
Effects of Extracorporeal Shockwave Therapy on Functional Recovery and Circulating miR-375 and miR-382-5p after Subacute and Chronic Spinal Cord Contusion Injury in Rats

 , , , and
, , , and 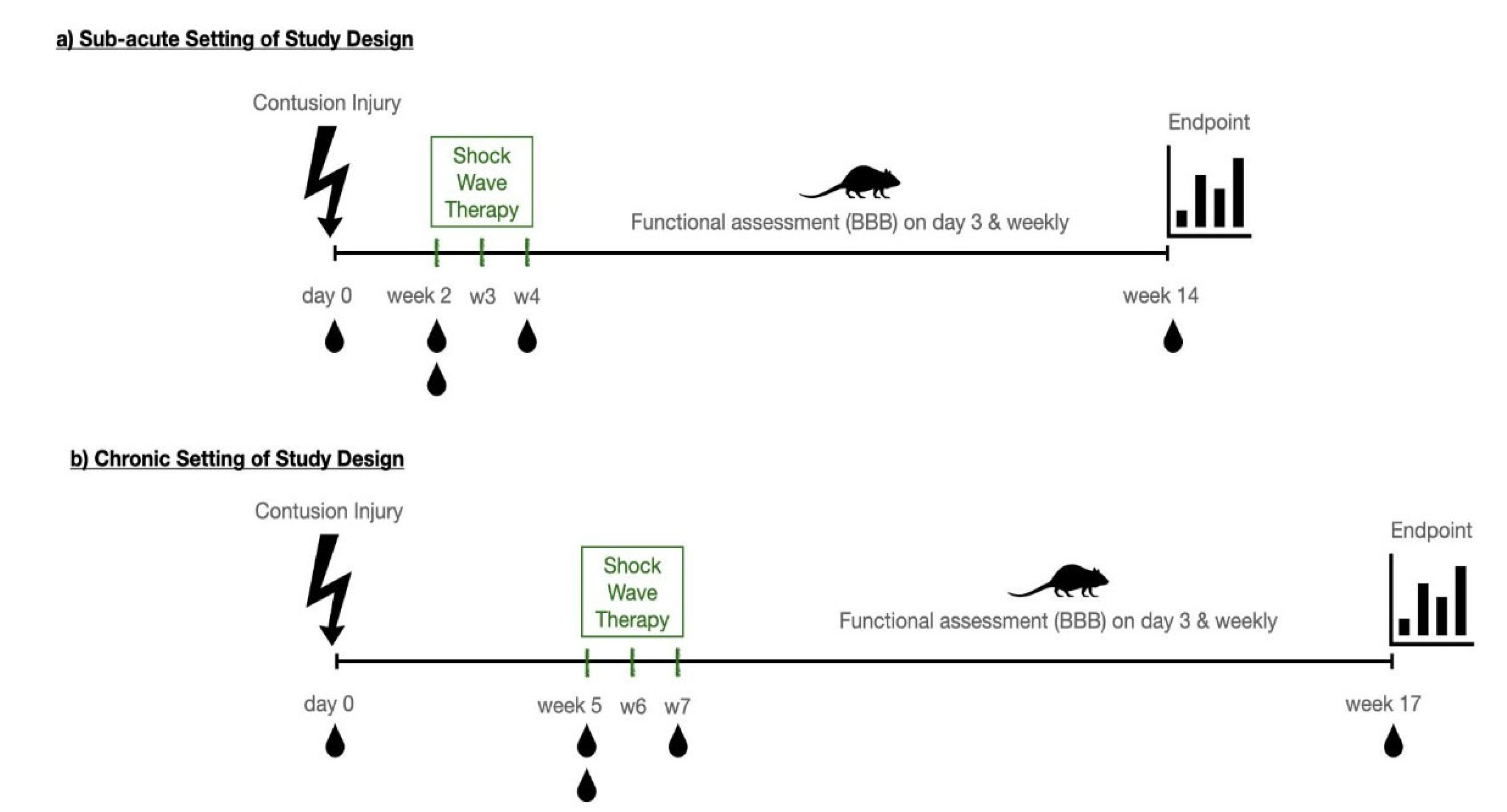{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Surgery
2.2. Surgical Procedure
2.3. ESWT Setup
2.4. Functional Testing—The BBB-Score
2.5. Histology and Morphometric Analysis
2.5.1. µ-CT
2.5.2. Immunohistochemistry
2.6. MicroRNA Sampling and Analysis
2.6.1. Serum RNA Extraction
2.6.2. MicroRNA Reverse-Transcription Quantitative PCR Analysis in Serum RNA (RT-qPCR)
2.6.3. Principal Component Analysis (PCA) of miRNAs
2.6.4. Hypothesis Tests for Diagnostic miRNAs and Treatment Effect
2.6.5. Correlation of Functional Scoring and miRNA Levels
3. Results
3.1. ESWT Significantly Improved Motor Function after SCI
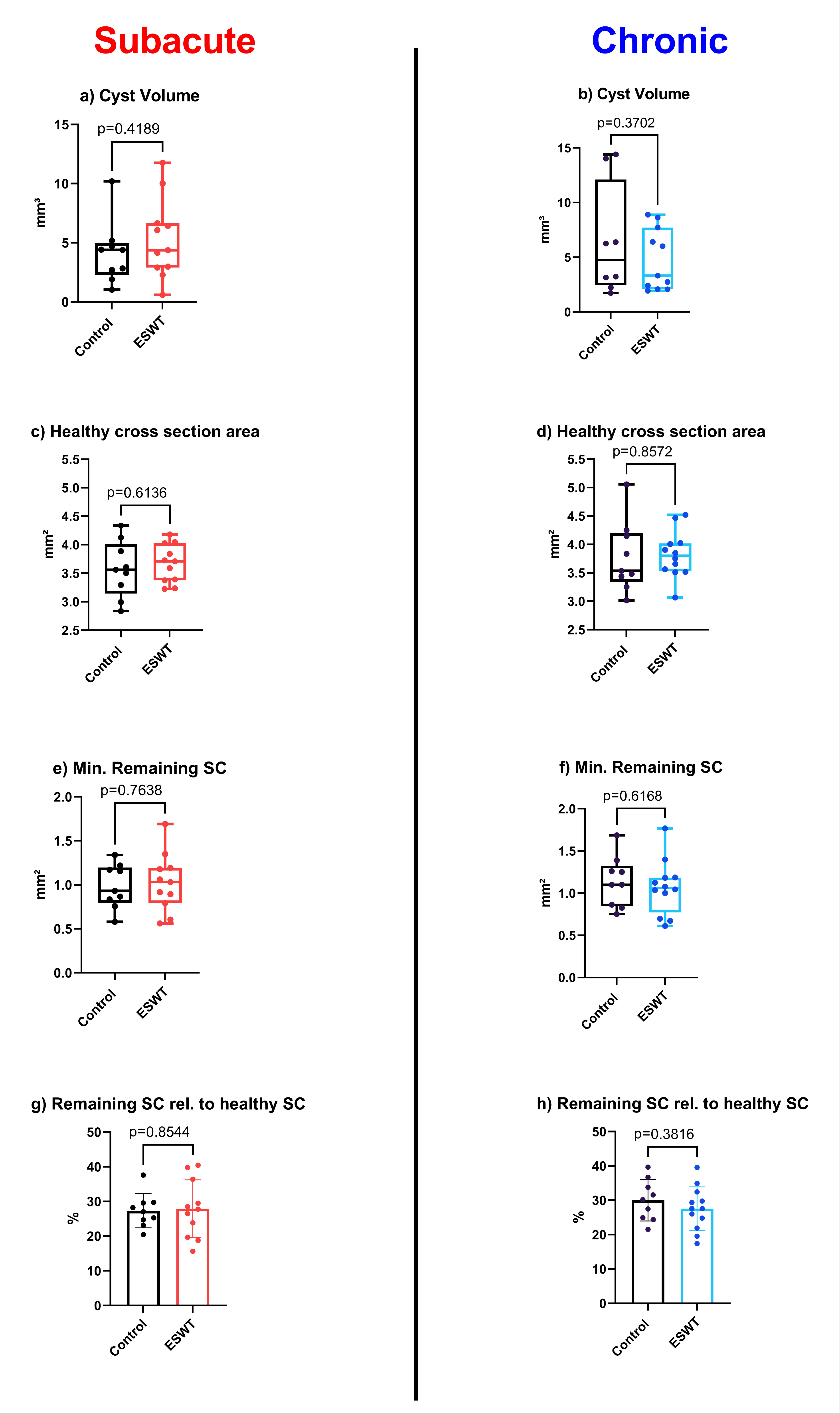
3.2. Calculated 3D Parameters Generated by µCT-Scans Did Not Show Significant Morphological Differences between Therapy and Control Groups
3.3. Histological and Immunohistochemical Analysis Did Not Reveal Significant Differences after ESWT in Comparison with Control Group
3.4. ESWT Significantly Influenced the Expression Patterns of Systemic miRNA
3.4.1. Several Identified miRNAs Showed Potential Therapeutic Relevance in Either Subacute or Chronic Experimental Setting
3.4.2. A Sub-Set of miRNAs Correlated with Functional Outcome
3.4.3. PCA of microRNA Expression Data Revealed Highest Relative Feature Importance of miRNA 382-5p across Both Treatment Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Furia, J.P.; Juliano, P.J.; Wade, A.M.; Schaden, W.; Mittermayr, R. Shock wave therapy compared with intramedullary screw fixation for nonunion of proximal fifth metatarsal metaphyseal-diaphyseal fractures. J. Bone. Joint Surg. Am. 2010, 92, 846–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, S.K.; Kailash. Shock wave treatment in medicine. J. Biosci. 2005, 30, 269–275. [Google Scholar] [CrossRef]
- Wang, C.J. Extracorporeal shockwave therapy in musculoskeletal disorders. J. Orthop. Surg. Res. 2012, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, T.; Cai, H.Y.; Ma, T.K.; Tao, S.M.; Sun, S.; Chen, M.Q.; Gu, Y.; Pang, J.H.; Xiao, J.M.; et al. Cardiac shock wave therapy reduces angina and improves myocardial function in patients with refractory coronary artery disease. Clin. Cardiol. 2010, 33, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Ito, K.; Shiroto, T.; Tsuburaya, R.; Yi, G.J.; Takeda, M.; Fukumoto, Y.; Yasuda, S.; Shimokawa, H. Cardiac shock wave therapy ameliorates left ventricular remodeling after myocardial ischemia-reperfusion injury in pigs in vivo. Coron. Artery Dis. 2010, 21, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Mori, L.; Marinelli, L.; Pelosin, E.; Currà, A.; Molfetta, L.; Abbruzzese, G.; Trompetto, C. Shock waves in the treatment of muscle hypertonia and dystonia. Biomed. Res. Int. 2014, 2014, 637450. [Google Scholar] [CrossRef]
