
Targeting Tumor Cells Overexpressing the Human Epidermal Growth Factor Receptor 3 with Potent Drug Conjugates Based on Affibody Molecules

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Gene Construction
2.3. Protein Expression and Purification
2.4. Conjugation with DM1
2.5. Biochemical Characterization
2.6. Affinity Determination
2.7. Cell Lines
2.8. Determination of Cytotoxicity
2.9. Radiolabeling with [99mTc]Tc and Analysis of the Stability of the Labeled Constructs
2.10. Binding Specificity, Internalization, and Retention
2.11. Cell Binding Analysis
2.12. Animal Studies
3. Results
3.1. Drug Conjugate Design
3.2. Protein Expression, DM1 Conjugation, and Biochemical Characterization
3.3. Determination of Affinity to HER3 and Murine ErbB3
3.4. Determination of Cytotoxic Potential
3.5. Radiolabeling with [99mTc]Tc
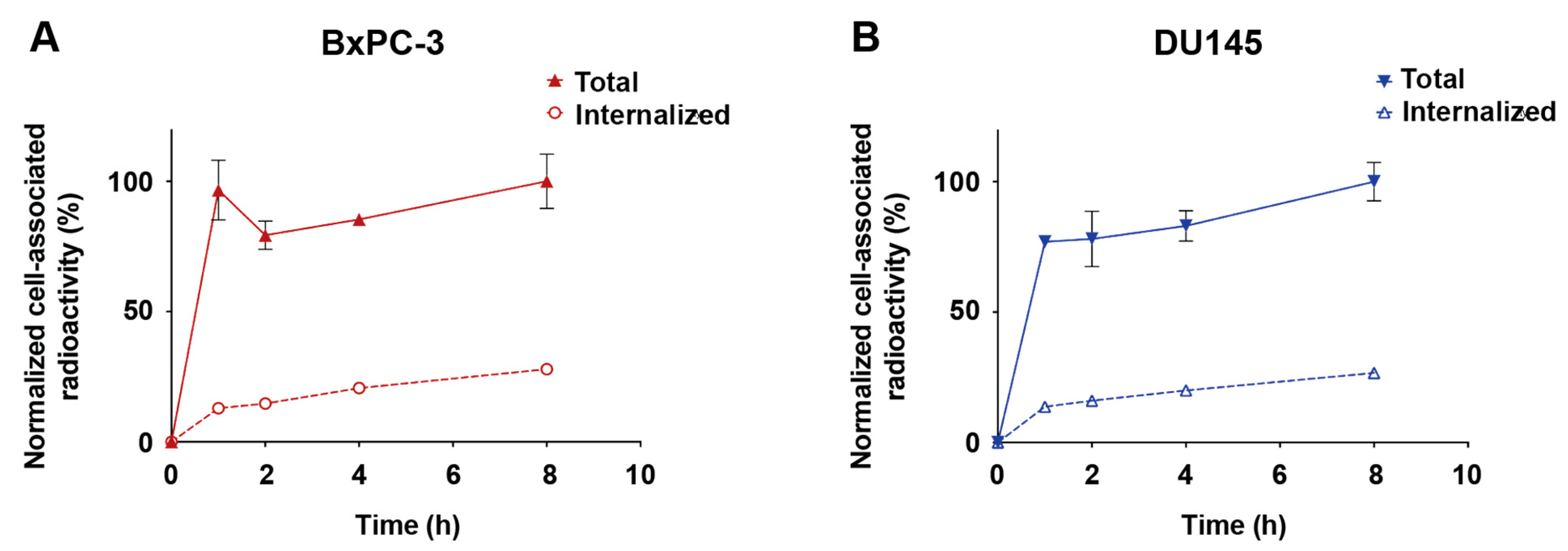
3.6. Cell-Binding Specificity and Rate of Internalization
3.7. Affinity to BxPC-3 Cells
3.8. Biodistribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zahavi, D.; Weiner, L. Monoclonal antibodies in cancer therapy. Antibodies 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Xenaki, K.T.; Oliveira, S.; van Bergen en Henegouwen, P.M.P. Antibody or antibody fragments: Implications for molecular imaging and targeted therapy of solid tumors. Front. Immunol. 2017, 8, 1287. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, M.; Skerra, A. Engineered Protein Scaffolds as Next-Generation Therapeutics. Annu. Rev. Pharm. Toxicol. 2020, 6, 391–415. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Patel, H.; Alanazi, S.; Yuan, L.; Garrett, J.T. HER3 signaling and targeted therapy in cancer. Oncol. Rev. 2018, 12, 45–62. [Google Scholar] [CrossRef] [Green Version]
- Siegfried, J.M.; Lin, Y.; Diergaarde, B.; Lin, H.M.; Dacic, S.; Pennathur, A.; Weissfeld, J.L.; Romkes, M.; Nukui, T.; Stabile, L.P. Expression of PAM50 Genes in Lung Cancer: Evidence that Interactions between Hormone Receptors and HER2/HER3 Contribute to Poor Outcome. Neoplasia 2015, 17, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Liles, J.S.; Arnoletti, J.P.; Tzeng, C.W.D.; Howard, J.H.; Kossenkov, A.V.; Kulesza, P.; Heslin, M.J.; Frolov, A. ErbB3 expression promotes tumorigenesis in pancreatic adenocarcinoma. Cancer Biol. Ther. 2010, 10, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Reschke, M.; Mihic-Probst, D.; Van Horst, E.H.; Der Knyazev, P.; Wild, P.J.; Hutterer, M.; Meyer, S.; Dummer, R.; Moch, H.; Ullrich, A. HER3 Is a determinant for poor prognosis in melanoma. Clin. Cancer Res. 2008, 14, 5188–5197. [Google Scholar] [CrossRef] [Green Version]
- Koumakpayi, I.H.; Diallo, J.S.; Le Page, C.; Lessard, L.; Gleave, M.; Bégin, L.R.; Mes-Masson, A.M.; Saad, F. Expression and nuclear localization of ErbB3 in prostate cancer. Clin. Cancer Res. 2006, 12, 2730–2737. [Google Scholar] [CrossRef] [Green Version]
