
Association of TP53 Single Nucleotide Polymorphisms with Prostate Cancer in a Racially Diverse Cohort of Men

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. SNP Analysis and Validation
2.3. Statistical Analysis
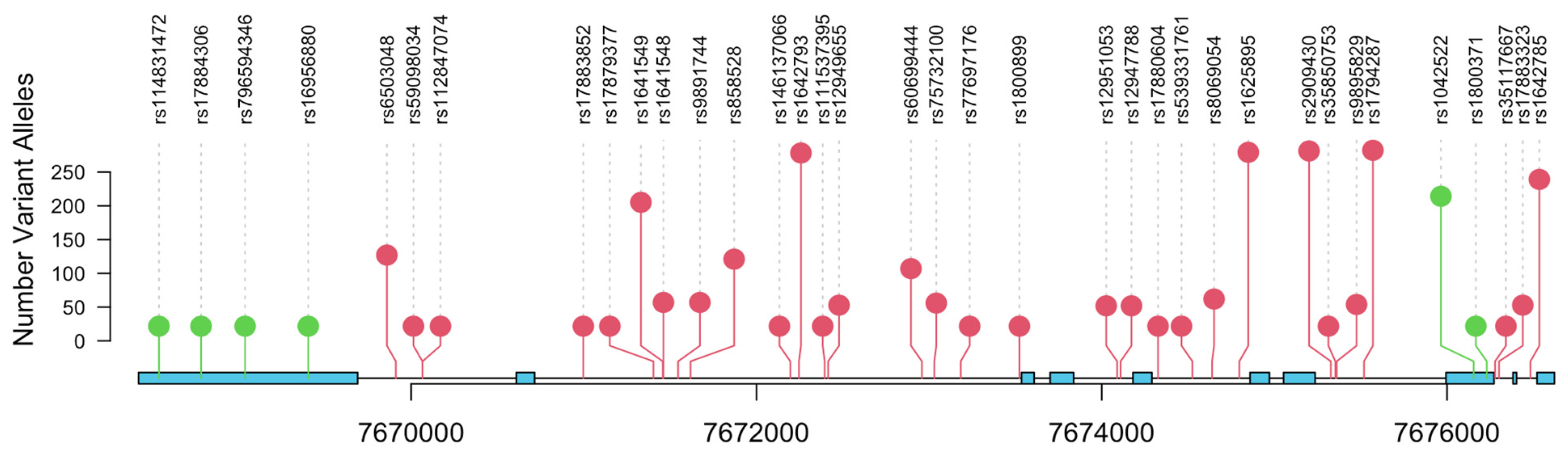
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- McHugh, J.; Saunders, E.J.; Dadaev, T.; McGrowder, E.; Bancroft, E.; Kote-Jarai, Z.; Eeles, R. Prostate cancer risk in men of differing genetic ancestry and approaches to disease screening and management in these groups. Br. J. Cancer 2022, 126, 1366–1373. [Google Scholar] [CrossRef]
- Kohaar, I.; Petrovics, G.; Srivastava, S. A Rich Array of Prostate Cancer Molecular Biomarkers: Opportunities and Challenges. Int. J. Mol. Sci. 2019, 20, 1813. [Google Scholar] [CrossRef] [PubMed]
- Conti, D.V.; Darst, B.F.; Moss, L.C.; Saunders, E.J.; Sheng, X.; Chou, A.; Schumacher, F.R.; Olama, A.A.A.; Benlloch, S.; Dadaev, T.; et al. Trans-ancestry genome-wide association meta-analysis of prostate cancer identifies new susceptibility loci and informs genetic risk prediction. Nat. Genet. 2021, 53, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.P. Cancer. p53, guardian of the genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 10. [Google Scholar] [CrossRef]
- Brosh, R.; Rotter, V. When mutants gain new powers: News from the mutant p53 field. Nat. Rev. Cancer 2009, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, F.P.; Fraumeni, J.F.; Mulvihill, J.J.; Blattner, W.A.; Dreyfus, M.G.; Tucker, M.A.; Miller, R.W. A cancer family syndrome in twenty-four kindreds. Cancer Res. 1988, 48, 5358–5362. [Google Scholar] [PubMed]
- Malkin, D.; Li, F.P.; Strong, L.C.; Fraumeni, J.F., Jr.; Nelson, C.E.; Kim, D.H.; Kassel, J.; Gryka, M.A.; Bischoff, F.Z.; Tainsky, M.A.; et al. Germ line p53 mutation in a familial syndrome of breast cancer. Science 1990, 250, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Zou, Z.; Pirollo, K.; Blattner, W.; Chang, E.H. Germ-line transmission of a mutated p53 gene in a cancer prone family with Li-Fraumeni syndrome. Nature 1990, 348, 747–749. [Google Scholar] [CrossRef]
- Zawacka-Pankau, J.E. The Role of p53 Family in Cancer. Cancers 2022, 14, 823. [Google Scholar] [CrossRef]
- Whibley, C.; Pharoah, P.D.; Hollstein, M. p53 polymorphisms: Cancer implications. Nat. Rev. Cancer 2009, 9, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Barnoud, T.; Parris, J.L.D.; Murphy, M.E. Common genetic variants in the TP53 pathway and their impact on cancer. J. Mol. Cell Biol. 2019, 11, 578–585. [Google Scholar] [CrossRef]
- Eiholzer, R.A.; Mehta, S.; Kazantseva, M.; Drummond, C.J.; McKinney, C.; Young, K.; Slater, D.; Morten, B.C.; Avery-Kiejda, K.A.; Lasham, A.; et al. Intronic TP53 Polymorphisms Are Associated with Increased Delta133TP53 Transcript, Immune Infiltration and Cancer Risk. Cancers 2020, 12, 2472. [Google Scholar] [CrossRef]