- Hausner, T.; Pajer, K.; Halat, G.; Hopf, R.; Schmidhammer, R.; Redl, H.; Nogradi, A. Improved rate of peripheral nerve regeneration induced by extracorporeal shock wave treatment in the rat. Exp. Neurol. 2012, 236, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Aicher, A.; Heeschen, C.; Sasaki, K.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Low-energy shock wave for enhancing recruitment of endothelial progenitor cells: A new modality to increase efficacy of cell therapy in chronic hind limb ischemia. Circulation 2006, 114, 2823–2830. [Google Scholar] [CrossRef] [Green Version]
- El-Shamy, S.M.; Eid, M.A.; El-Banna, M.F. Effect of extracorporeal shock wave therapy on gait pattern in hemiplegic cerebral palsy: A randomized controlled trial. Am. J. Phys. Med. Rehabil. 2014, 93, 1065–1072. [Google Scholar] [CrossRef]
- Lohse-Busch, H.; Reime, U.; Falland, R. Symptomatic treatment of unresponsive wakefulness syndrome with transcranially focused extracorporeal shock waves. NeuroRehabilitation 2014, 35, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Lobenwein, D.; Tepeköylü, C.; Kozaryn, R.; Pechriggl, E.J.; Bitsche, M.; Graber, M.; Fritsch, H.; Semsroth, S.; Stefanova, N.; Paulus, P.; et al. Shock Wave Treatment Protects from Neuronal Degeneration via a Toll-Like Receptor 3 Dependent Mechanism: Implications of a First-Ever Causal Treatment for Ischemic Spinal Cord Injury. J. Am. Heart Assoc. 2015, 4, e002440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weihs, A.M.; Fuchs, C.; Teuschl, A.H.; Hartinger, J.; Slezak, P.; Mittermayr, R.; Redl, H.; Junger, W.G.; Sitte, H.H.; Runzler, D. Shock wave treatment enhances cell proliferation and improves wound healing by ATP release-coupled extracellular signal-regulated kinase (ERK) activation. J. Biol. Chem. 2014, 289, 27090–27104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuh, C.M.; Heher, P.; Weihs, A.M.; Banerjee, A.; Fuchs, C.; Gabriel, C.; Wolbank, S.; Mittermayr, R.; Redl, H.; Runzler, D.; et al. In vitro extracorporeal shock wave treatment enhances stemness and preserves multipotency of rat and human adipose-derived stem cells. Cytotherapy 2014, 16, 1666–1678. [Google Scholar] [CrossRef] [PubMed]