- Lipton, A.; Goodman, L.; Leitzel, K.; Cook, J.; Sperinde, J.; Haddad, M.; Köstler, W.J.; Huang, W.; Weidler, J.M.; Ali, S.; et al. HER3, p95HER2, and HER2 protein expression levels define multiple subtypes of HER2-positive metastatic breast cancer. Breast Cancer Res. Treat. 2013, 141, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [Green Version]
- Laskin, J.; Liu, S.V.; Tolba, K.; Heining, C.; Schlenk, R.F.; Cheema, P.; Cadranel, J.; Jones, M.R.; Drilon, A.; Cseh, A.; et al. NRG1 fusion-driven tumors: Biology, detection, and the therapeutic role of afatinib and other ErbB-targeting agents. Ann. Oncol. 2020, 31, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Khongorzul, P.; Ling, C.J.; Khan, F.U.; Ihsan, A.U.; Zhang, J. Antibody-drug conjugates: A comprehensive review. Mol. Cancer Res. 2020, 18, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, N.; Beck, A.; Dumontet, C.; Denevault-sabourin, C. Antibody—Drug Conjugates: The Last Decade. Pharmaceuticals 2020, 13, 245. [Google Scholar] [CrossRef] [PubMed]
- Wolska-Washer, A.; Robak, T. Safety and Tolerability of Antibody-Drug Conjugates in Cancer. Drug Saf. 2019, 42, 295–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jänne, P.A.; Baik, C.; Su, W.-C.; Johnson, M.L.; Hayashi, H.; Nishio, M.; Kim, D.-W.; Koczywas, M.; Gold, K.A.; Steuer, C.E.; et al. Efficacy and Safety of Patritumab Deruxtecan (HER3-DXd) in EGFR Inhibitor–Resistant, EGFR-Mutated Non–Small Cell Lung Cancer. Cancer Discov. 2021, 12, 74–89. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Koyama, K.; Kamai, Y.; Hirotani, K.; Ogitani, Y.; Zembutsu, A.; Abe, M.; Kaneda, Y.; Maeda, N.; Shiose, Y.; et al. A novel HER3-targeting antibody–drug conjugate, U3-1402, exhibits potent therapeutic efficacy through the delivery of cytotoxic payload by efficient internalization. Clin. Cancer Res. 2019, 25, 7151–7161. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Larson, R. MCLA-128 Fights NRG1 Fusion-Positive Cancers. Cancer Discov. 2019, 9, 1636. [Google Scholar]
- Robinson, M.K.; Hodge, K.M.; Horak, E.; Sundberg, Å.L.; Russeva, M.; Shaller, C.C.; Von Mehren, M.; Shchaveleva, I.; Simmons, H.H.; Marks, J.D.; et al. Targeting ErbB2 and ErbB3 with a bispecific single-chain Fv enhances targeting selectivity and induces a therapeutic effect in vitro. Br. J. Cancer 2008, 99, 1415–1425. [Google Scholar] [CrossRef] [Green Version]
- Löfblom, J.; Feldwisch, J.; Tolmachev, V.; Carlsson, J.; Ståhl, S.; Frejd, F.Y. Affibody molecules: Engineered proteins for therapeutic, diagnostic and biotechnological applications. FEBS Lett. 2010, 584, 2670–2680. [Google Scholar] [CrossRef] [Green Version]
- Ståhl, S.; Gräslund, T.; Eriksson Karlström, A.; Frejd, F.Y.; Nygren, P.Å.; Löfblom, J. Affibody Molecules in Biotechnological and Medical Applications. Trends Biotechnol. 2017, 35, 691–712. [Google Scholar] [CrossRef]