- Freedland, S.J.; Isaacs, W.B. Explaining racial differences in prostate cancer in the United States: Sociology or biology? Prostate 2005, 62, 243–252. [Google Scholar] [CrossRef]
- Riviere, P.; Luterstein, E.; Kumar, A.; Vitzthum, L.K.; Deka, R.; Sarkar, R.R.; Bryant, A.K.; Bruggeman, A.; Einck, J.P.; Murphy, J.D.; et al. Survival of African American and non-Hispanic white men with prostate cancer in an equal-access health care system. Cancer 2020, 126, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Yamoah, K.; Lee, K.M.; Awasthi, S.; Alba, P.R.; Perez, C.; Anglin-Foote, T.R.; Robison, B.; Gao, A.; DuVall, S.L.; Katsoulakis, E.; et al. Racial and Ethnic Disparities in Prostate Cancer Outcomes in the Veterans Affairs Health Care System. JAMA Netw. Open 2022, 5, e2144027. [Google Scholar] [CrossRef]
- Cullen, J.; Young, D.; Chen, Y.; Degon, M.; Farrell, J.; Sedarsky, J.; Baptiste, W.; Rosen, P.; Tolstikov, V.; Kiebish, M.; et al. Predicting Prostate Cancer Progression as a Function of ETS-related Gene Status, Race, and Obesity in a Longitudinal Patient Cohort. Eur. Urol. Focus 2018, 4, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Loh, P.R.; Danecek, P.; Palamara, P.F.; Fuchsberger, C.A.; Reshef, Y.K.; Finucane, H.; Schoenherr, S.; Forer, L.; McCarthy, S.; Abecasis, G.R.; et al. Reference-based phasing using the haplotype reference consortium panel. Nat. Genet. 2016, 48, 1443–1448. [Google Scholar] [CrossRef]
- Taliun, D.; Harris, D.N.; Kessler, M.D.; Carlson, J.; Szpiech, Z.A.; Torres, R.; Taliun, S.A.G.; Corvelo, A.; Gogarten, S.M.; Kang, H.M.; et al. Sequencing of 53,831 diverse genomes from the NHLBI TOPMed Program. Nature 2021, 590, 290–299. [Google Scholar] [CrossRef]
- Dumont, P.; Leu, J.I.; Della Pietra, A.C.; George, D.L.; Murphy, M. The codon 72 polymorphic variants of p53 have markedly different apoptotic potential. Nat. Genet. 2003, 33, 357–365. [Google Scholar] [CrossRef]
- Hrstka, R.; Coates, P.J.; Vojtesek, B. Polymorphisms in p53 and the p53 pathway: Roles in cancer susceptibility and response to treatment. J. Cell. Mol. Med. 2009, 13, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, Z.; Atwal, G.S.; Levine, A.J. p53: A new player in reproduction. Cell Cycle 2008, 7, 848–852. [Google Scholar] [CrossRef]
- Bond, G.L.; Levine, A.J. A single nucleotide polymorphism in the p53 pathway interacts with gender, environmental stresses and tumor genetics to influence cancer in humans. Oncogene 2007, 26, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Khalil, A.; Rashid, H. Evaluation of the p53 Arg72Pro polymorphism and its association with cancer risk: A HuGE review and meta-analysis. Genet. Res. 2015, 97, e7. [Google Scholar] [CrossRef]