- Schuh, C.M.; Hercher, D.; Stainer, M.; Hopf, R.; Teuschl, A.H.; Schmidhammer, R.; Redl, H. Extracorporeal shockwave treatment: A novel tool to improve Schwann cell isolation and culture. Cytotherapy 2016, 18, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Gollmann-Tepeköylü, C.; Nägele, F.; Graber, M.; Pölzl, L.; Lobenwein, D.; Hirsch, J.; An, A.; Irschick, R.; Röhrs, B.; Kremser, C.; et al. Shock waves promote spinal cord repair via TLR3. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Yahata, K.; Kanno, H.; Ozawa, H.; Yamaya, S.; Tateda, S.; Ito, K.; Shimokawa, H.; Itoi, E. Low-energy extracorporeal shock wave therapy for promotion of vascular endothelial growth factor expression and angiogenesis and improvement of locomotor and sensory functions after spinal cord injury. J. Neurosurg. Spine 2016, 25, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Mittermayr, R.; Hartinger, J.; Antonic, V.; Meinl, A.; Pfeifer, S.; Stojadinovic, A.; Schaden, W.; Redl, H. Extracorporeal shock wave therapy (ESWT) minimizes ischemic tissue necrosis irrespective of application time and promotes tissue revascularization by stimulating angiogenesis. Ann. Surg. 2011, 253, 1024–1032. [Google Scholar] [CrossRef]
- Ogden, J.A.; Tóth-Kischkat, A.; Schultheiss, R. Principles of shock wave therapy. Clin. Orthop. Relat. Res. 2001, 8–17. [Google Scholar] [CrossRef]
- Krause, H. Physik und Technik Medizinischer Stoßwellensysteme. In Extrakorporale Stoßwellentherapie-Grundlagen, Indikation, Anwendung; Rompe, J.D., Ed.; Chapman & Hall: Weinheim, Germany, 1997. [Google Scholar]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Heneghan, H.M.; Miller, N.; Kelly, R.; Newell, J.; Kerin, M.J. Systemic miRNA-195 differentiates breast cancer from other malignancies and is a potential biomarker for detecting noninvasive and early stage disease. Oncologist 2010, 15, 673–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laterza, O.F.; Lim, L.; Garrett-Engele, P.W.; Vlasakova, K.; Muniappa, N.; Tanaka, W.K.; Johnson, J.M.; Sina, J.F.; Fare, T.L.; Sistare, F.D.; et al. Plasma MicroRNAs as sensitive and specific biomarkers of tissue injury. Clin. Chem. 2009, 55, 1977–1983. [Google Scholar] [CrossRef] [Green Version]
- Gasparri, M.L.; Casorelli, A.; Bardhi, E.; Besharat, A.R.; Savone, D.; Ruscito, I.; Farooqi, A.A.; Papadia, A.; Mueller, M.D.; Ferretti, E.; et al. Beyond circulating microRNA biomarkers: Urinary microRNAs in ovarian and breast cancer. Tumour Biol. 2017, 39, 1010428317695525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Xu, C.; Yuan, W.; Wang, C.; Zhao, P.; Chen, L.; Ma, J. Evaluation of Plasma MicroRNAs as Diagnostic and Prognostic Biomarkers in Pancreatic Adenocarcinoma: miR-196a and miR-210 Could Be Negative and Positive Prognostic Markers, Respectively. Biomed. Res. Int. 2017, 2017, 6495867. [Google Scholar] [CrossRef]
