Outcomes of Gallic Acid on Alternariol Induced Cyto-Morphic and Genotoxic In Vivo Changes in Parotid Gland: 4-HNE Incorporated


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals
2.3. Experimental Procedure
2.4. Histopathological Study
2.4.1. Haematoxylin and Eosin (H&E) Stain
2.4.2. Immunohistochemical Study
2.5. Ultrastructure Study
2.6. Alkaline Single Cell Gel Electrophoresis (Comet Assay)
2.7. Biochemical Analysis of Tissue Antioxidant Enzymes
2.8. Morphometric Study
2.9. Statistical Analysis
3. Results
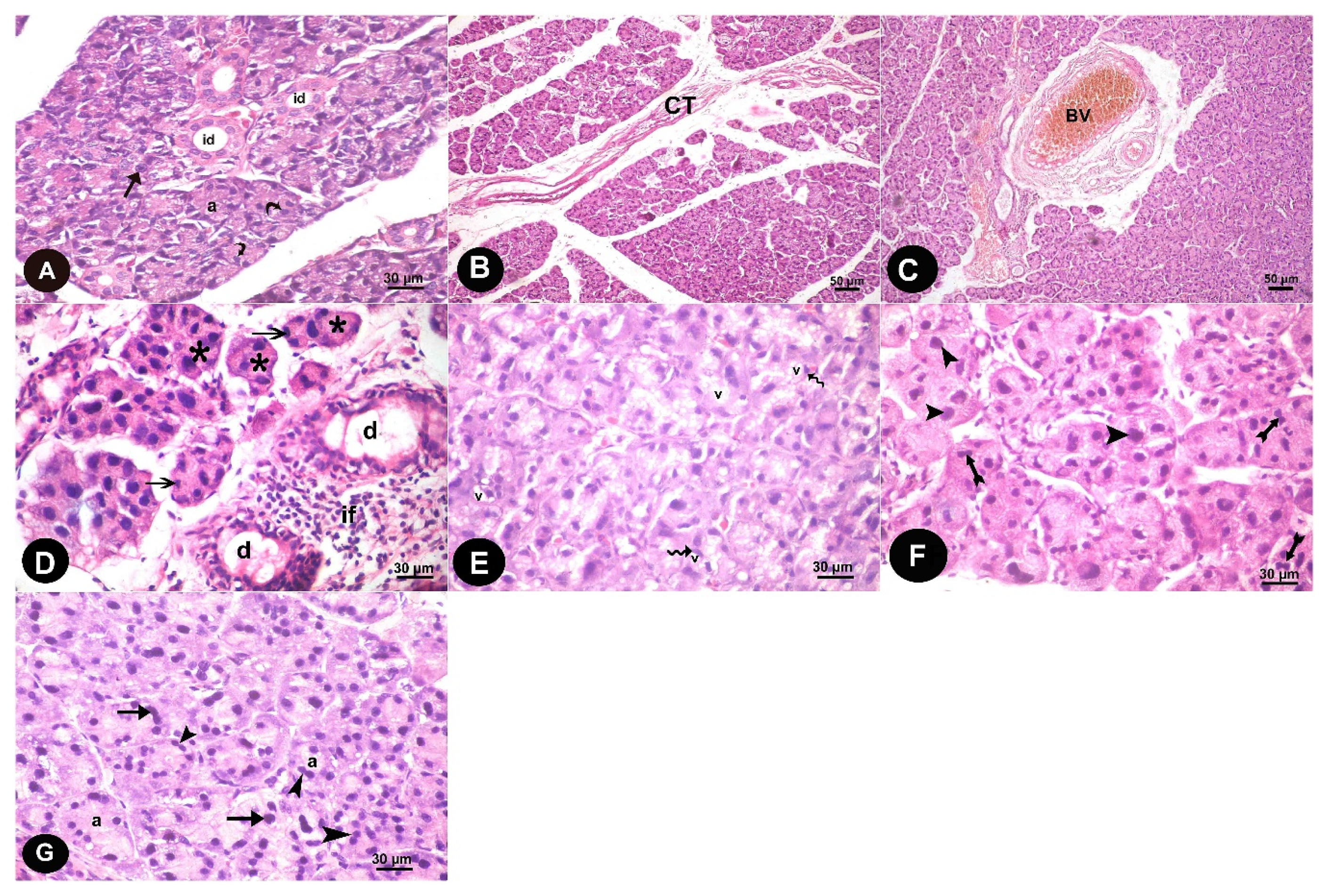
