
Osteoblast Secretome Modulated by Abiraterone Treatment Affects Castration Resistant Prostate Cancer Cell Proliferation
 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prostate Cancer Cell Line
2.2. Human Osteoblasts
2.3. Osteoblast Conditioned Media
2.4. AR Activity Assay
2.5. Cell Cycle Analysis
2.6. Proteomic Assay
2.7. Statistical Analysis
3. Results
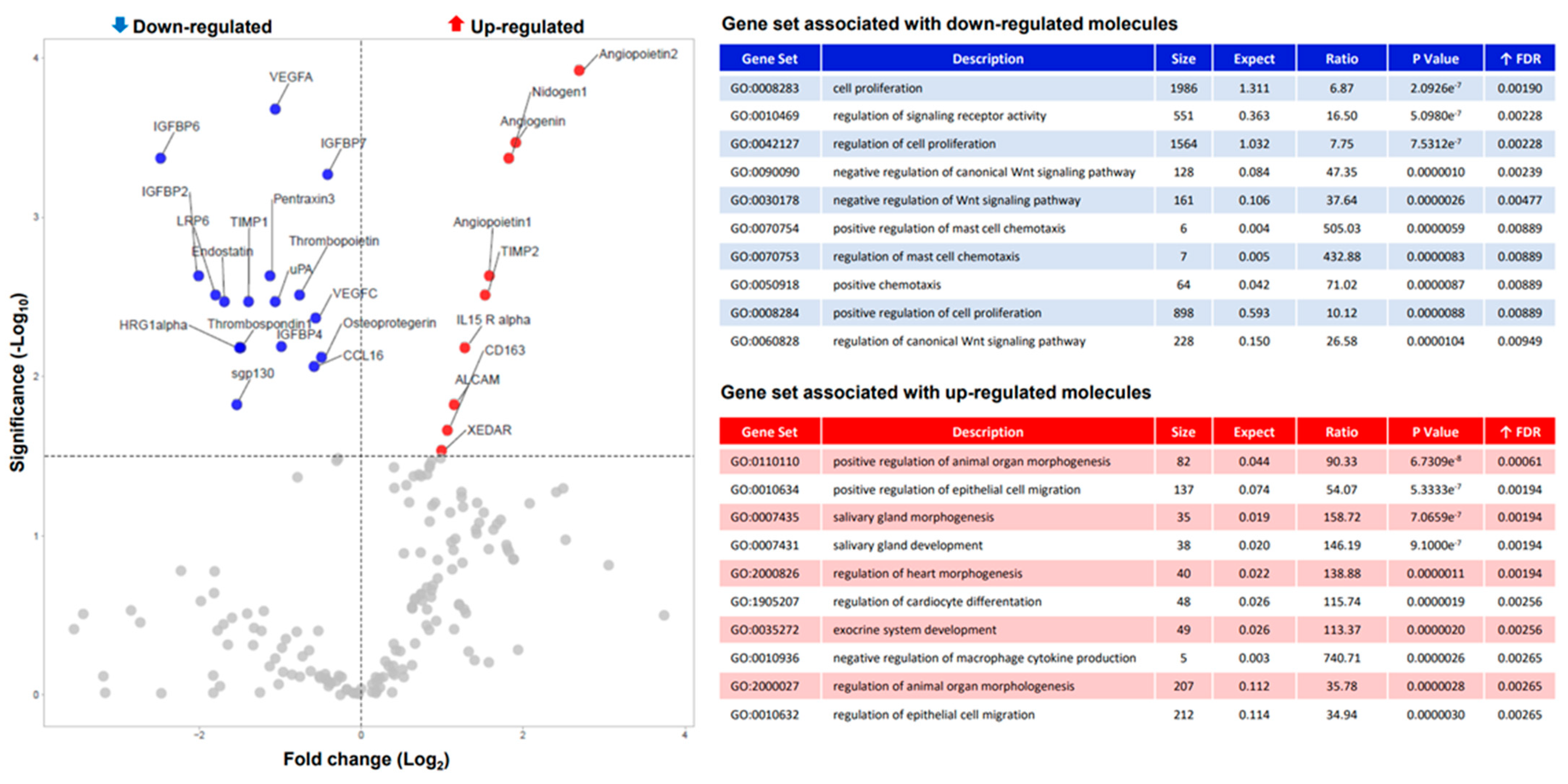
3.1. ABI Modulate Osteoblast Secretome
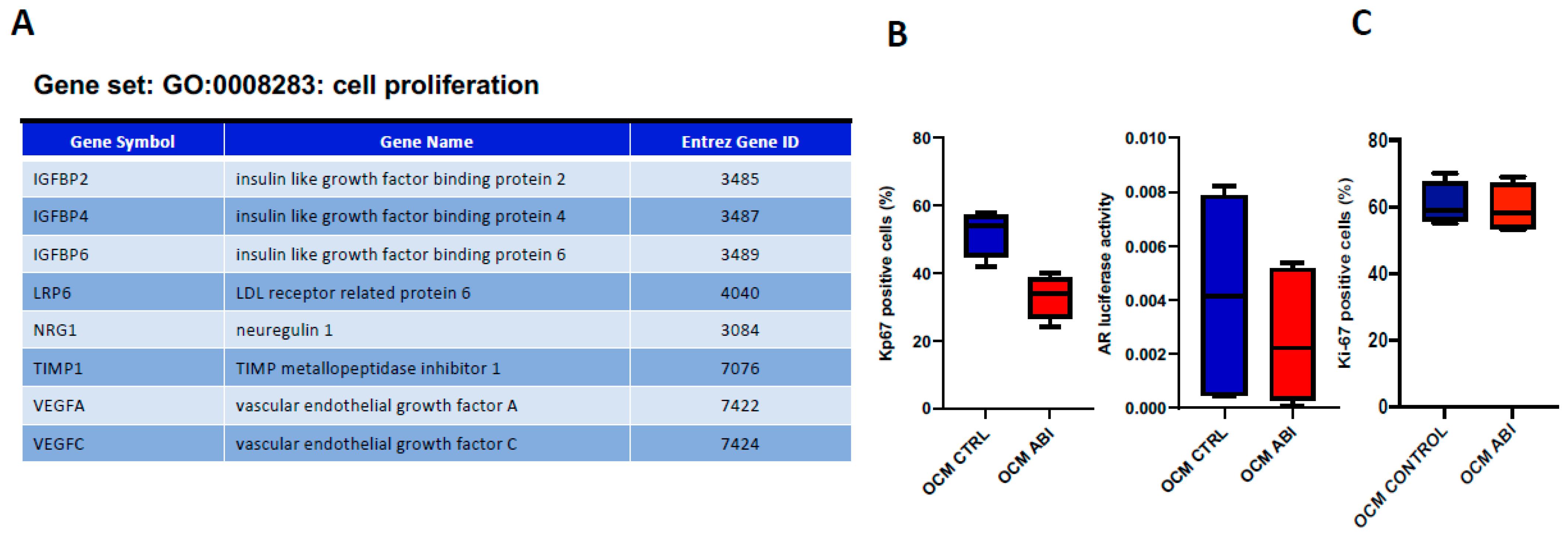
3.2. ABI Exerts an “Indirect” Anti-Tumor Effect Mediated by Osteoblasts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Attard, G.; Belldegrun, A.S.; De Bono, J.S. Selective Blockade of Androgenic Steroid Synthesis by Novel Lyase Inhibitors as a Therapeutic Strategy for Treating Metastatic Prostate Cancer. Br. J. Urol. 2005, 96, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Gartrell, B.A.; Saad, F. Abiraterone in the Management of Castration-Resistant Prostate Cancer Prior to Chemotherapy. Ther. Adv. Urol. 2015, 7, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Fizazi, K.; Scher, H.I.; Molina, A.; Logothetis, C.J.; Chi, K.N.; Jones, R.J.; Staffurth, J.N.; North, S.; Vogelzang, N.J.; Saad, F.; et al. Abiraterone Acetate for Treatment of Metastatic Castration-Resistant Prostate Cancer: Final Overall Survival Analysis of the COU-ABI-301 Randomised, Double-Blind, Placebo-Controlled Phase 3 Study. Lancet Oncol. 2012, 13, 983–992. [Google Scholar] [CrossRef]
- Ryan, C.J.; Smith, M.R.; De Bono, J.S.; Molina, A.; Logothetis, C.J.; De Souza, P.; Fizazi, K.; Mainwaring, P.; Piulats, J.M.; Ng, S.; et al. Abiraterone in Metastatic Prostate Cancer without Previous Chemotherapy. N. Engl. J. Med. 2013, 368, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.J.; Smith, M.R.; Fizazi, K.; Saad, F.; Mulders, P.F.; Sternberg, C.N.; Miller, K.; Logothetis, C.J.; Shore, N.D.; Small, E.J.; et al. Abiraterone Acetate Plus Prednisone Versus Placebo Plus Prednisone in Chemotherapy-Naïve Men with Metastatic Cas-Tration-Resistant Prostate Cancer (COU-ABI-302): Final Overall Survival Analysis of a Randomised, Double-Blind, Place-Bo-Controlled Phase 3 Study. Lancet Oncol. 