- Van Rossum, I.A.; Vos, S.; Handels, R.; Visser, P.J. Biomarkers as predictors for conversion from mild cognitive impairment to Alzheimer-type dementia: Implications for trial design. J. Alzheimers Dis. 2010, 20, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Lau, N.C.; Lim, L.P.; Weinstein, E.G.; Bartel, D.P. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 2001, 294, 858–862. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Zhou, H.; Lu, L.; Li, X.; Fu, Z.; Liu, J.; Kang, Y.; Wei, Z.; Pan, B.; Liu, L.; et al. The roles of microRNAs in spinal cord injury. Int. J. Neurosci. 2017, 127, 1104–1115. [Google Scholar] [CrossRef]
- Bhinge, A.; Namboori, S.C.; Bithell, A.; Soldati, C.; Buckley, N.J.; Stanton, L.W. MiR-375 is Essential for Human Spinal Motor Neuron Development and May Be Involved in Motor Neuron Degeneration. Stem Cells 2016, 34, 124–134. [Google Scholar] [CrossRef]
- Xiang, W.; Jiang, L.; Zhou, Y.; Li, Z.; Zhao, Q.; Wu, T.; Cao, Y.; Zhou, J. The lncRNA Ftx/miR-382-5p/Nrg1 axis improves the inflammation response of microglia and spinal cord injury repair. Neurochem. Int. 2021, 143, 104929. [Google Scholar] [CrossRef] [PubMed]
- Gollmann-Tepeköylü, C.; Pölzl, L.; Graber, M.; Hirsch, J.; Nägele, F.; Lobenwein, D.; Hess, M.W.; Blumer, M.J.; Kirchmair, E.; Zipperle, J.; et al. miR-19a-3p containing exosomes improve function of ischaemic myocardium upon shock wave therapy. Cardiovasc. Res. 2019, 116, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-C.; Huang, H.-Y.; Yang, Y.-L.; Hung, K.-S.; Cheng, C.-H.; Chang, N.-K.; Chung, Y.-H.; Hu, M.-S.; Wang, C.-J. Vulnerability of the spinal cord to injury from extracorporeal shock waves in rabbits. J. Clin. Neurosci. 2007, 14, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Yamaya, S.; Ozawa, H.; Kanno, H.; Kishimoto, K.N.; Sekiguchi, A.; Tateda, S.; Yahata, K.; Ito, K.; Shimokawa, H.; Itoi, E. Low-energy extracorporeal shock wave therapy promotes vascular endothelial growth factor expression and improves locomotor recovery after spinal cord injury. J. Neurosurg. 2014, 121, 1514–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, J.S.; Soon, P.S.; Marsh, D.J. Comparison of Methodologies to Detect Low Levels of Hemolysis in Serum for Accurate Assessment of Serum microRNAs. PLoS ONE 2016, 11, e0153200. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 29 February 2020).