3.1. Histopathological Results
3.1.1. Light Microscope Results
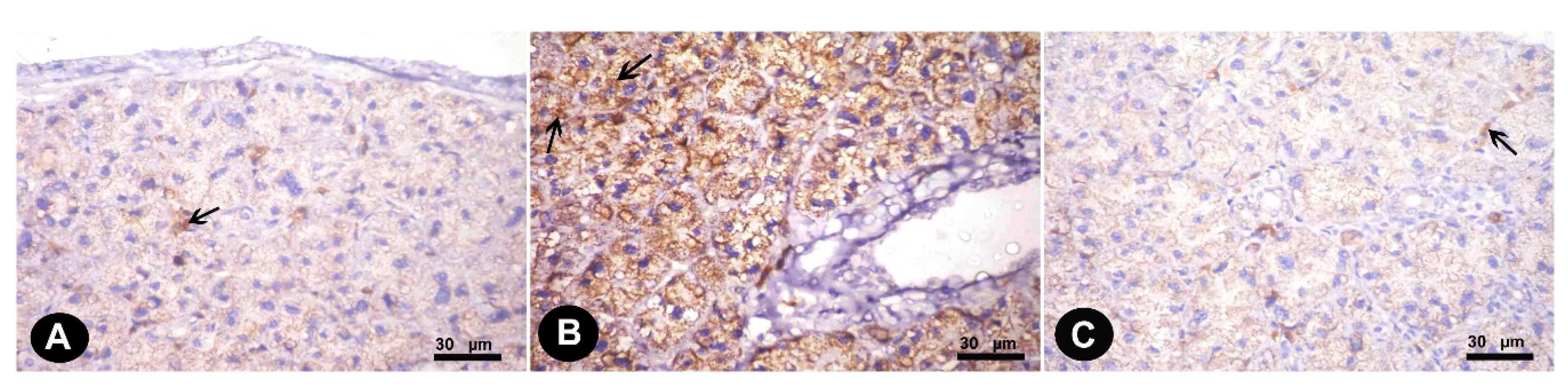
3.1.2. Immunohistochemical Results
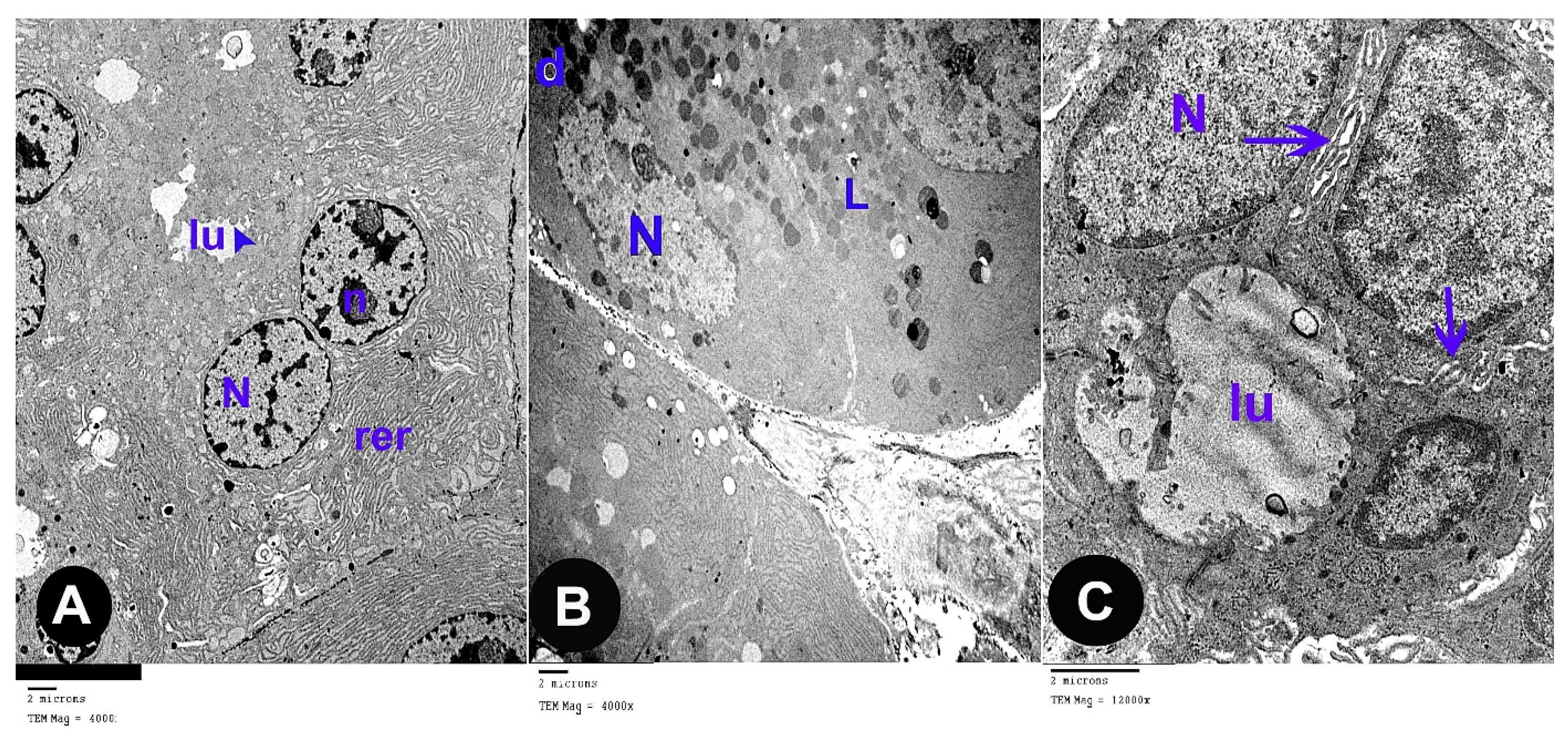
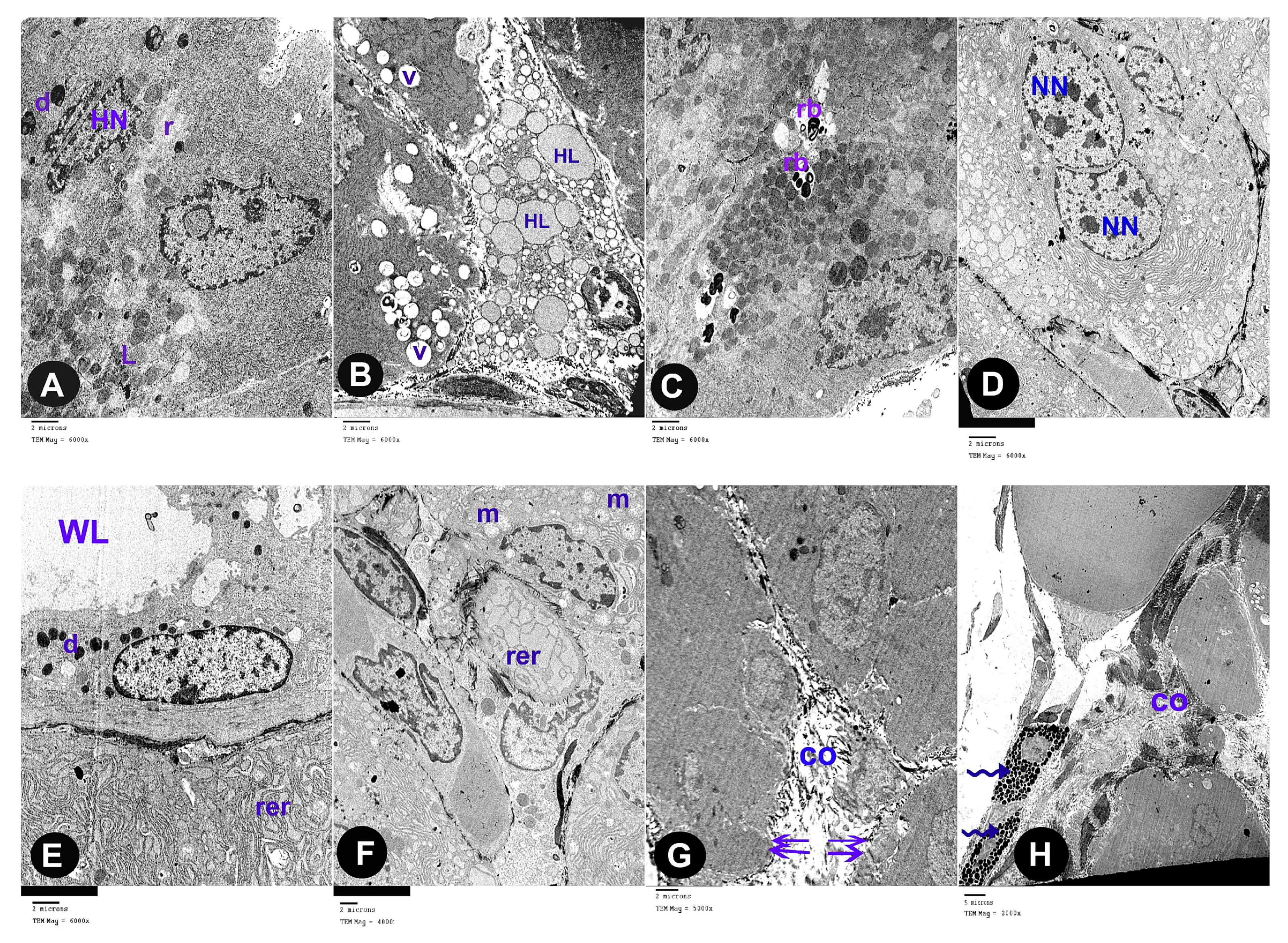
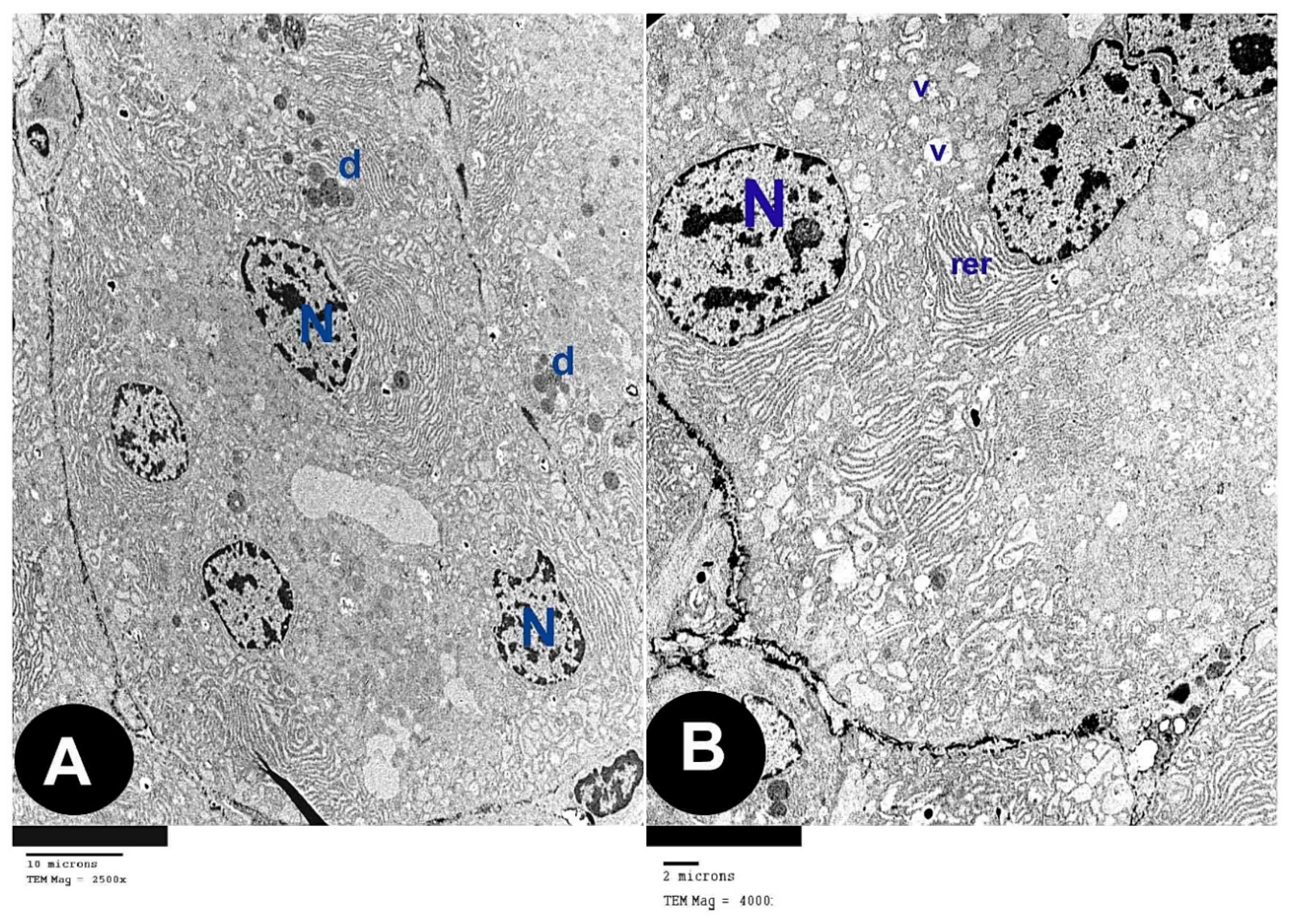
3.2. Electron Microscope Results
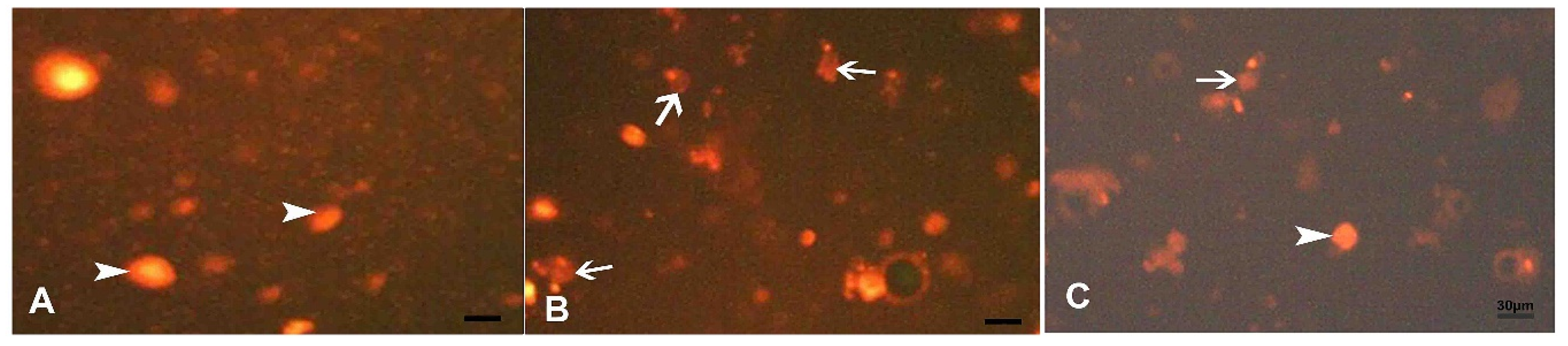
3.3. Results and Statistical Analysis of Comet Assay
3.4. Morphometric Results
3.5. Biochemical Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AOH | Alternariol |
| GA | gallic acid |
| 4-HNE | 4-hyroxynonenal |