- Huang, S.P.; Huang, C.Y.; Wang, J.S.; Liu, C.C.; Pu, Y.S.; Yu, H.J.; Yu, C.C.; Wu, T.T.; Huang, C.H.; Wu, W.J.; et al. Prognostic significance of p53 and X-ray repair cross-complementing group 1 polymorphisms on prostate-specific antigen recurrence in prostate cancer post radical prostatectomy. Clin. Cancer Res. 2007, 13, 6632–6638. [Google Scholar] [CrossRef]
- Quiñones, L.A.; Irarrázabal, C.E.; Rojas, C.R.; Orellana, C.E.; Acevedo, C.; Huidobro, C.; Varela, N.E.; Cáceres, D.D. Joint effect among p53, CYP1A1, GSTM1 polymorphism combinations and smoking on prostate cancer risk: An exploratory genotype-environment interaction study. Asian J. Androl. 2006, 8, 349–355. [Google Scholar] [CrossRef]
- Suzuki, K.; Matsui, H.; Ohtake, N.; Nakata, S.; Takei, T.; Nakazato, H.; Okugi, H.; Koike, H.; Ono, Y.; Ito, K.; et al. A p53 codon 72 polymorphism associated with prostate cancer development and progression in Japanese. J. Biomed. Sci. 2003, 10, 430–435. [Google Scholar] [CrossRef]
- Huang, S.P.; Wu, W.J.; Chang, W.S.; Wu, M.T.; Chen, Y.Y.; Chen, Y.J.; Yu, C.C.; Wu, T.T.; Lee, Y.H.; Huang, J.K.; et al. p53 Codon 72 and p21 codon 31 polymorphisms in prostate cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 2217–2224. [Google Scholar] [CrossRef]
- Henner, W.D.; Evans, A.J.; Hough, K.M.; Harris, E.L.; Lowe, B.A.; Beer, T.M. Association of codon 72 polymorphism of p53 with lower prostate cancer risk. Prostate 2001, 49, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Cintra, H.S.; Pinezi, J.C.; Machado, G.D.; de Carvalho, G.M.; Carvalho, A.T.; dos Santos, T.E.; Marciano, R.D.; Soares Rde, B. Investigation of genetic polymorphisms related to the outcome of radiotherapy for prostate cancer patients. Dis. Markers 2013, 35, 701–710. [Google Scholar] [CrossRef]
- Han, J.Y.; Lee, G.K.; Jang, D.H.; Lee, S.Y.; Lee, J.S. Association of p53 codon 72 polymorphism and MDM2 SNP309 with clinical outcome of advanced nonsmall cell lung cancer. Cancer 2008, 113, 799–807. [Google Scholar] [CrossRef]
- Kim, J.G.; Sohn, S.K.; Chae, Y.S.; Song, H.S.; Kwon, K.Y.; Do, Y.R.; Kim, M.K.; Lee, K.H.; Hyun, M.S.; Lee, W.S.; et al. TP53 codon 72 polymorphism associated with prognosis in patients with advanced gastric cancer treated with paclitaxel and cisplatin. Cancer Chemother. Pharmacol. 2009, 64, 355–360. [Google Scholar] [CrossRef]
- Toyama, T.; Zhang, Z.; Nishio, M.; Hamaguchi, M.; Kondo, N.; Iwase, H.; Iwata, H.; Takahashi, S.; Yamashita, H.; Fujii, Y. Association of TP53codon 72 polymorphism and the outcome of adjuvant therapy in breast cancer patients. Breast Cancer Res. 2007, 9, R34. [Google Scholar] [CrossRef] [PubMed]
- Kohaar, I.; Li, Q.; Chen, Y.; Ravindranath, L.; Young, D.; Ali, A.; Sesterhenn, I.A.; Rosner, I.L.; Cullen, J.; Srivastava, S.; et al. Association of germline genetic variants with TMPRSS2-ERG fusion status in prostate cancer. Oncotarget 2020, 11, 1321–1333. [Google Scholar] [CrossRef]
