
In Vitro Pharmacokinetic Behavior of Antiviral 3-Amidinophenylalanine Derivatives in Rat, Dog and Monkey Hepatocytes
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Primary Hepatocyte Suspensions
2.2. Incubation of Primary Hepatocytes with Inhibitors
2.3. Determination of Unchanged Parent Compound Concentrations
2.4. Assessment of Pharmacokinetics Parameters
2.5. Statistical Analysis
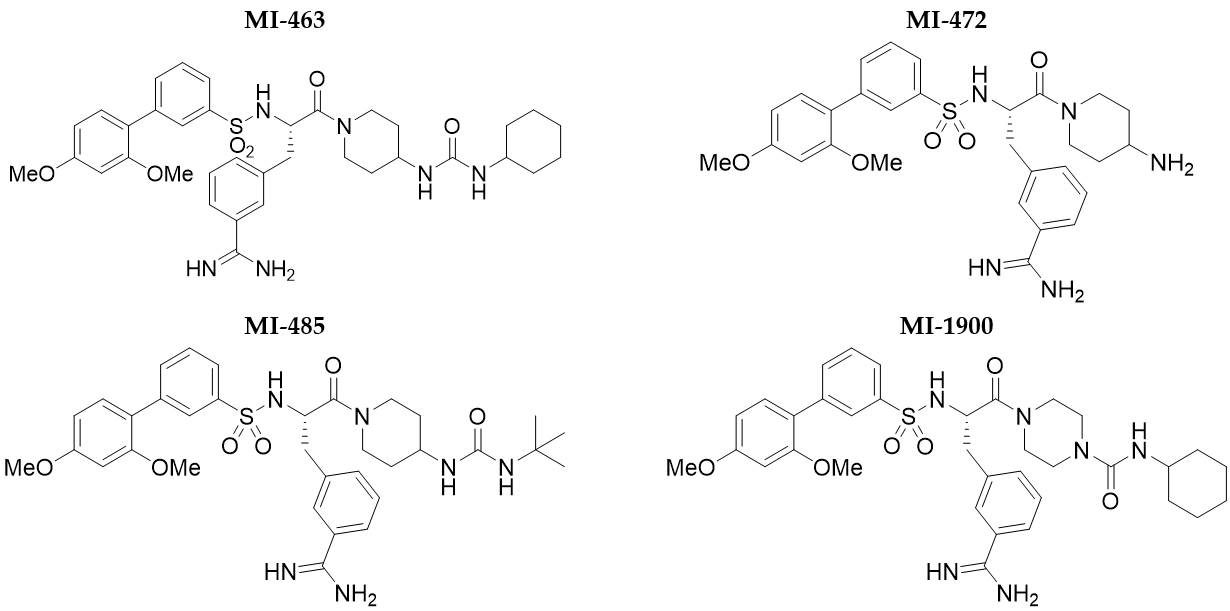
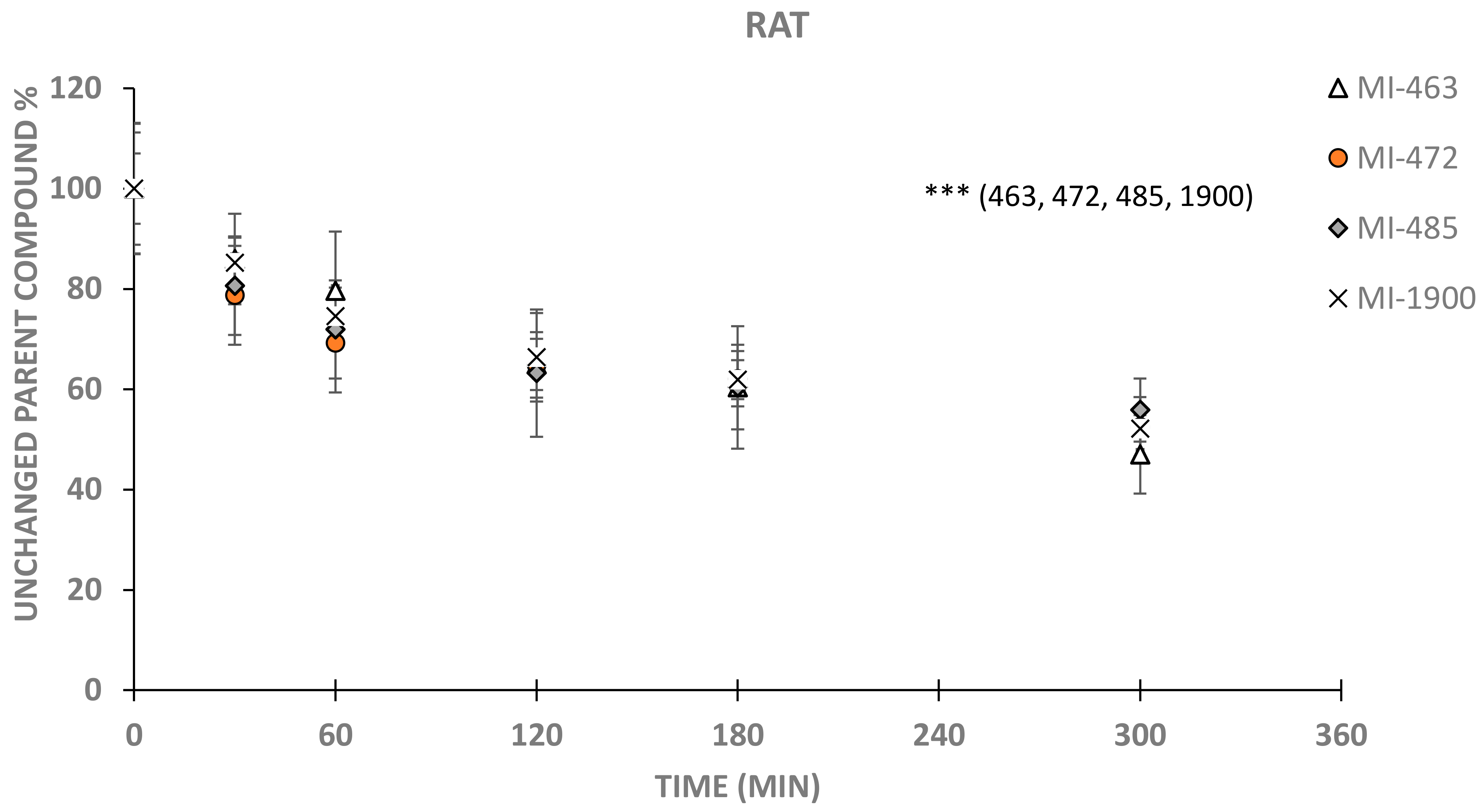
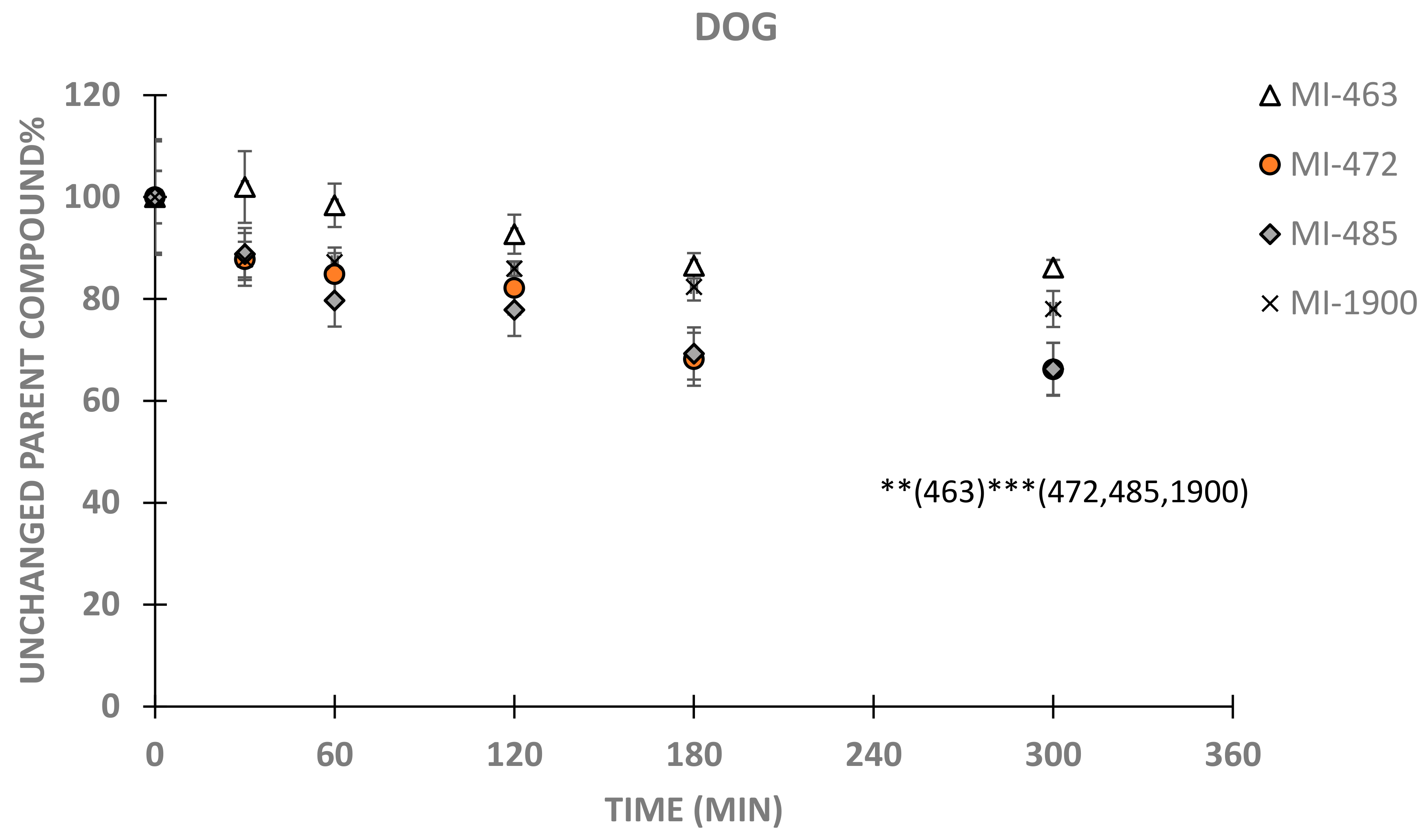
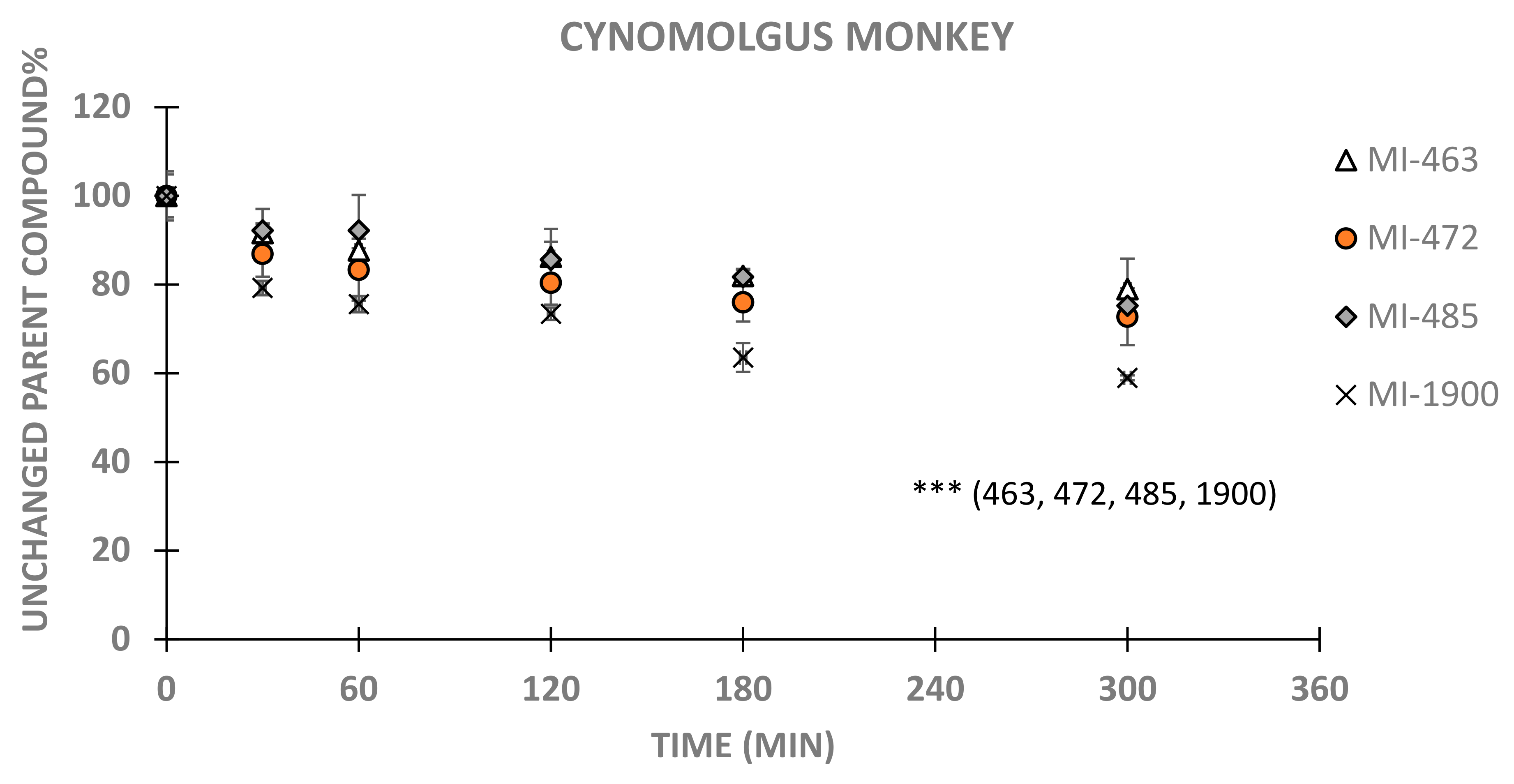
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bugge, T.H.; Antalis, T.M.; Wu, Q. Type II Transmembrane Serine Proteases. J. Biol. Chem. 2009, 284, 23177–23181. [Google Scholar] [CrossRef] [Green Version]
- Förbs, D.; Thiel, S.; Stella, M.C.; Stürzebecher, A.; Schweinitz, A.; Steinmetzer, T.; Stürzebecher, J.; Uhland, K. In vitro inhibition of matriptase prevents invasive growth of cell lines of prostate and colon carcinoma. Int. J. Oncol. 2005, 27, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.M.; Patel, A.; Davidson, R.K.; Swingler, T.E.; Desilets, A.; Young, D.A.; Kelso, E.B.; Donell, S.T.; Cawston, T.E.; Clark, I.M.; et al. Matriptase is a novel initiator of cartilage matrix degradation in osteoarthritis. Arthritis Rheum. 2010, 62, 1955–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krijt, J.; Frýdlová, J.; Gurieva, I.; Přikryl, P.; Báječný, M.; Steinbicker, A.U.; Vokurka, M.; Truksa, J. Matriptase-2 and Hemojuvelin in Hepcidin Regulation: In Vivo Immunoblot Studies in Mask Mice. Int. J. Mol. Sci. 2021, 22, 2650. [Google Scholar] [CrossRef] [PubMed]
- Hatesuer, B.; Bertram, S.; Mehnert, N.; Bahgat, M.M.; Nelson, P.S.; Pöhlman, S.; Schughart, K. Tmprss2 Is Essential for Influenza H1N1 Virus Pathogenesis in Mice. PLoS Pathog. 2013, 9, e1003774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttcher-Friebertshäuser, E. Membrane-Anchored Serine Proteases: Host Cell Factors in Proteolytic Activation of Viral Glycoproteins. Act. Viruses Host Proteases 2018, 16, 153–203. [Google Scholar] [CrossRef]
