
Novel Potential Markers of Myofibroblast Differentiation Revealed by Single-Cell RNA Sequencing Analysis of Mesenchymal Stromal Cells in Profibrotic and Adipogenic Conditions

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human MSC and HDF Isolation and Culture
2.2. Single-Cell Droplet-Based RNA-Seq Library Preparation and Sequencing
2.3. Analysis and Quality Control of Single-Cell RNA-Seq Data
2.4. cDNA Libraries Quality Analysis
2.5. Library Preparation, Sequencing, and Alignment
2.6. RNA-Seq Data Bioinformatic Analysis
2.7. Isolation of Total RNA
2.8. Real-Time PCR with Reverse Transcription
2.9. Immunocytochemistry
2.10. Statistical Analysis
3. Results
3.1. Cultured MSCs Respond Differently to Profibrotic Stimuli
3.2. Identification of Genes That Change Expression in MSCs in Opposite Directions upon Stimulation of Myofibroblast and Adipogenic Differentiation
3.3. Gene Expression Analysis in MSCs by RT-PCR
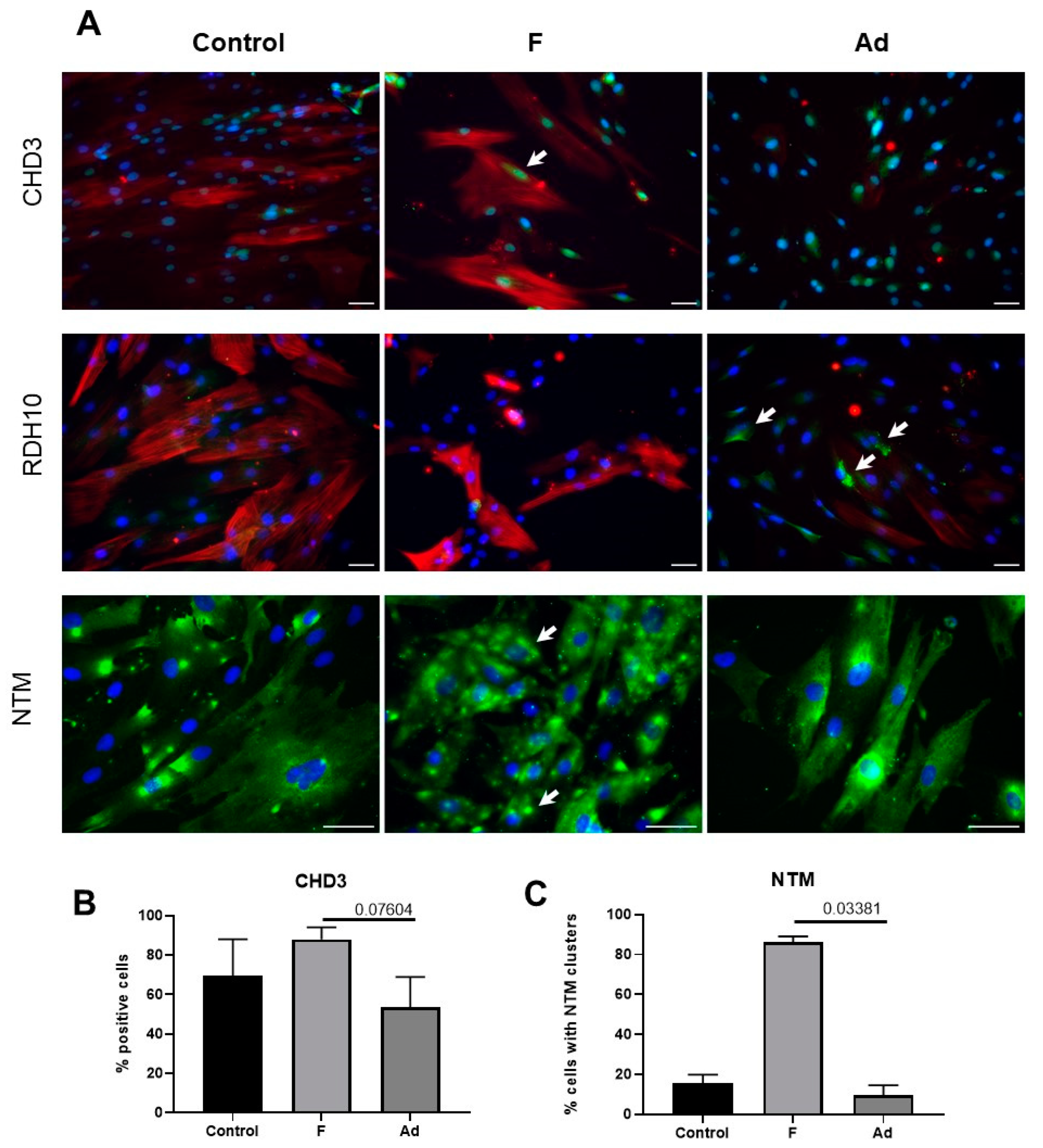
3.4. Analysis of Protein Markers in MSCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gabbiani, G. The Biology of the Myofibroblast. Kidney Int. 1992, 41, 530–532. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B.; Dugina, V.; Ballestrem, C.; Wehrle-Haller, B.; Chaponnier, C. α-Smooth Muscle Actin Is Crucial for Focal Adhesion Maturation in Myofibroblasts. MBoC 2003, 14, 2508–2519. [Google Scholar] [CrossRef] [Green Version]
- Evanko, S.P.; Potter-Perigo, S.; Petty, L.J.; Workman, G.A.; Wight, T.N. Hyaluronan Controls the Deposition of Fibronectin and Collagen and Modulates TGF-Β1 Induction of Lung Myofibroblasts. Matrix Biol. 2015, 42, 74–92. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Tamaki, Z.; Wang, W.; Hinchcliff, M.; Hoover, P.; Getsios, S.; White, E.S.; Varga, J. Fibronectin EDA Promotes Chronic Cutaneous Fibrosis Through Toll-Like Receptor Signaling. Sci. Transl. Med. 2014, 6, 232ra50. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, C.; McDonald, J.A. The Roles of the Myofibroblast in Idiopathic Pulmonary Fibrosis. Ultrastructural and Immunohistochemical Features of Sites of Active Extracellular Matrix Synthesis. Am. J. Pathol. 1991, 138, 1257–1265. [Google Scholar]
- Shinde, A.V.; Kelsh, R.; Peters, J.H.; Sekiguchi, K.; Van De Water, L.; McKeown-Longo, P.J. The A4β1 Integrin and the EDA Domain of Fibronectin Regulate a Profibrotic Phenotype in Dermal Fibroblasts. Matrix Biol. 2015, 41, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Kanisicak, O.; Khalil, H.; Ivey, M.J.; Karch, J.; Maliken, B.D.; Correll, R.N.; Brody, M.J.; Lin, S.-C.J.; Aronow, B.J.; Tallquist, M.D.; et al. Genetic Lineage Tracing Defines Myofibroblast Origin and Function in the Injured Heart. Nat. Commun. 2016, 7, 12260. [Google Scholar] [CrossRef] [Green Version]