2015, 16, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Rathkopf, D.E.; Smith, M.R.; de Bono, J.S.; Logothetis, C.J.; Shore, N.D.; de Souza, P.; Fizazi, K.; Mulders, P.F.; Mainwaring, P.; Hainsworth, J.D.; et al. Updated Interim Efficacy Analysis and Long-Term Safety of Abiraterone Acetate in Metastatic Castra-Tion-Resistant Prostate Cancer Patients without Prior Chemotherapy (COU-ABI-302). Eur. Urol. 2014, 66, 815–825. [Google Scholar] [CrossRef]
- Fizazi, K.; Tran, N.; Fein, L.; Matsubara, N.; Rodriguez-Antolin, A.; Alekseev, B.Y.; Özgüroğlu, M.; Ye, D.; Feyerabend, S.; Protheroe, A.; et al. Abiraterone Acetate Plus Prednisone in Patients with Newly Diagnosed High-Risk Metastatic Castra-Tion-Sensitive Prostate Cancer (LATITUDE): Final Overall Survival Analysis of a Randomised, Double-Blind, Phase 3 Trial. Lancet Oncol. 2019, 20, 686–700. [Google Scholar] [CrossRef]
- Fizazi, K.; Tran, N.; Fein, L.; Matsubara, N.; Rodriguez-Antolin, A.; Alekseev, B.Y.; Özgüroğlu, M.; Ye, D.; Feyerabend, S.; Protheroe, A.; et al. Abiraterone plus Prednisone in Metastatic, Castration-Sensitive Prostate Cancer. N. Engl. J. Med. 2017, 377, 352–360. [Google Scholar] [CrossRef]
- Rizzo, S.; Galvano, A.; Pantano, F.; Iuliani, M.; Vincenzi, B.; Passiglia, F.; Spoto, S.; Tonini, G.; Bazan, V.; Russo, A.; et al. The Effects of Enzalutamide and Abiraterone on Skeletal Related Events and Bone Radiological Progression Free Survival in Cas-Tration Resistant Prostate Cancer Patients: An Indirect Comparison of Randomized Controlled Trials. Crit. Rev. Oncol. /Hematol. 2017, 120, 227–233. [Google Scholar] [CrossRef]
- Santini, D.; Cinieri, S.; Gasparro, D.; Bordonaro, R.; Guglielmini, P.F.; Chiuri, V.E.; D’Angelillo, R.M.; Ceresoli, G.L.; Fagnani, D.; Acquati, M.; et al. Effects of Abiraterone Acetate Plus Prednisone on Bone Turnover Markers in Chemotherapy-Naïve Mcrpc Patients After ADT Failure: A Prospective Analysis of the Italian Real-World Study ABITUDE. J. Bone Oncol. 2020, 26, 100341. [Google Scholar] [CrossRef]
- Iuliani, M.; Pantano, F.; Buttigliero, C.; Fioramonti, M.; Bertaglia, V.; Vincenzi, B.; Zoccoli, A.; Ribelli, G.; Tucci, M.; Vignani, F.; et al. Biological and Clinical Effects of Abiraterone on Anti-Resorptive and Anabolic Activity in Bone Microenvironment. Oncotarget 2015, 6, 12520–12528. [Google Scholar] [CrossRef]