- Hamilton, B.S.; Gludish, D.W.; Whittaker, G.R. Cleavage Activation of the Human-Adapted Influenza Virus Subtypes by Matriptase Reveals both Subtype and Strain Specificities. J. Virol. 2012, 86, 10579–10586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, J.; Tarnow, C.; Mayoli-Nüssle, D.; Schilling, E.; Meyer, D.; Hammami, M.; Schwalm, F.; Steinmetzer, T.; Guan, Y.; Garten, W.; et al. Matriptase, HAT, and TMPRSS2 Activate the Hemagglutinin of H9N2 Influenza A Viruses. J. Virol. 2013, 87, 1811–1820. [Google Scholar] [CrossRef] [Green Version]
- Kokic, G.; Hillen, H.S.; Tegunov, D.; Dienemann, C.; Seitz, F.; Schmitzova, J.; Farnung, L.; Siewert, A.; Höbartner, C.; Cramer, P. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat. Commun. 2021, 12, 279. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of COVID-19—Preliminary report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 M pro inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [Google Scholar] [CrossRef]
- Wahl, A.; Gralinski, L.E.; Johnson, C.E.; Yao, W.; Kovarova, M.; Dinnon, K.H.; Liu, H.; Madden, V.J.; Krzystek, H.M.; De, C.; et al. SARS-CoV-2 infection is effectively treated and prevented by EIDD-2801. Nature 2021, 591, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Tobback, E.; Degroote, S.; Buysse, S.; Delesie, L.; Van Dooren, L.; Vanherrewege, S.; Barbezange, C.; Hutse, V.; Romano, M.; Thomas, I.; et al. Efficacy and safety of camostat mesylate in early COVID-19 disease in an ambulatory setting: A randomized placebo-controlled phase II trial. Int. J. Infect. Dis. 2022, 122, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Sielaff, F.; Hammami, M.; Böttcher-Friebertshäuser, E.; Garten, W.; Steinmetzer, T. Identification of the first synthetic inhibitors of the type II transmembrane