- Abu El-Asrar, A.M.; Missotten, L.; Geboes, K. Expression of Myofibroblast Activation Molecules in Proliferative Vitreoretinopathy Epiretinal Membranes. Acta Ophthalmol. 2011, 89, e115–e121. [Google Scholar] [CrossRef]
- Lemos, D.R.; Duffield, J.S. Tissue-Resident Mesenchymal Stromal Cells: Implications for Tissue-Specific Antifibrotic Therapies. Sci. Transl. Med. 2018, 10, eaan5174. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast Precursors in Normal and Irradiated Mouse Hematopoietic Organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Kalinina, N.; Kharlampieva, D.; Loguinova, M.; Butenko, I.; Pobeguts, O.; Efimenko, A.; Ageeva, L.; Sharonov, G.; Ischenko, D.; Alekseev, D.; et al. Characterization of Secretomes Provides Evidence for Adipose-Derived Mesenchymal Stromal Cells Subtypes. Stem Cell Res. Ther. 2015, 6, 221. [Google Scholar] [CrossRef] [Green Version]
- Harrell, C.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned Medium from Human Mesenchymal Stromal Cells: Towards the Clinical Translation. IJMS 2019, 20, 1656. [Google Scholar] [CrossRef] [Green Version]
- Basalova, N.; Sagaradze, G.; Arbatskiy, M.; Evtushenko, E.; Kulebyakin, K.; Grigorieva, O.; Akopyan, Z.; Kalinina, N.; Efimenko, A. Secretome of Mesenchymal Stromal Cells Prevents Myofibroblasts Differentiation by Transferring Fibrosis-Associated MicroRNAs within Extracellular Vesicles. Cells 2020, 9, 1272. [Google Scholar] [CrossRef]
- Dinh, P.-U.C.; Paudel, D.; Brochu, H.; Popowski, K.D.; Gracieux, M.C.; Cores, J.; Huang, K.; Hensley, M.T.; Harrell, E.; Vandergriff, A.C.; et al. Inhalation of Lung Spheroid Cell Secretome and Exosomes Promotes Lung Repair in Pulmonary Fibrosis. Nat. Commun. 2020, 11, 1064. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, N.; Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Nassiri, S.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Prevent and Revert Experimental Pulmonary Fibrosis through Modulation of Monocyte Phenotypes. JCI Insight 2019, 4, e128060. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Novoseletskaya, E.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Eremichev, R.; Milovskaya, I.; Kulebyakin, K.; Kulebyakina, M.; Rodionov, S.; Omelyanenko, N.; et al. Mesenchymal Stromal Cell-Produced Components of Extracellular Matrix Potentiate Multipotent Stem Cell Response to Differentiation Stimuli. Front. Cell Dev. Biol. 2020, 8, 555378. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated Analysis of Multimodal Single-Cell Data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef]