- Hastie, T. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Heimel, P.; Swiadek, N.V.; Slezak, P.; Kerbl, M.; Schneider, C.; Nürnberger, S.; Redl, H.; Teuschl, A.H.; Hercher, D. Iodine-Enhanced Micro-CT Imaging of Soft Tissue on the Example of Peripheral Nerve Regeneration. Contrast Media Mol. Imaging 2019, 2019, 7483745. [Google Scholar] [CrossRef] [Green Version]
- Alavi, S.N.R.; Neishaboori, A.M.; Yousefifard, M. Extracorporeal shockwave therapy in spinal cord injury, early to advance to clinical trials? A systematic review and meta-analysis on animal studies. Neuroradiol. J. 2021, 34, 552–561. [Google Scholar] [CrossRef]
- Cheng, J.H.; Wang, C.J.; Su, S.H.; Huang, C.Y.; Hsu, S.L. Next-generation sequencing identifies articular cartilage and subchondral bone miRNAs after ESWT on early osteoarthritis knee. Oncotarget 2016, 7, 84398–84407. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Shen, T.; Wang, Q.; Yu, X.; Jia, N.; He, Q. Cardiac shock wave therapy protects cardiomyocytes from hypoxia-induced injury by modulating miR-210. Mol. Med. Rep. 2020, 21, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Gazdarica, M.; Noda, J.; Durydivka, O.; Novosadova, V.; Mackie, K.; Pin, J.P.; Prezeau, L.; Blahos, J. SGIP1 modulates kinetics and interactions of the cannabinoid receptor 1 and G protein-coupled receptor kinase 3 signalosome. J. Neurochem. 2022, 160, 625–642. [Google Scholar] [CrossRef] [PubMed]
- Hájková, A.; Techlovská, Š.; Dvořáková, M.; Chambers, J.N.; Kumpošt, J.; Hubálková, P.; Prezeau, L.; Blahos, J. SGIP1 alters internalization and modulates signaling of activated cannabinoid receptor 1 in a biased manner. Neuropharmacology 2016, 107, 201–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Shi, X.; Zhao, J.; He, J.; Li, K.; Cen, Z.; Wu, Y.; Zong, S.; Zeng, G. The Effects of miR-136-5p-Mediated Regulation of A20 in Astrocytes from Cultured Spinal Cord Cultured Cells In Vitro. Cell Physiol. Biochem. 2017, 41, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhao, J.; Peng, X.; Shi, X.; Zong, S.; Zeng, G. Molecular Mechanism of MiR-136-5p Targeting NF-κB/A20 in the IL-17-Mediated Inflammatory Response after Spinal Cord Injury. Cell Physiol. Biochem 2017, 44, 1224–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, G.; Gao, Y.; Cen, Z.; He, J.; Cao, B.; Zeng, G.; Zong, S. miR-136-5p Regulates the Inflammatory Response by Targeting the IKKβ/NF-κB/A20 Pathway After Spinal Cord Injury. Cell Physiol. Biochem. 2018, 50, 512–524. [Google Scholar] [CrossRef]
- Jin, R.; Xu, S.; Lin, X.; Shen, M. MiR-136 controls neurocytes apoptosis by regulating Tissue Inhibitor of Metalloproteinases-3 in spinal cord ischemic injury. Biomed. Pharmacother. 2017, 94, 47–54. [Google Scholar] [CrossRef]
- Fan, Y.; Wu, Y. Tetramethylpyrazine alleviates neural apoptosis in injured spinal cord via the downregulation of miR-214-3p. Biomed. Pharmacother. 2017, 94, 827–833. [Google Scholar] [CrossRef]
- Che, H.; Li, H.; Li, Y.; Wang, Y.Q.; Yang, Z.Y.; Wang, R.L.; Wang, L.H. Melatonin exerts neuroprotective effects by inhibiting neuronal pyroptosis and autophagy in STZ-induced diabetic mice. FASEB J. 2020, 34, 14042–14054. [Google Scholar] [CrossRef]
- Pang, B.; Qiao, L.; Wang, S.; Guo, X.; Xie, Y.; Han, L. MiR-214-3p plays a protective role in diabetic neuropathic rats by regulating Nav1. 3 and TLR4. Cell Biol. Int. 2021, 45, 2294–2303. [Google Scholar] [CrossRef]
- Irie, K.; Tsujimura, K.; Nakashima, H.; Nakashima, K. MicroRNA-214 Promotes Dendritic Development by Targeting the Schizophrenia-associated Gene Quaking (Qki). J. Biol. Chem. 2016, 291, 13891–13904. [Google Scholar] [CrossRef] [Green Version]