serine protease TMPRSS2 suitable for inhibition of influenza virus activation. Biochem. J. 2013, 452, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilgram, O.; Keils, A.; Benary, G.E.; Müller, J.; Merkl, S.; Ngaha, S.; Huber, S.; Chevillard, F.; Harbig, A.; Magdolen, V.; et al. Improving the selectivity of 3-amidinophenylalanine-derived matriptase inhibitors. Eur. J. Med. Chem. 2022, 238, 114437. [Google Scholar] [CrossRef] [PubMed]
- Colombo, É.; Désilets, A.; Duchêne, D.; Chagnon, F.; Najmanovich, R.; LeDuc, R.; Marsault, E. Design and Synthesis of Potent, Selective Inhibitors of Matriptase. ACS Med. Chem. Lett. 2012, 3, 530–534. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, A.; Gravel, É.; Cloutier, A.; Marois, I.; Colombo, É.; Désilets, A.; Verreault, C.; Leduc, R.; Marsault, É.; Richter, M.V. Matriptase Proteolytically Activates Influenza Virus and Promotes Multicycle Replication in the Human Airway Epithelium. J. Virol. 2013, 87, 4237–4251. [Google Scholar] [CrossRef] [Green Version]
- Shapira, T.; Monreal, I.A.; Dion, S.P.; Buchholz, D.W.; Imbiakha, B.; Olmstead, A.D.; Jager, M.; Désilets, A.; Gao, G.; Martins, M.; et al. A TMPRSS2 inhibitor acts as a pan-SARS-CoV-2 prophylactic and therapeutic. Nature 2022, 605, 340–348. [Google Scholar] [CrossRef]
- Mahdy, M.A.A.; Younis, W.; Ewaida, Z. An Overview of SARS-CoV-2 and Animal Infection. Front. Vet.—Sci. 2020, 7, 596391. [Google Scholar] [CrossRef]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef]
- Johnston, S.C.; Ricks, K.M.; Jay, A.; Raymond, J.L.; Rossi, F.; Zeng, X.; Scruggs, J.; Dyer, D.; Frick, O.; Koehler, J.W.; et al. Development of a coronavirus disease 2019 nonhuman primate model using airborne exposure. PLoS ONE 2021, 16, e0246366. [Google Scholar] [CrossRef]