- Sun, C.; Wang, L.; Wang, H.; Huang, T.; Yao, W.; Li, J.; Zhang, X. Single-Cell RNA-Seq Highlights Heterogenei-ty in Human Primary Wharton’s Jelly Mesenchymal Stem/Stromal Cells Cultured in Vitro. Stem Cell Res. Ther. 2020, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, X.; Yang, J.; Gong, Y.; Zhang, H.; Qiu, X.; Liu, Y.; Zhou, C.; Chen, Y.; Greenbaum, J.; et al. Sin-gle-Cell RNA Sequencing Deconvolutes the in Vivo Heterogeneity of Human Bone Marrow-Derived Mesen-chymal Stem Cells. Int. J. Biol. Sci. 2021, 17, 4192–4206. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; Theret, M.; Scott, W.; Hill, L.; Underhill, T.M.; Hinz, B.; Rossi, F.M.V. Multipotent Stromal Cells: One Name, Multiple Identities. Cell Stem Cell 2021, 28, 1690–1707. [Google Scholar] [CrossRef] [PubMed]
- Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.E. TGF Beta −1, −2 and −3 in the Modulation of Fibrosis in the Cornea and Other Organs. Exp. Eye Res. 2021, 207, 108594. [Google Scholar] [CrossRef]
- Hinz, B. Formation and Function of the Myofibroblast during Tissue Repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Pakshir, P.; Hinz, B. The Big Five in Fibrosis: Macrophages, Myofibroblasts, Matrix, Mechanics, and Miscom-munication. Matrix Biol. 2018, 68–69, 81–93. [Google Scholar] [CrossRef]
- Merkt, W.; Zhou, Y.; Han, H.; Lagares, D. Myofibroblast Fate Plasticity in Tissue Repair and Fibrosis: Deactiva-tion, Apoptosis, Senescence and Reprogramming. Wound Repair Regen. 2021, 29, 678–691. [Google Scholar] [CrossRef]
- Hinz, B.; Lagares, D. Evasion of Apoptosis by Myofibroblasts: A Hallmark of Fibrotic Diseases. Nat. Rev. Rheu-Matol. 2020, 16, 11–31. [Google Scholar] [CrossRef]
- Hinz, B.; McCulloch, C.A.; Coelho, N.M. Mechanical Regulation of Myofibroblast Phenoconversion and Colla-gen Contraction. Exp. Cell Res. 2019, 379, 119–128. [Google Scholar] [CrossRef]
- Grigorieva, O.A.; Vigovskiy, M.A.; Dyachkova, U.D.; Basalova, N.A.; Aleksandrushkina, N.A.; Kulebyakina, M.A.; Zaitsev, I.L.; Popov, V.S.; Efimenko, A.Y. Mechanisms of Endothelial-to-Mesenchymal Transition In-duction by Extracellular Matrix Components in Pulmonary Fibrosis. Bull. Exp. Biol. Med. 2021, 171, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.-N. Interactions between TGF-Β1, Canonical WNT/β-Catenin Pathway and PPAR γ in Radiation-Induced Fibrosis. Oncotarget 2017, 8, 90579–90604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, N.U.B.; Willumsen, N.; Sand, J.M.B.; Larsen, L.; Karsdal, M.A.; Leeming, D.J. Type VIII Collagen Is Elevated in Diseases Associated with Angiogenesis and Vascular Remodeling. Clin. Biochem. 2016, 49, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Shuttleworth, C.A. Type VIII Collagen. Int. J. Biochem. Cell Biol. 1997, 29, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.U.B.; Gudmann, N.S.; Karsdal, M.A. Type VIII Collagen. In Biochemistry of Collagens, Laminins and Elastin; Elsevier: Amsterdam, The Netherlands, 2019; pp. 75–81. ISBN 978-0-12-817068-7. [Google Scholar]