- Teroerde, M.; Nientiedt, C.; Duensing, A.; Hohenfellner, M.; Stenzinger, A.; Duensing, S. Revisiting the Role of p53 in Prostate Cancer. In Prostate Cancer; Bott, S.R.J., Ng, K.L., Eds.; Exon Publications Brisbane: Brisbane City, Australia, 2021; Volume 1, pp. 116–117. ISBN 978-0-6450017-5-4. [Google Scholar] [CrossRef]
- Diakite, B.; Kassogue, Y.; Dolo, G.; Wang, J.; Neuschler, E.; Kassogue, O.; Keita, M.L.; Traore, C.B.; Kamate, B.; Dembele, E.; et al. p.Arg72Pro polymorphism of P53 and breast cancer risk: A meta-analysis of case-control studies. BMC Med. Genet. 2020, 21, 206. [Google Scholar] [CrossRef]
- Jafrin, S.; Aziz, M.A.; Anonna, S.N.; Akter, T.; Naznin, N.E.; Reza, S.; Islam, M.S. Association of TP53 codon 72 Arg>pro polymorphism with breast and lung Cancer risk in the south Asian population: A meta-analysis. Asian Pac. J. Cancer Prev. 2020, 21, 1511–1519. [Google Scholar] [CrossRef]
- Akhter, N.; Dar, S.A.; Chattopadhyay, S.; Haque, S.; Anwer, R.; Wahid, M.; Jawed, A.; Lohani, M.; Mandal, R.K.; Shukla, N.K.; et al. Impact of p53 arg72pro SNP on Breast Cancer Risk in North Indian Population. Curr. Genom. 2018, 19, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Langerød, A.; Bukholm, I.R.; Bregård, A.; Lønning, P.E.; Andersen, T.I.; Rognum, T.O.; Meling, G.I.; Lothe, R.A.; Børresen-Dale, A.L. The TP53 codon 72 polymorphism may affect the function of TP53 mutations in breast carcinomas but not in colorectal carcinomas. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1684–1688. [Google Scholar]
- Omori, S.; Yoshida, S.; Kennedy, S.H.; Negoro, K.; Hamana, S.; Barlow, D.H.; Maruo, T. Polymorphism at codon 72 of the p53 gene is not associated with endometriosis in a Japanese population. J. Soc. Gynecol. Investig. 2004, 11, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Bonafé, M.; Ceccarelli, C.; Farabegoli, F.; Santini, D.; Taffurelli, M.; Barbi, C.; Marzi, E.; Trapassi, C.; Storci, G.; Olivieri, F.; et al. Retention of the p53 codon 72 arginine allele is associated with a reduction of disease-free and overall survival in arginine/proline heterozygous breast cancer patients. Clin. Cancer Res. 2003, 9, 4860–4864. [Google Scholar]
- Siddique, M.M.; Balram, C.; Fiszer-Maliszewska, L.; Aggarwal, A.; Tan, A.; Tan, P.; Soo, K.C.; Sabapathy, K. Evidence for selective expression of the p53 codon 72 polymorphs: Implications in cancer development. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2245–2252. [Google Scholar] [CrossRef] [PubMed]
- Kyndi, M.; Alsner, J.; Hansen, L.L.; Sørensen, F.B.; Overgaard, J. LOH rather than genotypes of TP53 codon 72 is associated with disease-free survival in primary breast cancer. Acta Oncol. 2006, 45, 602–609. [Google Scholar] [CrossRef]