- Blaszczyk, N.; Masri, B.A.; Mawji, N.R.; Ueda, T.; McAlinden, G.; Duncan, C.P.; Bruchovsky, N.; Schweikert, H.U.; Schnabel, D.; Jones, E.C.; et al. Osteoblast-Derived Factors Induce Androgen-Independent Proliferation and Expression of Pros-tate-Specific Antigen in Human Prostate Cancer Cells. Clin. Cancer Res. 2004, 10, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- Hagberg Thulin, M.; Nilsson, M.E.; Thulin, P.; Céraline, J.; Ohlsson, C.; Damber, J.E.; Welén, K. Osteoblasts Promote Castra-tion-Resistant Prostate Cancer by Altering Intratumoral Steroidogenesis. Mol. Cell. Endocrinol. 2016, 422, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Bock, N.; Shokoohmand, A.; Kryza, T.; Röhl, J.; Meijer, J.; Tran, P.A.; Nelson, C.C.; Clements, J.A.; Hutmacher, D.W. Engi-neering Osteoblastic Metastases to Delineate the Adaptive Response of Androgen-Deprived Prostate Cancer in the Bone Metastatic Microenvironment. Bone Res. 2019, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.G.; Mathew, P.; Yang, J.; Starbuck, M.W.; Zurita, A.J.; Liu, J.; Sikes, C.; Multani, A.S.; Efstathiou, E.; Lopez, A.; et al. Androgen Receptor-Negative Human Prostate Cancer Cells Induce Osteogenesis in Mice Through FGF9-Mediated Mecha-Nisms. J. Clin. Investig. 2008, 118, 2697–2710. [Google Scholar]
- Ribelli, G.; Simonetti, S.; Iuliani, M.; Rossi, E.; Vincenzi, B.; Tonini, G.; Pantano, F.; Santini, D. Osteoblasts Promote Prostate Cancer Cell Proliferation Through Androgen Receptor Independent Mechanisms. Front. Oncol. 2021, 11, 789885. [Google Scholar] [CrossRef]
- Thalmann, G.N.; Anezinis, P.E.; Chang, S.M.; Zhau, H.E.; Kim, E.E.; Hopwood, V.L.; Pathak, S.; Von Eschenbach, A.C.; Chung, L.W. Androgen-Independent Cancer Progression and Bone Metastasis in the Lncap Model of Human Prostate Cancer. Cancer Res. 1994, 54, 2577–2581. [Google Scholar]
- Iuliani, M.; Simonetti, S.; Ribelli, G.; Napolitano, A.; Longo, U.G.; Vincenzi, B.; Orsaria, P.; Denaro, V.; Tonini, G.; Santini, D.; et al. Biological Effects of Cyclin-Dependent Kinase Inhibitors Ribociclib, Palbociclib and Abemaciclib on Breast Cancer Bone Microenvironment. Int. J. Mol. Sci. 2022, 23, 2477. [Google Scholar] [CrossRef]
