
Association between TGFβ1 Levels in Cord Blood and Weight Progress in the First Year of Life
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study Subjects
2.3. Assays and Laboratory Measurements
2.4. Statistical Analysis
3. Results
3.1. Association between Maternal and Cord TGFβ1 Levels
3.2. Association between Maternal TGFβ1 Levels and Child Growth
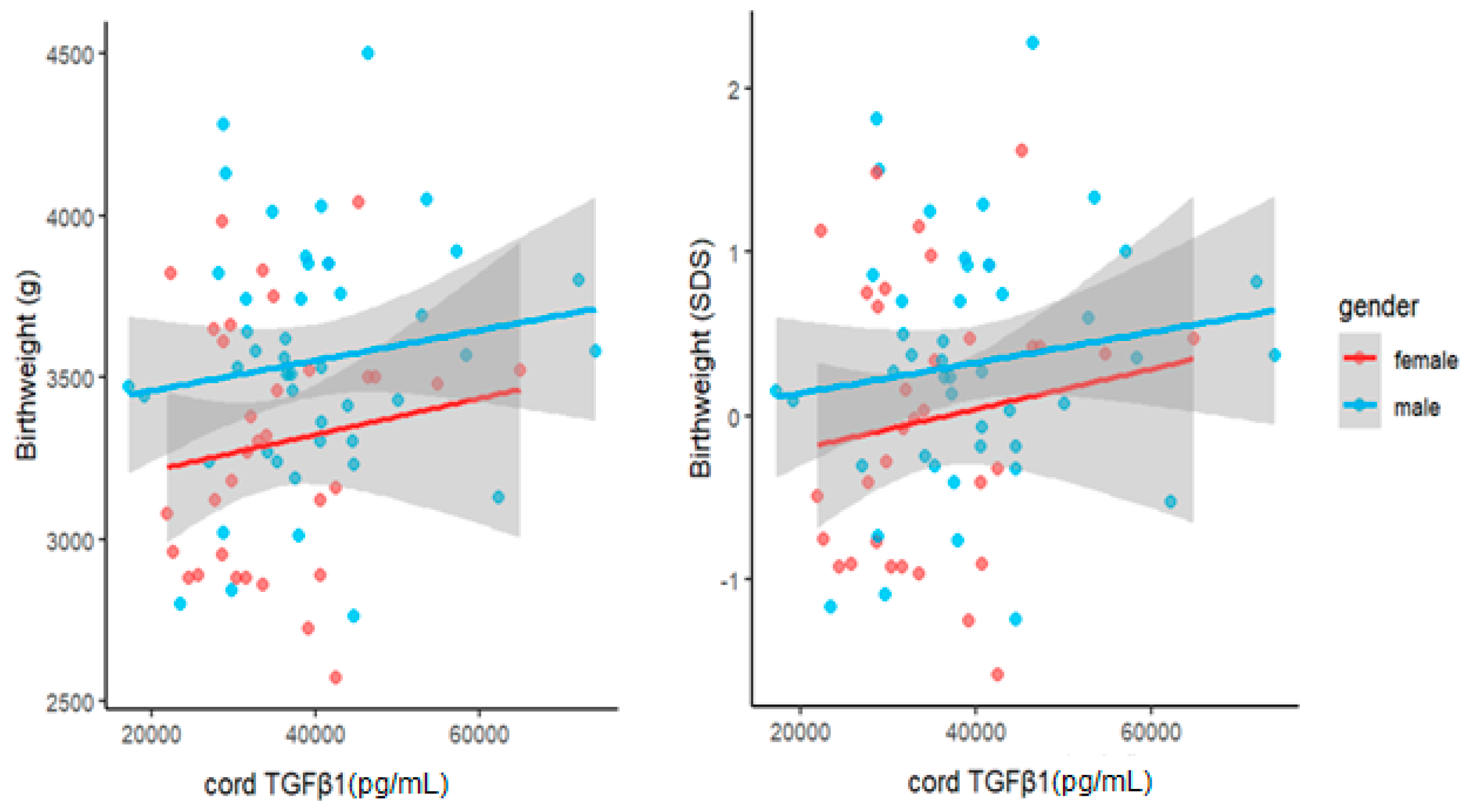
3.3. Association between Cord TGFβ1 Levels and Child Growth
3.4. Association between Maternal and Cord TGFβ1 Levels and Maternal Parameters
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fasshauer, M.; Blüher, M. Adipokines in Health and Disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Würfel, M.; Blüher, M.; Stumvoll, M.; Ebert, T.; Kovacs, P.; Tönjes, A.; Breitfeld, J. Adipokines as Clinically Relevant Therapeutic Targets in Obesity. Biomedicines 2023, 11, 1427. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, N.; Blüher, M.; Stepan, H.; Stumvoll, M.; Ebert, T.; Tönjes, A.; Schrey-Petersen, S. Adipokines in Pregnancy: A Systematic Review of Clinical Data. Biomedicines 2023, 11, 1419. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The Fetal and Infant Orgins of Adult Disease. Br. Med. J. 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myatt, L. Placental Adaptive Responses and Fetal Programming. J. Physiol. 2006, 572, 25–30. [Google Scholar] [CrossRef]
- Mahmoud Sindiani, A.; Obeidat, R.; Jbarah, O.; Hazaimeh, E. Relationship between Newborn Leptin Levels and Selected Growth Parameters. J. Int. Med. Res. 2019, 47, 2591–2597. [Google Scholar] [CrossRef]
- Joung, K.E.; Cataltepe, S.U.; Michael, Z.; Christou, H.; Mantzoros, C.S. Cord Blood Adipocyte Fatty Acid–Binding Protein Levels Correlate with Gestational Age and Birth Weight in Neonates. J. Clin. Endocrinol. Metab. 2017, 102, 1606–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrey-Petersen, S.; Bäumer, S.; Lössner, U.; Stepan, H. Adipocyte Fatty Acid-Binding Protein 4 Is Altered in Growth Discordant Dichorionic, but Not in Monochorionic Twins. J. Endocr. Soc. 2020, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Massague, J. The Transforming Growth Factor-β Family. Annu. Rev. Cell Biol. 1990, 6, 597–641. [Google Scholar] [CrossRef]
- Kingsley, D.M. The TGF-β Superfamily: New Members, New Receptors, and New Genetic Tests of Function in Different Organisms. Genes Dev. 1994, 8, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Hogg, N.; Browning, J.; Howard, T.; Winterford, C.; Fitzpatrick, D.; Gobé, G. Apoptosis in Vascular Endothelial Cells Caused by Serum Deprivation, Oxidative Stress and Transforming Growth Factor-β. Endothelium 1999, 7, 35–49. [Google Scholar] [CrossRef]