- Robinson, D.; Van Allen, E.M.; Wu, Y.M.; Schultz, N.; Lonigro, R.J.; Mosquera, J.M.; Montgomery, B.; Taplin, M.E.; Pritchard, C.C.; Attard, G.; et al. Integrative clinical genomics of advanced prostate cancer. Cell 2015, 161, 1215–1228. [Google Scholar] [CrossRef]
- Gesztes, W.; Schafer, C.; Young, D.; Fox, J.; Jiang, J.; Chen, Y.; Kuo, H.C.; Mwamukonda, K.B.; Dobi, A.; Burke, A.P.; et al. Focal p53 protein expression and lymphovascular invasion in primary prostate tumors predict metastatic progression. Sci. Rep. 2022, 12, 5404. [Google Scholar] [CrossRef]
- Levine, A.J. Spontaneous and inherited TP53 genetic alterations. Oncogene 2021, 40, 5975–5983. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.N.; Cheng, H.H.; Powers, J.; Gulati, R.; Ledet, E.M.; Morrison, C.; Le, A.; Hausler, R.; Stopfer, J.; Hyman, S.; et al. Inherited TP53 Variants and Risk of Prostate Cancer. Eur. Urol. 2022, 81, 243–250. [Google Scholar] [CrossRef]
- Nicolosi, P.; Ledet, E.; Yang, S.; Michalski, S.; Freschi, B.; O’Leary, E.; Esplin, E.D.; Nussbaum, R.L.; Sartor, O. Prevalence of Germline Variants in Prostate Cancer and Implications for Current Genetic Testing Guidelines. JAMA Oncol. 2019, 5, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Doffe, F.; Carbonnier, V.; Tissier, M.; Leroy, B.; Martins, I.; Mattsson, J.S.M.; Micke, P.; Pavlova, S.; Pospisilova, S.; Smardova, J.; et al. Identification and functional characterization of new missense SNPs in the coding region of the TP53 gene. Cell Death Differ. 2021, 28, 1477–1492. [Google Scholar] [CrossRef]
- Schaeffer, E.M.; Srinivas, S.; Adra, N.; An, Y.; Barocas, D.; Bitting, R.; Bryce, A.; Chapin, B.; Cheng, H.H.; D’Amico, A.V.; et al. NCCN Guidelines® Insights: Prostate Cancer, Version 1.2023. J. Natl. Compr. Cancer Netw. 2022, 20, 1288–1298. [Google Scholar] [CrossRef]
- US Preventive Services Task Force; Grossman, D.C.; Curry, S.J.; Owens, D.K.; Bibbins-Domingo, K.; Caughey, A.B.; Davidson, K.W.; Doubeni, C.A.; Ebell, M.; Epling, J.W.; et al. Screening for Prostate Cancer: US Preventive Services Task Force Recommendation Statement. JAMA 2018, 319, 1901–1913. [Google Scholar] [CrossRef]
- Carter, H.B.; Albertsen, P.C.; Barry, M.J.; Etzioni, R.; Freedland, S.J.; Greene, K.L.; Holmberg, L.; Kantoff, P.; Konety, B.R.; Murad, M.H.; et al. Early detection of prostate cancer: AUA Guideline. J. Urol. 2013, 190, 419–426. [Google Scholar] [CrossRef]
- Mersha, T.B.; Abebe, T. Self-reported race/ethnicity in the age of genomic research: Its potential impact on understanding health disparities. Hum. Genom. 2015, 9, 1. [Google Scholar] [CrossRef]
- Gnanapradeepan, K.; Leu, J.I.; Basu, S.; Barnoud, T.; Good, M.; Lee, J.V.; Quinn, W.J.; Kung, C.P.; Ahima, R.; Baur, J.A.; et al. Increased mTOR activity and metabolic efficiency in mouse and human cells containing the African-centric tumor-predisposing p53 variant Pro47Ser. Elife 2020, 9, 55994. [Google Scholar] [CrossRef]