- Hensel, J.; Thalmann, G.N. Biology of Bone Metastases in Prostate Cancer. Urology 2016, 92, 6–13. [Google Scholar] [CrossRef]
- Coleman, R.E. Clinical Features of Metastatic Bone Disease and Risk of Skeletal Morbidity. Clin. Cancer Res. 2006, 12, 6243s–6249s. [Google Scholar] [CrossRef]
- Mundy, G. Metastasis to bone: Causes, Consequences and Therapeutic Opportunities. Nat. Rev. Cancer 2002, 2, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, C.J.; Lin, S.-H. Osteoblasts in Prostate Cancer Metastasis to Bone. Nat. Rev. Cancer 2005, 5, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.J.; Edwards, C.M. The Role of the Microenvironment in Prostate Cancer-Associated Bone Disease. Curr. Osteoporos. Rep. 2016, 14, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Ottewell, P.D. The Role of Osteoblasts in Bone Metastasis. J. Bone Oncol. 2016, 5, 124–127. [Google Scholar] [CrossRef]
- Pollak, M. Insulin, Insulin-Like Growth Factors and Neoplasia. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 625–638. [Google Scholar] [CrossRef]
- Holly, J.; Perks, C. The Role of Insulin-Like Growth Factor Binding Proteins. Neuroendocrinology 2006, 83, 154–160. [Google Scholar] [CrossRef]
- Zhu, M.-L.; Kyprianou, N. Androgen Receptor and Growth Factor Signaling Crosstalk in Prostate Cancer Cells. Endocrine-Related Cancer 2008, 15, 841–849. [Google Scholar] [CrossRef]
- Degraff, D.J.; Aguiar, A.A.; Sikes, R.A. Disease Evidence for Igfbp-2 As A Key Player in Prostate Cancer Progression and De-Velopment of Osteosclerotic Lesions. Am. J. Transl. Res. 2009, 1, 115–130. [Google Scholar]
- Uzoh, C.C.; Holly, J.M.P.; Biernacka, K.M.; Persad, R.A.; Bahl, A.; Gillatt, D.; Perks, C.M. Insulin-Like Growth Factor-Binding Protein-2 Promotes Prostate Cancer Cell Growth Via IGF-Dependent or -Independent Mechanisms and Reduces the Efficacy of Docetaxel. Br. J. Cancer 2011, 104, 1587–1593. [Google Scholar] [CrossRef]
- Mehrian-Shai, R.; Chen, C.D.; Shi, T.; Horvath, S.; Nelson, S.F.; Reichardt, J.K.V.; Sawyers, C.L. Insulin Growth Factor-Binding Protein 2 Is A Candidate Biomarker for PTEN Status and PI3K/Akt Pathway Activation in Glioblastoma and Prostate Cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 5563–5568. [Google Scholar] [CrossRef]