- Assoian, R.K.; Komoriya, A.; Meyers, C.A.; Miller, D.M.; Sporn, M.B. Transforming Growth Factor-β in Human Platelets. Identification of a Major Storage Site, Purification, and Characterization. J. Biol. Chem. 1983, 258, 7155–7160. [Google Scholar] [CrossRef]
- Moses, H.L.; Yang, E.Y.; Pietenpol, J.A. TGF-β Stimulation and Inhibition of Cell Proliferation: New Mechanistic Insights Minireview. Cell 1990, 63, 245–247. [Google Scholar] [CrossRef]
- Fain, J.N.; Tichansky, D.S.; Madan, A.K. Transforming Growth Factor Β1 Release by Human Adipose Tissue Is Enhanced in Obesity. Metabolism 2005, 54, 1546–1551. [Google Scholar] [CrossRef] [PubMed]
- Felicidade, I.; Bocchi, M.; Rizzon, M.; Ramos, Z.; Carlos, L.D.O.; Ramori, N.; Wagner, F.; Campos, A.C.L. Transforming Growth Factor Beta 1 (TGFβ1) Plasmatic Levels and Haplotype Structures in Obesity: A Role for TGFβ1 in Steatosis Development. Mol. Biol. Rep. 2021, 48, 6401–6411. [Google Scholar] [CrossRef]
- Wang, H.-L.; Wang, L.; Zhao, C.-Y.; Lan, H.-Y.; Bartolomé, A. Role of TGF-Beta Signaling in Beta Cell Proliferation and Function in Diabetes. Biomolecules 2022, 12, 373. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.L.; Stoikos, C.; Findlay, J.K.; Salamonsen, L.A. TGF-β Superfamily Expression and Actions in the Endometrium and Placenta. Reproduction 2006, 132, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.B.; Heine, U.I.; Flanders, K.C.; Sporn, M.B. Transforming Growth Factor-β. Major Role in Regulation of Extracellular Matrix. Ann. N. Y. Acad. Sci. 1990, 580, 225–232. [Google Scholar] [CrossRef]
- Ha, C.T.; Waterhouse, R.; Wessells, J.; Wu, J.A.; Dveksler, G.S. Binding of Pregnancy-Specific Glycoprotein 17 to CD9 on Macrophages Induces Secretion of IL-10, IL-6, PGE2, and TGF-Β1. J. Leukoc. Biol. 2005, 77, 948–957. [Google Scholar] [CrossRef] [Green Version]
- Snyder, S.K.; Wessner, D.H.; Wessells, J.L.; Waterhouse, R.M.; Wahl, L.M.; Zimmermann, W.; Dveksler, G.S. Pregnancy-Specific Glycoproteins Function as Immunomodulators by Inducing Secretion of IL-10, IL-6 and TGF-Β1 by Human Monocytes. Am. J. Reprod. Immunol. 2001, 45, 205–216. [Google Scholar] [CrossRef]
- Chen, L.; Dupre, A.; Qiu, X.; Pellon-Cardenas, O.; Walton, K.D.; Wang, J.; Perekatt, A.O.; Hu, W.; Spence, J.R.; Verzi, M.P. TGFB1 Induces Fetal Reprogramming and Enhances Intestinal Regeneration. bioRxiv 2023. [Google Scholar] [CrossRef]
- LIFE Child-Forschungsprojekt für Zivilisationskrankheiten. Available online: https://home.uni-leipzig.de/lifechild/ (accessed on 2 June 2023).