- Kronqvist, N.; Malm, M.; Göstring, L.; Gunneriusson, E.; Nilsson, M.; Höidén Guthenberg, I.; Gedda, L.; Frejd, F.Y.; Ståhl, S.; Löfblom, J. Combining phage and staphylococcal surface display for generation of ErbB3-specific Affibody molecules. Protein Eng. Des. Sel. 2011, 24, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malm, M.; Kronqvist, N.; Lindberg, H.; Gudmundsdotter, L.; Bass, T.; Frejd, F.Y.; Höidén-Guthenberg, I.; Varasteh, Z.; Orlova, A.; Tolmachev, V.; et al. Inhibiting HER3-Mediated Tumor Cell Growth with Affibody Molecules Engineered to Low Picomolar Affinity by Position-Directed Error-Prone PCR-Like Diversification. PLoS ONE 2013, 8, e62791. [Google Scholar] [CrossRef]
- Da Pieve, C.; Allott, L.; Martins, C.D.; Vardon, A.; Ciobota, D.M.; Kramer-Marek, G.; Smith, G. Efficient [18F]AlF Radiolabeling of ZHER3:8698 Affibody Molecule for Imaging of HER3 Positive Tumors. Bioconjug. Chem. 2016, 27, 1839–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinne, S.S.; Leitao, C.D.; Abouzayed, A.; Vorobyeva, A.; Tolmachev, V.; Ståhl, S.; Löfblom, J.; Orlova, A. HER3 pet imaging:68Ga-labeled affibody molecules provide superior HER3 contrast to 89Zr-labeled antibody and antibody-fragment-based tracers. Cancers 2021, 13, 4791. [Google Scholar] [CrossRef] [PubMed]
- Orlova, A.; Malm, M.; Rosestedt, M.; Varasteh, Z.; Andersson, K.; Selvaraju, R.K.; Altai, M.; Honarvar, H.; Strand, J.; Ståhl, S.; et al. Imaging of HER3-expressing xenografts in mice using a 99mTc(CO) 3-HEHEHE-ZHER3:08699 affibody molecule. Eur. J. Nucl. Med. Mol. Imaging 2014, 41, 1450–1459. [Google Scholar] [CrossRef] [Green Version]
- Leitao, C.D.; Rinne, S.S.; Mitran, B.; Vorobyeva, A.; Andersson, K.G.; Tolmachev, V.; Ståhl, S.; Löfblom, J.; Orlova, A. Molecular design of HER3-targeting affibody molecules: Influence of chelator and presence of HEHEHE-Tag on biodistribution of 68 Ga-labeled tracers. Int. J. Mol. Sci. 2019, 20, 1080. [Google Scholar] [CrossRef] [Green Version]
- Rinne, S.S.; Leitao, C.D.; Saleh-Nihad, Z.; Mitran, B.; Tolmachev, V.; Ståhl, S.; Löfblom, J.; Orlova, A. Benefit of later-time-point PET imaging of her3 expression using optimized radiocobalt-labeled affibody molecules. Int. J. Mol. Sci. 2020, 21, 1972. [Google Scholar] [CrossRef] [Green Version]
- Rinne, S.S.; Orlova, A.; Tolmachev, V. Pet and spect imaging of the egfr family (Rtk class i) in oncology. Int. J. Mol. Sci. 2021, 22, 3663. [Google Scholar] [CrossRef]
- Rosestedt, M.; Andersson, K.G.; Mitran, B.; Tolmachev, V.; Löfblom, J.; Orlova, A.; Ståhl, S. Affibody-mediated PET imaging of HER3 expression in malignant tumours. Sci. Rep. 2015, 5, 15226. [Google Scholar] [CrossRef] [Green Version]
- Tolmachev, V.; Hofström, C.; Malmberg, J.; Ahlgren, S.; Hosseinimehr, S.J.; Sandström, M.; Abrahmsén, L.; Orlova, A.; Gräslund, T. HEHEHE-tagged affibody molecule may be purified by IMAC, is conveniently labeled with [99mTc(CO)3]+, and shows improved biodistribution with reduced hepatic radioactivity accumulation. Bioconjug. Chem. 2010, 21, 2013–2022. [Google Scholar] [CrossRef]