- Urano, E.; Okamura, T.; Ono, C.; Ueno, S.; Nagata, S.; Kamada, H.; Higuchi, M.; Furukawa, M.; Kamitani, W.; Matsuura, Y.; et al. COVID-19 cynomolgus macaque model reflecting human COVID-19 pathological conditions. Proc. Natl. Acad. Sci. USA 2021, 118, e2104847118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fan, J.; Liu, E. Animal Models for COVID-19 Therapeutic Development: Where We Are and Where We Need to Go. Front. Microbiol. 2022, 13, 907406. [Google Scholar] [CrossRef] [PubMed]
- Hammami, M.; Rühmann, E.; Maurer, E.; Heine, A.; Gütschow, M.; Klebe, G.; Steinmetzer, T. New 3-amidinophenylalanine-derived inhibitors of matriptase. MedChemComm 2012, 3, 807–813. [Google Scholar] [CrossRef]
- List, K.H.; Steinmetzer, T.; Hardes, K. The antiviral potential of host protease inhibitors. In Activation of Viruses by Host Proteases, 1st ed.; Böttcher-Friebertshäuser, E., Garten, W., Klenk, H.D., Eds.; Springer: Marburg, Germany, 2018; pp. 279–325. [Google Scholar]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef]
- Houston, J.B. Utility of in vitro drug metabolism data in predicting in vivo metabolic clearance. Biochem. Pharmacol. 1994, 47, 1469–1479. [Google Scholar] [CrossRef]
- Sohlenius-Sternbeck, A.-K. Determination of the hepatocellularity number for human, dog, rabbit, rat and mouse livers from protein concentration measurements. Toxicol. Vitr. 2006, 20, 1582–1586. [Google Scholar] [CrossRef]
- Davies, B.; Morris, T. Physiological Parameters in Laboratory Animals and Humans. Pharm. Res. 1993, 10, 1093–1095. [Google Scholar] [CrossRef]
- Naritomi, Y.; Terashita, S.; Kagayama, A.; Sugiyama, Y. Utility of Hepatocytes in Predicting Drug Metabolism: Comparison of Hepatic Intrinsic Clearance in Rats and Humans in Vivo and in Vitro. Drug Metab. Dispos. 2003, 31, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Chiba, M.; Sugiyama, Y.; Ishii, Y.; Takahashi, H.; Shibata, Y. Prediction of Hepatic Clearance in Humans from Experimental Animals and In Vitro Data. In Drug Metabolizing Enyzmes. Cytochrome P450 and Other Enzymes in Drug Discovery and Development; Lee, J.S., Obach, R.S., Fisher, M.B., Eds.; Fontis Media: Lausanne, Switzerland, 2003; pp. 453–481. [Google Scholar] [CrossRef]