- Drivdahl, R.H.; Sprenger, C.; Trimm, K.; Plymate, S.R. Inhibition of Growth and Increased Expression of Insulin-Like Growth Factor-Binding Protein-3 (IGFBP-3) and -6 in Prostate Cancer Cells Stably Transfected with Antisense IGFBP-4 Complementary Deoxyribonucleic Acid*. Endocrinology 2001, 142, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.A. Recent Insights into the Actions of IGFBP-6. J. Cell Commun. Signal. 2015, 9, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Khurana, N.; Sikka, S.C. Interplay Between SOX9, Wnt/β-Catenin and Androgen Receptor Signaling in Castration-Resistant Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Li, Y. Salinomycin Suppresses LRP6 Expression and Inhibits Both Wnt/β-catenin and mTORC1 Signaling in Breast and Prostate Cancer Cells. J. Cell. Biochem. 2014, 115, 1799–1807. [Google Scholar] [CrossRef]
- Zhang, Z.; Karthaus, W.R.; Lee, Y.S.; Gao, V.R.; Wu, C.; Russo, J.W.; Liu, M.; Mota, J.M.; Abida, W.; Linton, E.; et al. Tumor Microenvironment-Derived NRG1 Promotes Antiandrogen Resistance in Prostate Cancer. Cancer Cell 2020, 38, 279–296.e9. [Google Scholar] [CrossRef]
- Yamamura, A.; Nayeem, J.; Muramatsu, H.; Nakamura, K.; Sato, M. MAZ51 Blocks the Tumor Growth of Prostate Cancer by Inhibiting Vascular Endothelial Growth Factor Receptor 3. Front. Pharmacol. 2021, 12, 667474. [Google Scholar] [CrossRef]
- Rinaldo, F.; Li, J.; Wang, E.; Muders, M.; Datta, K. Rala Regulates Vascular Endothelial Growth Factor-C (VEGF-C) Synthesis in Prostate Cancer Cells During Androgen Ablation. Oncogene 2006, 26, 1731–1738. [Google Scholar] [CrossRef]
- Jung, K.-K.; Liu, X.-W.; Chirco, R.; Fridman, R.; Kim, H.-R.C. Identification of CD63 as a Tissue Inhibitor of Metalloproteinase-1 Interacting Cell Surface Protein. EMBO J. 2006, 25, 3934–3942. [Google Scholar] [CrossRef]
- D’Angelo, R.C.; Liu, X.-W.; Najy, A.J.; Jung, Y.S.; Won, J.; Chai, K.X.; Fridman, R.; Kim, H.-R.C. TIMP-1 via TWIST1 Induces EMT Phenotypes in Human Breast Epithelial Cells. Mol. Cancer Res. 2014, 12, 1324–1333. [Google Scholar] [CrossRef]