- Skrbic, B.; Engebretsen, K.V.T.; Strand, M.E.; Lunde, I.G.; Herum, K.M.; Marstein, H.S.; Sjaastad, I.; Lunde, P.K.; Carlson, C.R.; Christensen, G.; et al. Lack of Collagen VIII Reduces Fibrosis and Promotes Early Mortality and Cardiac Dilatation in Pressure Overload in Mice. Cardiovasc. Res. 2015, 106, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Szabo-Rogers, H.L. Chondrocyte Polarity During Endochondral Ossification Requires Protein–Protein Interactions Between Prickle1 and Dishevelled2/3. J. Bone Miner. Res. 2021, 36, 2399–2412. [Google Scholar] [CrossRef]
- Wan, Y.; Lantz, B.; Cusack, B.J.; Szabo-Rogers, H.L. Prickle1 Regulates Differentiation of Frontal Bone Osteoblasts. Sci. Rep. 2018, 8, 18021. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Cao, N.; Miao, Y.; Dai, Y.; Wei, Z. Morin Acts as a USP7 Inhibitor to Hold Back the Migration of Rheumatoid Arthritis Fibroblast-Like Synoviocytes in a “Prickle1-mTORC2” Dependent Manner. Mol. Nutr. Food Res. 2021, 65, 2100367. [Google Scholar] [CrossRef]
- Lim, B.C.; Matsumoto, S.; Yamamoto, H.; Mizuno, H.; Kikuta, J.; Ishii, M.; Kikuchi, A. Prickle1 Promotes Focal Adhesion Disassembly in Cooperation with CLASP-LL5β Complex in Migrating Cells. J. Cell Sci. 2016, 129, jcs.185439. [Google Scholar] [CrossRef] [Green Version]
- Daulat, A.M.; Bertucci, F.; Audebert, S.; Sergé, A.; Finetti, P.; Josselin, E.; Castellano, R.; Birnbaum, D.; Angers, S.; Borg, J.-P. PRICKLE1 Contributes to Cancer Cell Dissemination through Its Interaction with MTORC2. Dev. Cell 2016, 37, 311–325. [Google Scholar] [CrossRef]
- Andrews, G.K. The Transcription Factors MTF-1 and USF1 Cooperate to Regulate Mouse Metallothionein-I Expression in Response to the Essential Metal Zinc in Visceral Endoderm Cells during Early Development. EMBO J. 2001, 20, 1114–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.H.; Sul, H.S. Insulin Signaling in Fatty Acid and Fat Synthesis: A Transcriptional Perspective. Curr. Opin. Pharmacol. 2010, 10, 684–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodarzi, A.A.; Kurka, T.; Jeggo, P.A. KAP-1 Phosphorylation Regulates CHD3 Nucleosome Remodeling during the DNA Double-Strand Break Response. Nat. Struct. Mol. Biol. 2011, 18, 831–839. [Google Scholar] [CrossRef]
- Goodman, J.V.; Bonni, A. Regulation of Neuronal Connectivity in the Mammalian Brain by Chromatin Remodeling. Curr. Opin. Neurobiol. 2019, 59, 59–68. [Google Scholar] [CrossRef]