- Tóth, K.; Sirok, D.; Kiss, Á.; Mayer, A.; Pátfalusi, M.; Hirka, G.; Monostory, K. Utility of in vitro clearance in primary hepatocyte model for prediction of in vivo hepatic clearance of psychopharmacons. Microchem. J. 2018, 136, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Fedor, Z.; Szentkirályi-Tóth, A.; Nagy, G.; Szimrók, Z.; Varga, E.; Pászti, A.; Pászti, Z.; Jerzsele, Á.; Pilgram, O.; Steinmetzer, T.; et al. Interspecies Comparisons of the Effects of Potential Antiviral 3-Amidinophenylalanine Derivatives on Cytochrome P450 1A2 Isoenzyme. Vet.—Sci. 2022, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Pászti-Gere, E.; Szentkirályi, A.; Fedor, Z.; Nagy, G.; Szimrók, Z.; Pászti, Z.; Pászti, A.; Pilgram, O.; Steinmetzer, T.; Bodnárová, S.; et al. In vitro interaction of potential antiviral TMPRSS2 inhibitors with human serum albumin and cytochrome P 450 isoenzymes. Biomed. Pharmacother. 2022, 146, 112513. [Google Scholar] [CrossRef]
- Gerets, H.H.J.; Tilmant, K.; Gerin, B.; Chanteux, H.; Depelchin, B.O.; Dhalluin, S.; Atienzar, F.A. Characterization of primary human hepatocytes, HepG2 cells, and HepaRG cells at the mRNA level and CYP activity in response to inducers and their predictivity for the detection of human hepatotoxins. Cell Biol. Toxicol. 2012, 28, 69–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkening, S.; Stahl, F.; Bader, A. Comparison of primary human hepatocytes and hepatoma cell line hepg2 with regard to their biotransformation properties. Drug Metab. Dispos. 2003, 31, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.; Donzelli, M.; Maseneni, S.; Boess, F.; Roth, A.; Krähenbühl, S.; Haschke, M. Comparison of Liver Cell Models Using the Basel Phenotyping Cocktail. Front. Pharmacol. 