- Shu, P.; Fu, H.; Zhao, X.; Wu, C.; Ruan, X.; Zeng, Y.; Liu, W.; Wang, M.; Hou, L.; Chen, P.; et al. MicroRNA-214 modulates neural progenitor cell differentiation by targeting Quaking during cerebral cortex development. Sci. Rep. 2017, 7, 8014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xu, D.; Wang, T.; Zhang, Y.; Yang, X.; Wang, X.; Tang, Y. Epigenetic reduction of miR-214-3p upregulates astrocytic colony-stimulating factor-1 and contributes to neuropathic pain induced by nerve injury. Pain 2020, 161, 96–108. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, H.; Yin, H.; Geng, S.; Liu, Y.; Liu, C.; Zhao, J.; Liu, Y.; Wang, X.; Wang, Y. Up-regulated microRNA-218-5p ameliorates the damage of dopaminergic neurons in rats with Parkinson’s disease via suppression of LASP1. Brain Res. Bull. 2021, 166, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Mosarrezaii Aghdam, A.; Rezaei, S.; Zarza Nalivan, F.; Babaie, F.; Amiri Nikpour, M.R.; Torkamandi, S. Downregulation of miR-125a-5p and miR-218-5p in Peripheral Blood Mononuclear Cells of Patients with Relapsing-Remitting Multiple Sclerosis. Immunol. Investig. 2022, 51, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.N.; Lee, S.J.; Sahoo, P.K.; Thames, E.; Yoo, S.; Houle, J.D.; Twiss, J.L. MicroRNAs 21 and 199a-3p Regulate Axon Growth Potential through Modulation of Pten and mTor mRNAs. eNeuro 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Detloff, M.R.; Miller, K.N.; Santi, L.; Houlé, J.D. Exercise modulates microRNAs that affect the PTEN/mTOR pathway in rats after spinal cord injury. Exp. Neurol. 2012, 233, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Han, J.; Wang, D. Identification of key microRNAs and the underlying molecular mechanism in spinal cord ischemia-reperfusion injury in rats. PeerJ 2021, 9, e11454. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lai, X.; Wu, D.; Liu, B.; Wang, N.; Rong, L. Umbilical mesenchymal stem cell-derived exosomes facilitate spinal cord functional recovery through the miR-199a-3p/145-5p-mediated NGF/TrkA signaling pathway in rats. Stem Cell Res. Ther. 2021, 12, 117. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Zhang, D.; Li, J.; Cui, S. MiR-375 potentially enhances GnRH expression by targeting Sp1 in GT1-7 cells. In Vitro Cell Dev. Biol. Anim. 2021, 57, 438–447. [Google Scholar] [CrossRef]
- Díaz-Galindo, M.D.C.; Calderón-Vallejo, D.; Olvera-Sandoval, C.; Quintanar, J.L. Therapeutic approaches of trophic factors in animal models and in patients with spinal cord injury. Growth Factors 2020, 38, 1–15. [Google Scholar] [CrossRef]
- Quintanar, J.L.; Salinas, E.; González, R. Gonadotropin-releasing hormone receptor in spinal cord neurons of embryos and adult rats. Neurosci. Lett. 2009, 461, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Díaz Galindo, C.; Gómez-González, B.; Salinas, E.; Calderón-Vallejo, D.; Hernández-Jasso, I.; Bautista, E.; Quintanar, J.L. Leuprolide acetate induces structural and functional recovery of injured spinal cord in rats. Neural Regen. Res. 2015, 10, 1819–1824. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Vallejo, D.; Quintanar-Stephano, A.; Hernández-Jasso, I.; Jiménez-Hernández, V.; Ruiz-Ornelas, J.; Jiménez, I.; Quintanar, J.L. Functional and structural recovery of the injured spinal cord in rats treated with gonadotropin-releasing hormone. Neurochem. Res. 2015, 40, 455–462. [Google Scholar] [CrossRef]
- Quintanar, J.L.; Díaz-Galindo, C.; Calderón-Vallejo, D.; Hernández-Jasso, I.; Rojas, F.; Medina-Aguiñaga, D.; Olvera-Sandoval, C. Neurological improvement in patients with chronic spinal cord injury treated with leuprolide acetate, an agonist of GnRH. Acta Neurobiol. Exp. 2018, 78, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.Q.; Chen, J.; Wang, S.N.; Duan, F.X.; Chen, Y.Q.; Shi, Y.J.; Hu, J.G.; Lü, H.Z. Identification of serum exosomal microRNAs in acute spinal cord injured rats. Exp. Biol. Med. 2019, 244, 1149–1161. [Google Scholar] [CrossRef]
- Luchetti, A.; Ciafrè, S.A.; Murdocca, M.; Malgieri, A.; Masotti, A.; Sanchez, M.; Farace, M.G.; Novelli, G.; Sangiuolo, F. A Perturbed MicroRNA Expression Pattern Characterizes Embryonic Neural Stem Cells Derived from a Severe Mouse Model of Spinal Muscular Atrophy (SMA). Int. J. Mol. Sci. 2015, 16, 18312–18327. [Google Scholar] [CrossRef] [Green Version]
- Díez-Planelles, C.; Sánchez-Lozano, P.; Crespo, M.C.; Gil-Zamorano, J.; Ribacoba, R.; González, N.; Suárez, E.; Martínez-Descals, A.; Martínez-Camblor, P.; Álvarez, V.; et al. Circulating microRNAs in Huntington’s disease: Emerging mediators in metabolic impairment. Pharmacol. Res. 2016, 108, 102–110. [Google Scholar] [CrossRef]