- Gil, O.D.; Zanazzi, G.; Struyk, A.F.; Salzer, J.L. Neurotrimin Mediates Bifunctional Effects on Neurite Outgrowth via Homophilic and Heterophilic Interactions. J. Neurosci. 1998, 18, 9312–9325. [Google Scholar] [CrossRef] [Green Version]
- Crigler, L.; Robey, R.C.; Asawachaicharn, A.; Gaupp, D.; Phinney, D.G. Human Mesenchymal Stem Cell Subpopulations Express a Variety of Neuro-Regulatory Molecules and Promote Neuronal Cell Survival and Neuritogenesis. Exp. Neurol. 2006, 198, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Geyer, P.E.; Gupta, R.; Santos, A.; Meier, F.; Doll, S.; Wewer Albrechtsen, N.J.; Klein, S.; Ortiz, C.; Uschner, F.E.; et al. Dynamic Human Liver Proteome Atlas Reveals Functional Insights into Disease Pathways. Mol. Syst. Biol. 2022, 18, e10947. [Google Scholar] [CrossRef] [PubMed]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. MicroRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Sheu, C.; Chang, W.; Tsai, M.; Liao, S.; Chong, I.; Kuo, P. Bioinformatic Analysis of Next-generation Sequencing Data to Identify Dysregulated Genes in Fibroblasts of Idiopathic Pulmonary Fibrosis. Int. J. Mol. Med. 2019, 43, 1643–1656. [Google Scholar] [CrossRef]
- Defnet, A.E.; Shah, S.D.; Huang, W.; Shapiro, P.; Deshpande, D.A.; Kane, M.A. Dysregulated Retinoic Acid Signaling in Airway Smooth Muscle Cells in Asthma. FASEB J. 2021, 35, e22016. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | Forward | Reverse |
|---|---|---|---|
| 36B4 | Acidic ribosomal phosphoprotein P0 | 5′-GCTGCTGCCCGTGCTGGTG-3′ | 5′-TGGTGCCCCTGGAGATTTTAGTGG-3′ |
| ADIPOQ | Adiponectin | 5′-GACCAGGAAACCACGACTCA-3′ | 5′-TTTCACCGATGTCTCCCTTAGG-3′ |
| PPARG | Peroxisome proliferator-activated receptors | 5′-TCAGGTTTGGGCGGATGC-3′ | 5′-TCAGCGGGAAGGACTTTATGTATG-3′ |
| CEBPA | CCAAT—enhancer-binding protein alpha | 5′-TATAGGCTGGGCTTCCCCTT-3′ | 5′-AGCTTTCTGGTGTGACTCGG-3′ |
| FABP4 | Fatty acid binding protein 4 | 5′-ACTGGGCCAGGAATTTGACG-3′ | 5′-CTCGTGGAAGTGACGCCTT-3′ |
| CHD3 | Chromodomain-helicase-DNA-binding protein 3 | 5′-CCGTCAGCATTGGGTGTGAA-3′ | 5′-TCTTGCGTTTTCGGGGTTTTC-3′ |
| NTM | Neurotrimin | 5′-CCAAAGACCTCTAGGGTCCAC-3′ | 5′-GTCTCCAAGTAACCGTAGGCT-3′ |
| RDH10 | Retinol dehydrogenase 10 | 5′-ACCTGACGGCTGAAAGAGTC-3′ | 5′-GAAAAGCCTTAGTGGTCCAGAAG-3′ |
| FN1 | Fibronectin-1 | 5′-CGGTGGCTGTCAGTCAAAG-3′ | 5′-AAACCTCGGCTTCCTCCATAA-3′ |
| VCAN | Versican | 5′-GTAACCCATGCGCTACATAAAGT-3′ | 5′-GGCAAAGTAGGCATCGTTGAAA-3′ |
| COL8A1 | Collagen Type VIII Alpha 1 Chain | 5′-GGGAGTGCTGCTTACCATTTC-3′ | 5′-AGCGGCTTGATCCCATAGTAG-3′ |
| PLN2 | Perilipin-2 | 5′-ATGGCATCCGTTGCAGTTGAT-3′ | 5′-GGACATGAGGTCATACGTGGAG-3′ |
| SERPINE2 | Serpine 2 | 5′-TGGTGATGAGATACGGCGTAA-3′ | 5′-GTTAGCCACTGTCACAATGTCTT-3′ |