- Tutton, S.; Deng, Z.; Gulve, N.; Vladimirova, O.; Beishline, K.; Wiedmer, A.; Murphy, M.; Lieberman, P.M. Elevated telomere dysfunction in cells containing the African-centric Pro47Ser cancer-risk variant of TP53. Oncotarget 2019, 10, 3581–3591. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.R.; Yoshioka, F.K.; Silva, R.L.; Clara, C.A.; Santos, M.J.; Almeida, J.R.; Burbano, R.R.; Rey, J.A.; Casartelli, C. Prognostic value of TP53 Pro47Ser and Arg72Pro single nucleotide polymorphisms and the susceptibility to gliomas in individuals from Southeast Brazil. Genet. Mol. Res. 2008, 7, 207–216. [Google Scholar] [CrossRef]
- Murphy, M.E.; Liu, S.; Yao, S.; Huo, D.; Liu, Q.; Dolfi, S.C.; Hirshfield, K.M.; Hong, C.C.; Hu, Q.; Olshan, A.F.; et al. A functionally significant SNP in TP53 and breast cancer risk in African-American women. NPJ Breast Cancer 2017, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.O.; Custódio, A.C.; Pinto, G.R.; Santos, M.J.; Almeida, J.R.; Clara, C.A.; Rey, J.A.; Casartelli, C. Polymorphisms and DNA methylation of gene TP53 associated with extra-axial brain tumors. Genet. Mol. Res. 2009, 8, 8–18. [Google Scholar] [CrossRef]
- Jaiswal, P.K.; Goel, A.; Mittal, R.D. Association of p53 codon 248 (exon7) with urinary bladder cancer risk in the North Indian population. Biosci. Trend 2011, 5, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Siraj, A.K.; Al-Rasheed, M.; Ibrahim, M.; Siddiqui, K.; Al-Dayel, F.; Al-Sanea, O.; Uddin, S.; Al-Kuraya, K. RAD52 polymorphisms contribute to the development of papillary thyroid cancer susceptibility in Middle Eastern population. J. Endocrinol. Investig. 2008, 31, 893–899. [Google Scholar] [CrossRef]
- Daugherty, C.L.; Curtis, H.; Realini, T.; Charlton, J.F.; Zareparsi, S. Primary open angle glaucoma in a Caucasian population is associated with the p53 codon 72 polymorphism. Mol. Vis. 2009, 15, 1939–1944. [Google Scholar] [PubMed]
- Ruggeri, R.M.; Vicchio, T.M.; Giovinazzo, S.; Certo, R.; Alibrandi, A.; Trimarchi, F.; Benvenga, S.; Trovato, M. TP53 polymorphism may contribute to genetic susceptibility to develop Hashimoto’s thyroiditis. J. Endocrinol. Investig. 2015, 38, 1175–1182. [Google Scholar] [CrossRef]
- Elsaid, A.; Zahran, R.; Elshazli, R.; El-Sayed, A.; Abou Samra, M.; El-Tarapely, F.; Abdel-Malak, C. Genetic polymorphisms of TP53 Arg72Pro and Pro47Ser among Egyptian patients with colorectal carcinoma. Arch. Physiol. Biochem. 2019, 125, 255–262. [Google Scholar] [CrossRef]
- Sameer, A.S.; Shah, Z.A.; Syeed, N.; Banday, M.Z.; Bashir, S.M.; Bhat, B.A.; Siddiqi, M.A. TP53 Pro47Ser and Arg72Pro polymorphisms and colorectal cancer predisposition in an ethnic Kashmiri population. Genet. Mol. Res. 2010, 9, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.E.; Guilhen, A.C.; de Andrade, R.A.; Sumi, L.G.; Ward, L.S. The role of TP53 PRO47SER and ARG72PRO single nucleotide polymorphisms in the susceptibility to bladder cancer. Urol. Oncol. 2011, 29, 291–294. [Google Scholar] [CrossRef]