- Aganzo, M.; Montojo, M.T.; López de Las Hazas, M.C.; Martínez-Descals, A.; Ricote-Vila, M.; Sanz, R.; González-Peralta, I.; Martín-Hernández, R.; de Dios, O.; Garcés, C.; et al. Customized Dietary Intervention Avoids Unintentional Weight Loss and Modulates Circulating miRNAs Footprint in Huntington’s Disease. Mol. Nutr. Food Res. 2018, 62, e1800619. [Google Scholar] [CrossRef]
- Wallach, T.; Mossmann, Z.J.; Szczepek, M.; Wetzel, M.; Machado, R.; Raden, M.; Miladi, M.; Kleinau, G.; Krüger, C.; Dembny, P.; et al. MicroRNA-100-5p and microRNA-298-5p released from apoptotic cortical neurons are endogenous Toll-like receptor 7/8 ligands that contribute to neurodegeneration. Mol. Neurodegener. 2021, 16, 80. [Google Scholar] [CrossRef]
- Serpente, M.; Fenoglio, C.; D’Anca, M.; Arcaro, M.; Sorrentino, F.; Visconte, C.; Arighi, A.; Fumagalli, G.G.; Porretti, L.; Cattaneo, A.; et al. MiRNA Profiling in Plasma Neural-Derived Small Extracellular Vesicles from Patients with Alzheimer’s Disease. Cells 2020, 9, 1443. [Google Scholar] [CrossRef]
- Fang, Y.; Hong, X. miR-124-3p Inhibits Microglial Secondary Inflammation After Basal Ganglia Hemorrhage by Targeting TRAF6 and Repressing the Activation of NLRP3 Inflammasome. Front. Neurol. 2021, 12, 653321. [Google Scholar] [CrossRef] [PubMed]
- Vuokila, N.; Aronica, E.; Korotkov, A.; van Vliet, E.A.; Nuzhat, S.; Puhakka, N.; Pitkänen, A. Chronic Regulation of miR-124-3p in the Perilesional Cortex after Experimental and Human TBI. Int. J. Mol. Sci. 2020, 21, 2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuokila, N.; Lukasiuk, K.; Bot, A.M.; van Vliet, E.A.; Aronica, E.; Pitkänen, A.; Puhakka, N. miR-124-3p is a chronic regulator of gene expression after brain injury. Cell Mol. Life Sci. 2018, 75, 4557–4581. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, G.C.; Smothers, C.G.; Winkelman, C. Bioinformatic analysis of brain-specific miRNAs for identification of candidate traumatic brain injury blood biomarkers. Brain Inj. 2020, 34, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhao, K.; Ruan, Q.; Meng, C.; Yin, F. Bone marrow mesenchymal stem cell-derived exosomal microRNA-124-3p attenuates neurological damage in spinal cord ischemia-reperfusion injury by downregulating Ern1 and promoting M2 macrophage polarization. Arthritis Res. Ther. 2020, 22, 75. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Gong, F.; Ge, X.; Lv, C.; Huang, C.; Feng, S.; Zhou, Z.; Rong, Y.; Wang, J.; Ji, C.; et al. Neuron-derived exosomes-transmitted miR-124-3p protect traumatically injured spinal cord by suppressing the activation of neurotoxic microglia and astrocytes. J. Nanobiotechnol. 2020, 18, 105. [Google Scholar] [CrossRef]
- Ge, X.; Guo, M.; Hu, T.; Li, W.; Huang, S.; Yin, Z.; Li, Y.; Chen, F.; Zhu, L.; Kang, C.; et al. Increased Microglial Exosomal miR-124-3p Alleviates Neurodegeneration and Improves Cognitive Outcome after rmTBI. Mol. Ther. 2020, 28, 503–522. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Ye, Y.; Yang, Y.; Zhang, K.; Bai, W.; Chen, H.; Kang, E.; Kong, C.; He, X. The Effect of SPTLC2 on Promoting Neuronal Apoptosis is Alleviated by MiR-124-3p Through TLR4 Signalling Pathway. Neurochem. Res. 2019, 44, 2113–2122. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A.E. microRNA dysregulation in neurodegenerative diseases: A systematic review. Prog. Neurobiol. 2019, 182, 101664. [Google Scholar] [CrossRef]
- Vuokila, N.; Das Gupta, S.; Huusko, R.; Tohka, J.; Puhakka, N.; Pitkänen, A. Elevated Acute Plasma miR-124-3p Level Relates to Evolution of Larger Cortical Lesion Area after Traumatic Brain Injury. Neuroscience 2020, 433, 21–35. [Google Scholar] [CrossRef]