- Würbach, A.; Zellner, K.; Kromeyer-Hauschild, K. Meal Patterns among Children and Adolescents and Their Associations with Weight Status and Parental Characteristics. Public Health Nutr. 2009, 12, 1115–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulain, T.; Baber, R.; Vogel, M.; Pietzner, D.; Kirsten, T.; Jurkutat, A.; Hiemisch, A.; Hilbert, A.; Kratzsch, J.; Thiery, J.; et al. The LIFE Child Study: A Population-Based Perinatal and Pediatric Cohort in Germany. Eur. J. Epidemiol. 2017, 32, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Briana, D.D.; Papathanasiou, A.E.; Gavrili, S.; Georgantzi, S.; Marmarinos, A.; Christou, C.; Voulgaris, K.; Gourgiotis, D.; Malamitsi-Puchner, A. Preadipocyte Factor-1 in Maternal, Umbilical Cord Serum and Breast Milk: The Impact of Fetal Growth. Cytokine 2019, 114, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Eirini Papathanasiou, A.; Malamitsi-Puchner, A.; Gavrili, S.; Zachaki, S.; Georgantzi, S.; Marmarinos, A.; Christou, C.; Voulgaris, K.; Gourgiotis, D.; Briana, D.D. Perinatal Lipocalin-2 Profile at the Extremes of Fetal Growth. J. Matern. Fetal Neonatal Med. 2021, 34, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Power, L.L.; Popplewell, E.J.; Holloway, J.A.; Diaper, N.D.; Warner, J.O.; Jones, C.A. Immunoregulatory Molecules during Pregnancy and at Birth. J. Reprod. Immunol. 2002, 56, 19–28. [Google Scholar] [CrossRef]
- Tutdibi, E.; Hunecke, A.; Lindner, U.; Monz, D.; Gortner, L. Levels of Cytokines in Umbilical Cord Blood in Relation to Spontaneous Term Labor. J. Perinat. Med. 2012, 40, 527–532. [Google Scholar] [CrossRef]
- Briana, D.D.; Liosi, S.; Gourgiotis, D.; Boutsikou, M.; Marmarinos, A.; Baka, S.; Hassiakos, D.; Malamitsi-Puchner, A. Fetal Concentrations of the Growth Factors TGF-α and TGF-Β1 in Relation to Normal and Restricted Fetal Growth at Term. Cytokine 2012, 60, 157–161. [Google Scholar] [CrossRef]
- Simpson, H.; Robson, S.C.; Bulmer, J.N.; Barber, A.; Lyall, F. Transforming Growth Factor β Expression in Human Placenta and Placental Bed during Early Pregnancy. Placenta 2002, 23, 44–58. [Google Scholar] [CrossRef]
- Morrish, D.W.; Bhardwaj, D.; Paras, M.T. Transforming Growth Factor β1 Inhibits Placental Differentiation and Human Chorionic Gonadotropin and Human Placental Lactogen Secretion. Endocrinology 1991, 129, 22–26. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.H.; Rane, S.G. TGF-β Signaling in Pancreatic Islet β Cell Development and Function. Endocrinology 2021, 162, bqaa233. [Google Scholar] [CrossRef] [PubMed]