- Hofström, C.; Altai, M.; Honarvar, H.; Strand, J.; Malmberg, J.; Hosseinimehr, S.J.; Orlova, A.; Gräslund, T.; Tolmachev, V. HAHAHA, HEHEHE, HIHIHI, or HKHKHK: Influence of position and composition of histidine containing tags on biodistribution of [99mTc(CO)3]+-labeled affibody molecules. J. Med. Chem. 2013, 56, 4966–4974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofström, C.; Orlova, A.; Altai, M.; Wangsell, F.; Gräslund, T.; Tolmachev, V. Use of a HEHEHE purification tag instead of a hexahistidine tag improves biodistribution of affibody molecules site-specifically labeled with 99mTc, 111In, and 125I. J. Med. Chem. 2011, 54, 3817–3826. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.J.L.; Hurvitz, S.A. Recent advances in the development of anti-HER2 antibodies and antibody-drug conjugates. Ann. Transl. Med. 2014, 2, 122–136. [Google Scholar] [PubMed]
- Verma, S.; Miles, D.; Gianni, L.; Krop, I.E.; Welslau, M.; Baselga, J.; Pegram, M.; Oh, D.-Y.; Diéras, V.; Guardino, E.; et al. Trastuzumab emtansine for HER2-positive advanced breast cancer. N. Engl. J. Med. 2012, 367, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontermann, R.E. Strategies for extended serum half-life of protein therapeutics. Curr. Opin. Biotechnol. 2011, 22, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Altai, M.; Liu, H.; Ding, H.; Mitran, B.; Edqvist, P.-H.; Tolmachev, V.; Orlova, A.; Gräslund, T. Affibody-derived drug conjugates: Potent cytotoxic molecules for treatment of HER2 over-expressing tumors. J. Control. Release 2018, 288, 84–95. [Google Scholar] [CrossRef]
- Jonsson, A.; Dogan, J.; Herne, N.; Abrahmsén, L.; Nygren, P.Å. Engineering of a femtomolar affinity binding protein to human serum albumin. Protein Eng. Des. Sel. 2008, 21, 515–527. [Google Scholar] [CrossRef]
- Salmiheimo, A.; Mustonen, H.; Stenman, U.-H.; Puolakkainen, P.; Kemppainen, E.; Seppänen, H.; Haglund, C. Systemic Inflammatory Response and Elevated Tumour Markers Predict Worse Survival in Resectable Pancreatic Ductal Adenocarcinoma. PLoS ONE 2016, 11, e0163064. [Google Scholar] [CrossRef]
- Ding, H.; Altai, M.; Rinne, S.S.; Vorobyeva, A.; Tolmachev, V.; Gräslund, T.; Orlova, A. Incorporation of a Hydrophilic Spacer Reduces Hepatic Uptake of HER2-Targeting Affibody–DM1 Drug Conjugates. Cancers 2019, 11, 1168. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Ding, H.; Vorobyeva, A.; Oroujeni, M.; Orlova, A.; Tolmachev, V.; Gräslund, T. Drug conjugates based on a monovalent affibody targeting vector can efficiently eradicate HER2 positive human tumors in an experimental mouse model. Cancers 2021, 13, 85. [Google Scholar] [CrossRef]