- Alawadi, S.; Ghabreau, L.; Alsaleh, M.; Abdulaziz, Z.; Rafeek, M.; Akil, N.; Alkhalaf, M. P53 gene polymorphisms and breast cancer risk in Arab women. Med. Oncol. 2011, 28, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Mostaid, M.S.; Ahmed, M.U.; Islam, M.S.; Bin Sayeed, M.S.; Hasnat, A. Lung cancer risk in relation to TP53 codon 47 and codon 72 polymorphism in Bangladeshi population. Tumour Biol. 2014, 35, 10309–10317. [Google Scholar] [CrossRef]
- Nairuz, T.; Rahman, M.; Bushra, M.U.; Kabir, Y. TP53 Arg72Pro and XPD Lys751Gln Gene Polymorphisms and Risk of Lung Cancer in Bangladeshi Patients. Asian Pac. J. Cancer Prev. 2020, 21, 2091–2098. [Google Scholar] [CrossRef]
- Fang, Y.; Wu, X.; Li, L.; Zhu, J.; Wu, H.; Zhou, H.; He, J.; Wang, Y. TP53 Arg72Pro polymorphism and neuroblastoma susceptibility in eastern Chinese children: A three-center case-control study. Biosci. Rep. 2020, 40, BSR20200854. [Google Scholar] [CrossRef]
- Drokow, E.K.; Chen, Y.; Waqas Ahmed, H.A.; Oppong, T.B.; Akpabla, G.S.; Pei, Y.; Kumah, M.A.; Neku, E.A.; Sun, K. The relationship between leukemia and TP53 gene codon Arg72Pro polymorphism: Analysis in a multi-ethnic population. Future Oncol. 2020, 16, 923–937. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristic | N = 308 (%) |
|---|---|
| Diagnosis Age (in years) | 57 (9) |
| Unknown | 1 |
| Race | |
| African American | 212 (69%) |
| Caucasian | 95 (31%) |
| Unknown | 1 |
| Diagnosis PSA (ng/mL) | |
| 1: <4 | 72 (24%) |
| 2: 4–9 | 193 (63%) |
| 3: 10–20 | 31 (10%) |
| 4: >20 | 10 (3.3%) |
| Unknown | 2 |
| Pathologic T Stage | |
| T2 | 228 (74%) |
| T3–4 | 80 (26%) |
| Biopsy Gleason score | |
| ≤6 | 204 (70%) |
| 7 | 69 (24%) |
| 8–10 | 17 (5.9%) |
| Unknown | 18 |
| Pathologic Gleason score | |
| 3 + 3 | 169 (57%) |
| 3 + 4 | 76 (26%) |
| 4 + 3 | 24 (8.2%) |
| 8–10 | 25 (8.5%) |
| Unknown | 14 |
| BCR | 47 (15%) |
| Unknown | 3 |
| Metastasis | 9 (2.9%) |
| SNP | TP53 rs1800371 SNP | TP53 rs1042522 SNP | |||||
|---|---|---|---|---|---|---|---|
| Patient Characteristics | GG, N = 302 | GA, N = 6 | p-Value | GG, N = 87 | GC, N = 137 | CC, N = 84 | p-value |
| Diagnosis Age | 57 (9) | 56 (12) | 0.6 | 58 (9) | 57 (8) | 57 (9) | 0.8 |
| Unknown | 1 | 0 | 0 | 1 | 0 | ||
| Race | 0.2 | <0.001 | |||||
| African American | 206 (68%) | 6 (100%) | 81 (93%) | 87 (64%) | 44 (52%) | ||
| Caucasian | 95 (32%) | 0 (0%) | 6 (6.9%) | 49 (36%) | 40 (48%) | ||
| Unknown | 1 | 0 | 0 | 1 | 0 | ||
| Diagnosis PSA | 0.085 | 0.8 | |||||
| 1: <4 | 70 (23%) | 2 (33%) | 22 (25%) | 28 (21%) | 22 (26%) | ||