- Mandò, C.; Abati, S.; Anelli, G.M.; Favero, C.; Serati, A.; Dioni, L.; Zambon, M.; Albetti, B.; Bollati, V.; Cetin, I. Epigenetic Profiling in the Saliva of Obese Pregnant Women. Nutrients 2022, 14, 2122. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, J.; Li, H.; Wang, T. Identification of Key Genes in Pathogenesis of Placental Insufficiency Intrauterine Growth Restriction. BMC Pregnancy Childbirth 2022, 22, 77. [Google Scholar] [CrossRef] [PubMed]
- Rab, A.; Szentpéteri, I.; Kornya, L.; Börzsönyi, B.; Demendi, C.; Valent, S.; Zsom, L.; Hejja, H.; Joó, J.G. Placental Gene Expression of Transforming Growth Factor Beta 1 (TGF-Β1) in Small for Gestational Age Newborns. J. Matern. Fetal Neonatal Med. 2015, 28, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.J.; Kim, Y.J. What Is Fetal Programming?: A Lifetime Health Is under the Control of in Utero Health. Obstet. Gynecol. Sci. 2017, 60, 506. [Google Scholar] [CrossRef]
- Reynolds, L.P.; Borowicz, P.P.; Caton, J.S.; Crouse, M.S.; Dahlen, C.R.; Ward, A.K. Developmental Programming of Fetal Growth and Development. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 229–247. [Google Scholar] [CrossRef]
- Marciniak, A.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Marciniak, B.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Fetal Programming of the Metabolic Syndrome. Taiwan J. Obstet. Gynecol. 2017, 56, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Kratzsch, J.; Höckel, M.; Kiess, W. Leptin and Pregnancy Outcome. Curr. Opin. Obstet. Gynecol. 2000, 12, 501–505. [Google Scholar] [CrossRef]
- Dejani, N.N.; Nicoletti, C.F.; Argentato, P.P.; Pereira, L.d.S.; Saraiva, A.C.; de Assis, L.M.; Nakandakare, P.Y.; Batista, L.P.R.; Teles, L.d.F.d.S.; Leitão, M.P.; et al. Maternal Plasma Transforming Growth Factor-Β1 (TGF-Β1) and Newborn Size: The Araraquara Cohort Study. J. Pediatr. 2023, 99, 284–288. [Google Scholar] [CrossRef]
- Singh, M.; Orazulike, N.C.; Ashmore, J.; Konje, J.C. Changes in Maternal Serum Transforming Growth Factor Beta-1 during Pregnancy: A Cross-Sectional Study. Biomed. Res. Int. 2013, 2013, 318464. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.; Hefler, L.; Tempfer, C.; Zeisler, H.; Lebrecht, A.; Husslein, P. Transforming Growth Factor-Beta 1 Serum Levels in Pregnancy and Pre-Eclampsia. Acta Obstet. Gynecol. Scand. 2002, 81, 168–171. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean | Min | Max | 25th Quartile | 75th Quartile | SD | ||
|---|---|---|---|---|---|---|---|
| Maternal Parameters: n = 79 | |||||||
| Maternal age | years | 30.02 | 18.74 | 41.55 | 26.69 | 32.45 | 4.649 |