2016, 7, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Wu, Y.; Rui, X.; Yang, Y.; Ling, C.; Liu, S.; Liu, S.; Wang, Y. Animal models for COVID-19: Advances, gaps and perspectives. Signal Transduct. Target. Ther. 2022, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Caldera-Crespo, L.A.; Paidas, M.J.; Roy, S.; Schulman, C.I.; Kenyon, N.S.; Daunert, S.; Jayakumar, A.R. Experimental Models of COVID-19. Front. Cell Infect. Microbiol. 2022, 11, 792584. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.; Lowen, A.C. Animal models for SARS-CoV-2. Curr. Opin. Virol. 2021, 48, 73–81. [Google Scholar] [CrossRef]
- Choudhary, S.; Kanevsky, I.; Tomlinson, L. Animal models for studying COVID-19, prevention, and therapy: Pathology and disease phenotypes. Vet.—Pathol. 2022, 59, 516–527. [Google Scholar] [CrossRef]
- Rosa, R.B.; Dantas, W.M.; Do Nascimento, J.C.F.; Da Silva, M.V.; De Oliveira, R.N.; Pena, L.J. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses 2021, 13, 379. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Uno, Y. Cynomolgus monkey CYPs: A comparison with human CYPs. Xenobiotica 2009, 39, 578–581. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, T.; Ufuk, A.; Cantrill, C.; Kosa, R.E.; Bi, Y.-A.; Niosi, M.; Modi, S.; Rodrigues, A.D.; Tremaine, L.M.; Varma, M.V.S.; et al. Predicting Human Clearance of Organic Anion Transporting Polypeptide Substrates Using Cynomolgus Monkey: In Vitro–In Vivo Scaling of Hepatic Uptake Clearance. Drug Metab. Dispos. 2018, 46, 989–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Yang, Z.; Rodrigues, A.D. Cynomolgus Monkey as an Emerging Animal Model to Study Drug Transporters: In Vitro, In Vivo, In Vitro-to-In Vivo Translation. Drug Metab. Dispos. 2022, 50, 299–319. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Rat | Dog | Monkey |
|---|---|---|---|
| Number of hepatocytes in liver (×106 cells/g liver) | 117 | 215 | 114 |
| Liver weight (g) | 10 | 320 | 150 |
| Body weight (kg) | 0.25 | 10 | 5 |
| Liver blood flow (mL/min/kg) | 55.2 | 30.9 | 43.6 |
