0851 | Mouse | 1:300 | Dako, Glostrup, Denmark | |
| Lectins | Fluorescein labeled Dolichos biflorus agglutinin (DBA) | FL-1031 | - | 1:400 | Vector Laboratories Ltd., Peterborough, UK |
| Fluorescein labeled Lotus Tetragonolobus lectin (LTL) | FL-1321 | - | 1:400 | Vector Laboratories Ltd., Peterborough, UK | |
| Secondary | Alexa Fluor®488 AffiniPure Anti-Mouse lgG (H+L) | 715-545-150 | Donkey | 1:400 | Jackson Immuno Research Laboratories, Inc., Baltimore, PA, USA |
| Alexa Fluor®488 AffiniPure Anti-Rabbit lgG (H+L) | 711-545-152 | Donkey | 1:400 | Jackson Immuno Research Laboratories, Inc., Baltimore, PA, USA | |
| Rhodamine Red™-X (RRX) AffiniPure Anti-Mouse IgG (H+L) | 715-295-151 | Donkey | 1:400 | Jackson Immuno Research Laboratories, Inc., Baltimore, PA, USA | |
| Rhodamine Red™-X (RRX) AffiniPure Donkey Anti-Rabbit IgG (H+L) | 711-295-152 | Donkey | 1:400 | Jackson Immuno Research Laboratories, Inc., Baltimore, PA, USA |
| Panx 1 | Panx 1 | |||||
|---|---|---|---|---|---|---|
| N | N | Age | Creatinine | Glomeruli | t/i | |
| Male | Female | (Years) | (μmol/L) | % Area | % Area | |
| All | 29 | 12 | 66.7 ± 9.43 (44–89) | 103.4 ± 42.2 (50–250) | 8.53 ± 6.89 (1.94–41.33) | 2.67 ± 1.16 (0.85–1.13) |
| Nondiabetic | 16 | 4 | 64.4 ± 8.8 (44–75) | 99.6 ± 27.8 (50–164) | 7.57 ± 5.13 (2.01–34.2) | 2.5 ± 0.99 (0.85–8.13) |
| Diabetic | 13 | 8 | 68.6 ± 9.9 (49–89) | 107.3 ± 54.2 (61–250) | 9.45 ± 8.13 (1.94–41.33) | 2.82 ± 1.3 (0.9–6.54) |
| Panx 1 % Area Glomeruli | Panx 1 % Area t/i | ||
|---|---|---|---|
| All | Age | 0.151 | 0.105 |
| p = 0.003 | p = 0.044 | ||
| Creatinine | 0.162 | −0.033 | |
| p = 0.002 | p = 0.543 | ||
| Nondiabetic | Age | 0.065 | 0.089 |
| p = 0.384 | p = 0.249 | ||
| Creatinine | 0.008 | −0.071 | |
| p = 0.914 | p = 0.344 | ||
| Diabetic | Age | 0.227 | 0.213 |
| p < 0.001 | p = 0.003 | ||
| Creatinine | 0.265 | 0.004 | |
| p < 0.001 | p = 0.963 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeličić, I.; Vukojević, K.; Racetin, A.; Čarić, D.; Glavina Durdov, M.; Saraga-Babić, M.; Filipović, N. Expression of Pannexin 1 in the Human Kidney during Embryonal, Early Fetal and Postnatal Development and Its Prognostic Significance in Diabetic Nephropathy. Biomedicines 2022, 10, 944. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10050944
Jeličić I, Vukojević K, Racetin A, Čarić D, Glavina Durdov M, Saraga-Babić M, Filipović N. Expression of Pannexin 1 in the Human Kidney during Embryonal, Early Fetal and Postnatal Development and Its Prognostic Significance in Diabetic Nephropathy. Biomedicines. 2022; 10(5):944. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10050944
Chicago/Turabian StyleJeličić, Ivo, Katarina Vukojević, Anita Racetin, Davor Čarić, Merica Glavina Durdov, Mirna Saraga-Babić, and Natalija Filipović. 2022. "Expression of Pannexin 1 in the Human Kidney during Embryonal, Early Fetal and Postnatal Development and Its Prognostic Significance in Diabetic Nephropathy" Biomedicines 10, no. 5: 944. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10050944