| TPN2 | Tropomyosin 2 | 5′-AGACCCGAGCAGAGTTTGC-3′ | 5′-TGGTGAATCTCGACGTTCTCC-3′ |
| AVEN | Caspase and apoptosis activator inhibitor | 5′-GCGCCGGTTGAAGATGACA-3′ | 5′-TGCAGAGCTAAGGAGGACACT-3′ |
| CLIP1 | CAP-Gly domain-containing binding protein 1 | 5′-AGGAAGGTGCAAGCAGAAGAT-3′ | 5′-GTTTTTGTAAGGTTGCTGATCGG-3′ |
| GARS | Glycyl-tRNA synthetase 1 | 5′-ATGGAGGTGTTAGTGGTCTGT-3′ | 5′-CTGTTCCTCTTGGATAAAGTGCT-3′ |
| LIMS2 | LIM zinc finger domain containing protein 2 | 5′-GCACCGGCACTATGAGAAGAA-3′ | 5′-ACGGGCTTCATGTCGAACTC-3′ |
| MINDY2 | MINDY lysine 48 deubiquitinase 2 | 5′-TTGCACAAACTACAGACAGGC-3′ | 5′-TGAGGGTCTACTAACCACCCA-3′ |
| MT1X | Metallothionein 1× | 5′-AACTCCTGCTTCTCCTTGCC-3′ | 5′-GCTCTATTTACATCTGAGAGCACAA-3′ |
| MYO1B | Myosin 1b | 5′-CGGATGAAGCATACAGATCCC-3′ | 5′-CTGCCACATAGGACATGACAAG-3′ |
| PDLIM1 | PDZ and LIM domain 1 | 5′-GCTGGCCTCTACTCTTCTGAA-3′ | 5′-GCTGAGCATGGTCTAAGGGT-3′ |
| PRICKLE1 | Prickle planar cell polarity protein 1 | 5′-GCTGCCTTGAGTGTGAAACG-3′ | 5′-TGCCCGTCATAGGTCATCTGT-3′ |
| SLIT3 | Slit- guidance ligand 3 | 5′-GGCATCGTCGAAATACGCCTA-3′ | 5′-GCTGATGTCTATTCGCTTCAGTT-3′ |
| UCHL1 | Ubiquitin carboxy-terminal hydrolase L1 | 5′-AATGTCGGGTAGATGACAAGGT-3′ | 5′-GGCATTCGTCCATCAAGTTCATA-3′ |
| SOD2 | Superoxide dismutase 2 | 5′-TTTCAATAAGGAACGGGGACAC-3′ | 5′-GTGCTCCCACACATCAATCC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigorieva, O.; Basalova, N.; Vigovskiy, M.; Arbatskiy, M.; Dyachkova, U.; Kulebyakina, M.; Kulebyakin, K.; Tyurin-Kuzmin, P.; Kalinina, N.; Efimenko, A. Novel Potential Markers of Myofibroblast Differentiation Revealed by Single-Cell RNA Sequencing Analysis of Mesenchymal Stromal Cells in Profibrotic and Adipogenic Conditions. Biomedicines 2023, 11, 840. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030840
Grigorieva O, Basalova N, Vigovskiy M, Arbatskiy M, Dyachkova U, Kulebyakina M, Kulebyakin K, Tyurin-Kuzmin P, Kalinina N, Efimenko A. Novel Potential Markers of Myofibroblast Differentiation Revealed by Single-Cell RNA Sequencing Analysis of Mesenchymal Stromal Cells in Profibrotic and Adipogenic Conditions. Biomedicines. 2023; 11(3):840. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030840
Chicago/Turabian StyleGrigorieva, Olga, Nataliya Basalova, Maksim Vigovskiy, Mikhail Arbatskiy, Uliana Dyachkova, Maria Kulebyakina, Konstantin Kulebyakin, Pyotr Tyurin-Kuzmin, Natalia Kalinina, and Anastasia Efimenko. 2023. "Novel Potential Markers of Myofibroblast Differentiation Revealed by Single-Cell RNA Sequencing Analysis of Mesenchymal Stromal Cells in Profibrotic and Adipogenic Conditions" Biomedicines 11, no. 3: 840. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11030840