| 2: 4–9 | 191 (64%) | 2 (33%) | 55 (63%) | 87 (64%) | 51 (61%) | ||
| 3: 10–20 | 30 (10%) | 1 (17%) | 9 (10%) | 14 (10%) | 8 (9.5%) | ||
| 4: >20 | 9 (3.0%) | 1 (17%) | 1 (1.1%) | 6 (4.4%) | 3 (3.6%) | ||
| Unknown | 2 | 0 | 0 | 2 | 0 | ||
| Pathologic T Stage | >0.9 | 0.6 | |||||
| T2 | 223 (74%) | 5 (83%) | 57 (68%) | 104 (76%) | 59 (70%) | ||
| T3–4 | 79 (26%) | 1 (17%) | 22 (25%) | 33 (24%) | 25 (30%) | ||
| Biopsy Gleason score | 0.2 | 0.8 | |||||
| ≤6 | 199 (70%) | 5 (83%) | 57 (68%) | 93 (73%) | 54 (69%) | ||
| 7 | 69 (24%) | 0 (0%) | 21 (25%) | 27 (21%) | 21 (27%) | ||
| 8–10 | 16 (5.6%) | 1 (17%) | 6 (7.1%) | 8 (6.2%) | 3 (3.8%) | ||
| Unknown | 18 | 0 | 3 | 9 | 6 | ||
| Pathologic Gleason score | 0.6 | >0.9 | |||||
| 3 + 3 | 164 (57%) | 5 (100%) | 46 (55%) | 74 (57%) | 49 (60%) | ||
| 3 + 4 | 76 (26%) | 0 (0%) | 22 (26%) | 35 (27%) | 19 (23%) | ||
| 4 + 3 | 24 (8.3%) | 0 (0%) | 7 (8.3%) | 10 (7.8%) | 7 (8.6%) | ||
| 8–10 | 25 (8.7%) | 0 (0%) | 9 (11%) | 10 (7.8%) | 6 (7.4%) | ||
| Unknown | 13 | 1 | 3 | 8 | 3 | ||
| BCR | 47 (16%) | 0 (0%) | 0.6 | 13 (15%) | 20 (15%) | 14 (17%) | >0.9 |
| Unknown | 3 | 0 | 1 | 1 | 1 | ||
| Metastasis | 8 (2.6%) | 1 (17%) | 0.2 | 3 (3.4%) | 6 (4.4%) | 0 (0%) | 0.2 |
| Position | SNP | Location | HR | p-Value |
|---|---|---|---|---|
| chr17:7667612:A:G | rs4968186 | downstream | 1.753 | 0.004 |
| chr17:7676963:G:A | rs9894227 | intronic | 1.720 | 0.007 |
| chr17:7676734:C:T | rs8079544 | intronic | 1.632 | 0.020 |
| chr17:7676154:G:C | rs1042522 | exonic | 1.523 | 0.046 |
| chr17:7664197:C:T | rs35119871 | intergenic | 1.461 | 0.046 |
| chr17:7675519:A:G | rs1794287 | intronic | 1.508 | 0.058 |
| chr17:7676230:G:A | rs1800371 | exonic | 1.473 | 0.067 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duncan, A.; Nousome, D.; Ricks, R.; Kuo, H.-C.; Ravindranath, L.; Dobi, A.; Cullen, J.; Srivastava, S.; Chesnut, G.T.; Petrovics, G.; et al. Association of TP53 Single Nucleotide Polymorphisms with Prostate Cancer in a Racially Diverse Cohort of Men. Biomedicines 2023, 11, 1404. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11051404
Duncan A, Nousome D, Ricks R, Kuo H-C, Ravindranath L, Dobi A, Cullen J, Srivastava S, Chesnut GT, Petrovics G, et al. Association of TP53 Single Nucleotide Polymorphisms with Prostate Cancer in a Racially Diverse Cohort of Men. Biomedicines. 2023; 11(5):1404. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11051404
Chicago/Turabian StyleDuncan, Allison, Darryl Nousome, Randy Ricks, Huai-Ching Kuo, Lakshmi Ravindranath, Albert Dobi, Jennifer Cullen, Shiv Srivastava, Gregory T. Chesnut, Gyorgy Petrovics, and et al. 2023. "Association of TP53 Single Nucleotide Polymorphisms with Prostate Cancer in a Racially Diverse Cohort of Men" Biomedicines 11, no. 5: 1404. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11051404