| GWG (n = 72) | kg | 14.97 | 3.000 | 31.00 | 11.00 | 17.50 | 5.729 |
| GA | weeks | 39.80 | 37.00 | 41.70 | 38.70 | 40.90 | 1.218 |
| Serum TGFβ1 at 36 Weeks gestation | pg/mL | 32,310 | 19,172 | 50,542 | 26,480 | 37,453 | 7073 |
| Child Parameters at birth: n = 79: female = 33, male = 46 | |||||||
| Weight | g | 3445 | 2570 | 4500 | 3160 | 3740 | 397.7 |
| Length | cm | 49.86 | 46.00 | 55.00 | 49.00 | 51.00 | 1.824 |
| BMI | kg/m2 | 13.83 | 11.20 | 16.71 | 13.05 | 14.56 | 1.148 |
| Cord TGFβ1 | pg/mL | 37,738 | 17,238 | 74,115 | 29,696 | 42,534 | 11,186 |
| Child Parameters at U3: n = 79 | |||||||
| Age | years | 0.086 | 0.024 | 0.153 | 0.064 | 0.109 | 0.030 |
| Weight | g | 4419 | 3140 | 6600 | 4070 | 4700 | 579.9 |
| Length | cm | 54.61 | 49.40 | 59.50 | 53.00 | 56.00 | 2.008 |
| BMI | kg/m2 | 14.77 | 12.20 | 19.76 | 13.79 | 15.57 | 1.340 |
| Child Parameters at U6: n = 78 | |||||||
| Age | years | 0.959 | 0.766 | 1.177 | 0.9075 | 0.999 | 0.074 |
| Weight | g | 9391 | 7020 | 11,940 | 8699 | 10,113 | 980.7 |
| Length | cm | 74.73 | 68.00 | 80.00 | 73.00 | 76.50 | 2.708 |
| BMI | kg/m2 | 16.79 | 14.15 | 20.09 | 15.82 | 17.63 | 1.255 |
| β10,000 | p | R2 Adjusted | ||
|---|---|---|---|---|
| Variable: Birthweight | 0.20 | |||
| Predictor: | Cord TGFβ1 | 88 | 0.02 | |
| GA | 14 × 105 | <10−3 | ||
| Variable: U3 weight (1 month) | 0.30 | |||
| Predictor: | Cord TGFβ1 | 156 | <10−2 | |
| Male sex | 49 × 105 | <10−3 | ||
| Variable: U6 weight (1 year) | 0.19 | |||
| Predictor: | Cord TGFβ1 | 201 | 0.03 | |
| Male sex | 68 × 105 | <10−2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabbani, N.; Stepan, H.; Blüher, M.; Ebert, T.; Baber, R.; Vogel, M.; Kiess, W.; Stumvoll, M.; Breitfeld, J.; Lössner, U.; et al. Association between TGFβ1 Levels in Cord Blood and Weight Progress in the First Year of Life. Biomedicines 2023, 11, 2220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11082220
Kabbani N, Stepan H, Blüher M, Ebert T, Baber R, Vogel M, Kiess W, Stumvoll M, Breitfeld J, Lössner U, et al. Association between TGFβ1 Levels in Cord Blood and Weight Progress in the First Year of Life. Biomedicines. 2023; 11(8):2220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11082220
Chicago/Turabian StyleKabbani, Noura, Holger Stepan, Matthias Blüher, Thomas Ebert, Ronny Baber, Mandy Vogel, Wieland Kiess, Michael Stumvoll, Jana Breitfeld, Ulrike Lössner, and et al. 2023. "Association between TGFβ1 Levels in Cord Blood and Weight Progress in the First Year of Life" Biomedicines 11, no. 8: 2220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11082220