- Yin, W.; Xu, T.; Altai, M.; Oroujeni, M.; Zhang, J.; Vorobyeva, A.; Vorontsova, O.; Vtorushin, S.V.; Tolmachev, V.; Gräslund, T.; et al. The influence of domain permutations of an albumin-binding domain-fused her2-targeting affibody-based drug conjugate on tumor cell proliferation and therapy efficacy. Pharmaceutics 2021, 13, 1974. [Google Scholar] [CrossRef] [PubMed]
- Vorobyeva, A.; Schulga, A.; Konovalova, E.; Güler, R.; Löfblom, J.; Sandström, M.; Garousi, J.; Chernov, V.; Bragina, O.; Orlova, A.; et al. Optimal composition and position of histidine-containing tags improves biodistribution of 99mTc-labeled DARPin G3. Sci. Rep. 2019, 9, 9405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitao, C.D.; Rinne, S.S.; Altai, M.; Vorontsova, O.; Dunås, F.; Jonasson, P.; Tolmachev, V.; Löfblom, J.; Ståhl, S.; Orlova, A. Evaluating the therapeutic efficacy of mono-and bivalent affibody-based fusion proteins targeting HER3 in a pancreatic cancer xenograft model. Pharmaceutics 2020, 12, 551. [Google Scholar] [CrossRef]
- Thurber, G.M.; Schmidt, M.M.; Wittrup, K.D. Factors determining antibody distribution in tumors. Trends Pharmacol. Sci. 2008, 29, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Debie, P.; Lafont, C.; Defrise, M.; Hansen, I.; van Willigen, D.M.; van Leeuwen, F.W.B.; Gijsbers, R.; D’Huyvetter, M.; Devoogdt, N.; Lahoutte, T.; et al. Size and affinity kinetics of nanobodies influence targeting and penetration of solid tumours. J. Control. Release 2020, 317, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.; Brown, A.; Mudd, G.; Huxley, P.; Rietschoten, K.; Van Pavan, S.; Chen, L.; Watcham, S.; Lahdenranta, J.; Keen, N. MMAE delivery using the Bicycle toxin conjugate BT5528. Mol. Cancer Ther. 2020, 19, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Brandl, F.; Busslinger, S.; Zangemeister-Wittke, U.; Plückthun, A. Optimizing the anti-tumor efficacy of protein-drug conjugates by engineering the molecular size and half-life. J. Control. Release 2020, 327, 186–197. [Google Scholar] [CrossRef]
- Schmidt, M.M.; Wittrup, K.D. A modeling analysis of the effects of molecular size and binding affinity on tumor targeting. Mol. Cancer Ther. 2009, 8, 2861–2871. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, V.M.; Campani, G.; Santos, M.P.; Silva, G.G.; Pires, M.C.; Gonçalves, V.M.; de Giordano, R.C.; Sargo, C.R.; Horta, A.C.L.; Zangirolami, T.C. Cost analysis based on bioreactor cultivation conditions: Production of a soluble recombinant protein using Escherichia coli BL21(DE3). Biotechnol. Rep. 2020, 26, e00441. [Google Scholar] [CrossRef]
- Perols, A.; Karlström, A.E. Site-specific photoconjugation of antibodies using chemically synthesized IgG-binding domains. Bioconjug. Chem. 2014, 25, 481–488. [Google Scholar] [CrossRef]
- Ding, H.; Xu, T.; Zhang, J.; Tolmachev, V.; Oroujeni, M.; Orlova, A.; Gräslund, T.; Vorobyeva, A. Affibody-derived drug conjugates targeting HER2: Effect of drug load on cytotoxicity and biodistribution. Pharmaceutics 2021, 13, 430. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Temperature (°C) | Analyte | ka (1/Ms) | kd (1/s) | KD (nM) |