| Plasma/blood ratio | 0.58 | 0.57 | 0.61 |
| Clint (mL/min/kg) | Predicted Cl CLH | Extraction Ratio EH | F (%) | Classification | |
|---|---|---|---|---|---|
| MI-463 | |||||
| Rat | 31.87 | 15.97 | 0.289 | 71.07 | Low-extraction drug |
| Dog | 19.487 | 9.25 | 0.299 | 70.06 | Low-extraction drug |
| Macaque | 6.58 | 5.27 | 0.121 | 87.91 | Low-extraction drug |
| MI-472 | |||||
| Rat | 35.58 | 16.85 | 0.305 | 69.47 | Intermediate-extraction drug |
| Dog | 26.69 | 10.61 | 0.343 | 65.66 | Intermediate-extraction drug |
| Macaque | 12.00 | 8.27 | 0.190 | 87.91 | Low-extraction drug |
| MI-485 | |||||
| Rat | 37.78 | 17.33 | 0.314 | 68.60 | Intermediate-extraction drug |
| Dog | 22.75 | 9.93 | 0.321 | 67.87 | Intermediate-extraction drug |
| Macaque | 11.40 | 7.98 | 0.183 | 81.69 | Low-extraction drug |
| MI-1900 | |||||
| Rat | 33.76 | 16.43 | 0.298 | 70.23 | Low-extraction drug |
| Dog | 10.46 | 6.56 | 0.212 | 78.76 | Low-extraction drug |
| Macaque | 16.10 | 10.03 | 0.230 | 76.99 | Low-extraction drug |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lányi, K.; Monostory, K.; Steinmetzer, T.; Jerzsele, Á.; Pászti-Gere, E. In Vitro Pharmacokinetic Behavior of Antiviral 3-Amidinophenylalanine Derivatives in Rat, Dog and Monkey Hepatocytes. Biomedicines 2023, 11, 682. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030682
Lányi K, Monostory K, Steinmetzer T, Jerzsele Á, Pászti-Gere E. In Vitro Pharmacokinetic Behavior of Antiviral 3-Amidinophenylalanine Derivatives in Rat, Dog and Monkey Hepatocytes. Biomedicines. 2023; 11(3):682. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030682
Chicago/Turabian StyleLányi, Katalin, Katalin Monostory, Torsten Steinmetzer, Ákos Jerzsele, and Erzsébet Pászti-Gere. 2023. "In Vitro Pharmacokinetic Behavior of Antiviral 3-Amidinophenylalanine Derivatives in Rat, Dog and Monkey Hepatocytes" Biomedicines 11, no. 3: 682. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030682