- Liu, X.-W.; Bernardo, M.M.; Fridman, R.; Kim, H.-R.C. Tissue Inhibitor of Metalloproteinase-1 Protects Human Breast Epithelial Cells Against Intrinsic Apoptotic Cell Death via the Focal Adhesion Kinase/Phosphatidylinositol 3-Kinase and MAPK Signaling Pathway. J. Biol. Chem. 2003, 278, 40364–40372. [Google Scholar] [CrossRef]
- Liu, X.W.; Taube, M.E.; Jung, K.K.; Dong, Z.; Lee, Y.J.; Roshy, S.; Sloane, B.F.; Fridman, R.; Kim, H.R. Tissue Inhibitor of Met-Alloproteinase-1 Protects Human Breast Epithelial Cells from Extrinsic Cell Death: A Potential Oncogenic Activity of Tissue Inhibitor of Metalloproteinase-1. Cancer Res. 2005, 65, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.K.; Vargas, R.; Jacobus, S.; Leitzel, K.; Regan, M.M.; Hamer, P.; Pierce, K.; Brown-Shimer, S.; Carney, W.; Ali, S.M.; et al. Elevated Plasma Tissue Inhibitor of Metalloproteinase-1 Levels Predict Decreased Survival in Castra-Tion-Resistant Prostate Cancer Patients. Cancer 2011, 117, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Pfitzenmaier, J.; Quinn, J.E.; Odman, A.M.; Zhang, J.; Keller, E.T.; Vessella, R.L.; Corey, E. Characterization of C4-2 Prostate Cancer Bone Metastases and Their Response to Castration. J. Bone Miner. Res. 2003, 18, 1882–1888. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iuliani, M.; Simonetti, S.; Ribelli, G.; Cavaliere, S.; Vincenzi, B.; Tonini, G.; Pantano, F.; Santini, D. Osteoblast Secretome Modulated by Abiraterone Treatment Affects Castration Resistant Prostate Cancer Cell Proliferation. Biomedicines 2022, 10, 2154. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10092154
Iuliani M, Simonetti S, Ribelli G, Cavaliere S, Vincenzi B, Tonini G, Pantano F, Santini D. Osteoblast Secretome Modulated by Abiraterone Treatment Affects Castration Resistant Prostate Cancer Cell Proliferation. Biomedicines. 2022; 10(9):2154. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10092154
Chicago/Turabian StyleIuliani, Michele, Sonia Simonetti, Giulia Ribelli, Silvia Cavaliere, Bruno Vincenzi, Giuseppe Tonini, Francesco Pantano, and Daniele Santini. 2022. "Osteoblast Secretome Modulated by Abiraterone Treatment Affects Castration Resistant Prostate Cancer Cell Proliferation" Biomedicines 10, no. 9: 2154. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10092154