|---|---|---|---|---|---|
| ZHER3-ABD-mcDM1 | 25 | HER3 | 4.3 × 104 | 2.5 × 10−4 | 6 |
| ZHER3-ABD-AA | 25 | HER3 | 1.4 × 105 | 2.5 × 10−4 | 2 |
| ZHER3-ABD-mcDM1 | 25 | mErbB3 | 1.1 × 105 | 5.7 × 10−4 | 5 |
| ZHER3-ABD-AA | 25 | mErbB3 | 1.0 × 105 | 4.7 × 10−4 | 5 |
| ZHER3-ABD-mcDM1 * | 25 | HER3 | 6.0 × 104 | 2.2 × 10−4 | 4 |
| ZHER3-ABD-AA * | 25 | HER3 | 1.5 × 105 | 2.3 × 10−4 | 2 |
| ZHER3-ABD-mcDM1 * | 25 | mErbB3 | 5.6 × 104 | 4.3 × 10−4 | 8 |
| ZHER3-ABD-AA * | 25 | mErbB3 | 1.0 × 105 | 4.7 × 10−4 | 5 |
| ZHER3-ABD-mcDM1 | 37 | HER3 | 1.3 × 105 | 1.5 × 10−3 | 10 |
| ZHER3-ABD-AA | 37 | HER3 | 2.2 × 105 | 1.2 × 10−3 | 5 |
| ZHER3-ABD-mcDM1 | 37 | mErbB3 | 2.7 × 104 | 2.7 × 10−3 | 100 |
| ZHER3-ABD-AA | 37 | mErbB3 | 1.2 × 105 | 2.7 × 10−3 | 20 |
| ZHER3-ABD-mcDM1 * | 37 | HER3 | 1.6 × 105 | 1.8 × 10−3 | 10 |
| ZHER3-ABD-AA * | 37 | HER3 | 2.4 × 105 | 1.2 × 10−3 | 5 |
| ZHER3-ABD-mcDM1 * | 37 | mErbB3 | 6.6 × 104 | 3.7 × 10−3 | 60 |
| ZHER3-ABD-AA * | 37 | mErbB3 | 9.2 × 104 | 2.4 × 10−3 | 30 |
| Condition | [99mTc]Tc-ZHER3-ABD-mcDM1 | [99mTc]Tc-ZHER3-ABD-AA | ||
|---|---|---|---|---|
| 1 h | 4 h | 1 h | 4 h | |
| 500× histidine, 22 °C | 1.4 ± 0.5 * | 2.3 ± 0.1 | 1.3 ± 0.1 | 2.0 ± 0.4 |
| 500× histidine, 37 °C | 4.0 ± 0.5 | 8.8 ± 0.6 | 4.3 ± 0.5 | 5.7 ± 0.2 |
| PBS, 22 °C | 1.3 ± 0.9 | 0.9 ± 0.5 | 1.0 ± 0.6 | 1.0 ± 0.5 |
| Construct | ka (1/Ms) a | kd (1/s) | KD (nM) |
|---|---|---|---|
| [99mTc]Tc-ZHER3-ABD-mcDM1 | 1.4 × 105 ± 0.1 × 105 | 3.0 × 10−5 ± 2.0 × 10−5 | 0.2 ± 0.1 |
| [99mTc]Tc-ZHER3-ABD-AA | 1.8 × 105 ± 0.1 × 105 | 4.0 × 10−5 ± 3.0 × 10−5 | 0.2 ± 0.1 |
| [99mTc]Tc-(HE)3-ZHER3 | 3.0 × 105 ± 0.4 × 105 | 1.1 × 10−5 ± 0.1 × 10−5 | 0.04 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinne, S.S.; Yin, W.; Borras, A.M.; Abouzayed, A.; Leitao, C.D.; Vorobyeva, A.; Löfblom, J.; Ståhl, S.; Orlova, A.; Gräslund, T. Targeting Tumor Cells Overexpressing the Human Epidermal Growth Factor Receptor 3 with Potent Drug Conjugates Based on Affibody Molecules. Biomedicines 2022, 10, 1293. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061293
Rinne SS, Yin W, Borras AM, Abouzayed A, Leitao CD, Vorobyeva A, Löfblom J, Ståhl S, Orlova A, Gräslund T. Targeting Tumor Cells Overexpressing the Human Epidermal Growth Factor Receptor 3 with Potent Drug Conjugates Based on Affibody Molecules. Biomedicines. 2022; 10(6):1293. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061293
Chicago/Turabian StyleRinne, Sara S., Wen Yin, Anna Mestre Borras, Ayman Abouzayed, Charles Dahlsson Leitao, Anzhelika Vorobyeva, John Löfblom, Stefan Ståhl, Anna Orlova, and Torbjörn Gräslund. 2022. "Targeting Tumor Cells Overexpressing the Human Epidermal Growth Factor Receptor 3 with Potent Drug Conjugates Based on Affibody Molecules" Biomedicines 10, no. 6: 1293. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061293