- Huang, S.; Ge, X.; Yu, J.; Han, Z.; Yin, Z.; Li, Y.; Chen, F.; Wang, H.; Zhang, J.; Lei, P. Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons. FASEB J. 2018, 32, 512–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.C.; Cheng, J.T.; Li, T.Y.; Tan, P.H. Integrated analysis of microRNA and mRNA expression profiles in the rat spinal cord under inflammatory pain conditions. Eur. J. Neurosci. 2017, 46, 2713–2728. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Liu, W.; Chen, Y. miR-124-3p attenuates MPP(+)-induced neuronal injury by targeting STAT3 in SH-SY5Y cells. Exp. Biol. Med. 2017, 242, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Rainer, T.H.; Leung, L.Y.; Chan, C.P.Y.; Leung, Y.K.; Abrigo, J.M.; Wang, D.; Graham, C.A. Plasma miR-124-3p and miR-16 concentrations as prognostic markers in acute stroke. Clin. Biochem. 2016, 49, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Jiang, C.; Sun, R.; Yang, D.; Liu, Q. Circular Noncoding RNA NR3C1 Acts as a miR-382-5p Sponge to Protect RPE Functions via Regulating PTEN/AKT/mTOR Signaling Pathway. Mol. Ther. 2020, 28, 929–945. [Google Scholar] [CrossRef]
- Li, S.; Ma, H.; Yuan, X.; Zhou, X.; Wan, Y.; Chen, S. MicroRNA-382-5p Targets Nuclear Receptor Subfamily 3 Group C Member 1 to Regulate Depressive-Like Behaviors Induced by Chronic Unpredictable Mild Stress in Rats. Neuropsychiatr. Dis. Treat. 2020, 16, 2053–2061. [Google Scholar] [CrossRef]
- Garraway, S.M.; Huie, J.R. Spinal Plasticity and Behavior: BDNF-Induced Neuromodulation in Uninjured and Injured Spinal Cord. Neural. Plast. 2016, 2016, 9857201. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Wang, X.; Chen, S. Downregulation of MiR-218-5p Protects Against Oxygen-Glucose Deprivation/Reperfusion-Induced Injuries of PC12 Cells via Upregulating N-myc Downstream Regulated Gene 4 (NDRG4). Med. Sci. Monit. 2020, 26, e920101. [Google Scholar] [CrossRef]
- Jin, F.; Xing, J. Circulating miR-126 and miR-130a levels correlate with lower disease risk, disease severity, and reduced inflammatory cytokine levels in acute ischemic stroke patients. Neurol. Sci. 2018, 39, 1757–1765. [Google Scholar] [CrossRef]
- Paim, L.R.; Schreiber, R.; de Rossi, G.; Matos-Souza, J.R.; Costa, E.S.A.A.; Calegari, D.R.; Cheng, S.; Marques, F.Z.; Sposito, A.C.; Gorla, J.I.; et al. Circulating microRNAs, Vascular Risk, and Physical Activity in Spinal Cord-Injured Subjects. J. Neurotrauma 2019, 36, 845–852. [Google Scholar] [CrossRef]
- Tigchelaar, S.; Streijger, F.; Sinha, S.; Flibotte, S.; Manouchehri, N.; So, K.; Shortt, K.; Okon, E.; Rizzuto, M.A.; Malenica, I.; et al. Serum MicroRNAs Reflect Injury Severity in a Large Animal Model of Thoracic Spinal Cord Injury. Sci. Rep. 2017, 7, 1376. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, P.; Yao, X.; Li, H.; Shen, H.; Li, X.; Wu, J.; Lu, X. Exosomes Derived From miR-133b-Modified Mesenchymal Stem Cells Promote Recovery After Spinal Cord Injury. Front. Neurosci. 2018, 12, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, D.; Liu, W.; Zhu, S.; Fan, B.; Yu, N.; Ning, G.; Feng, S. Potential of different cells-derived exosomal microRNA cargos for treating spinal cord injury. J. Orthop. Translat. 2021, 31, 33–40. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashmwe, M.; Posa, K.; Rührnößl, A.; Heinzel, J.C.; Heimel, P.; Mock, M.; Schädl, B.; Keibl, C.; Couillard-Despres, S.; Redl, H.; et al. Effects of Extracorporeal Shockwave Therapy on Functional Recovery and Circulating miR-375 and miR-382-5p after Subacute and Chronic Spinal Cord Contusion Injury in Rats. Biomedicines 2022, 10, 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071630
Ashmwe M, Posa K, Rührnößl A, Heinzel JC, Heimel P, Mock M, Schädl B, Keibl C, Couillard-Despres S, Redl H, et al. Effects of Extracorporeal Shockwave Therapy on Functional Recovery and Circulating miR-375 and miR-382-5p after Subacute and Chronic Spinal Cord Contusion Injury in Rats. Biomedicines. 2022; 10(7):1630. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071630
Chicago/Turabian StyleAshmwe, Mohamed, Katja Posa, Alexander Rührnößl, Johannes Christoph Heinzel, Patrick Heimel, Michael Mock, Barbara Schädl, Claudia Keibl, Sebastien Couillard-Despres, Heinz Redl, and et al. 2022. "Effects of Extracorporeal Shockwave Therapy on Functional Recovery and Circulating miR-375 and miR-382-5p after Subacute and Chronic Spinal Cord Contusion Injury in Rats" Biomedicines 10, no. 7: 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10071630