| MDA | Malondialdehyde |
| CAT | Catalase |
| SOD | superoxide dismutase |
References
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Oswald, I.P. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef]
- Yin, Y.N.; Yan, L.Y.; Jiang, J.H.; Ma, Z.H. Biological control of aflatoxin contamination of crops. J. Zhejiang Univ. Sci. B 2008, 9, 787–792. [Google Scholar] [CrossRef]
- Mateo, R.; Medina, Á.; Mateo, E.M.; Mateo, F.; Jiménez, M. An overview of ochratoxin A in beer and wine. Int. J. Food Microbial. 2007, 119, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, A.W.; Hooker, D.C. Climatic models to predict occurrence of Fusarium toxins in wheat and maize. Int. J. Food Microbial. 2007, 119, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Troncoso-Rojas, R.; Tiznado-Hernández, M.E. Postharvest Decay; Chapter 5-Alternaria alternata (black rot, black spot); Elsevier: Amsterdam, The Netherlands, 2014; pp. 147–187. [Google Scholar]
- López, P.; Venema, D.; de Rijk, T.; de Kok, A.; Scholten, J.M.; Mol, H.G.; de Nijs, M. Occurrence of Alternaria toxins in food products in The Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- Ackermann, Y.; Curtui, V.; Dietrich, R.; Gross, M.; Latif, H.; Märtlbauer, E.; Usleber, E. Widespread occurrence of low levels of alternariol in apple and tomato products, as determined by comparative immunochemical assessment using monoclonal and polyclonal antibodies. J. Agric. food Chem. 2011, 59, 6360–6368. [Google Scholar] [CrossRef]
- Müller, M.E.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the northeast of Germany. Food Control 2013, 34, 191–197. [Google Scholar] [CrossRef]
- Marín, S.; Ramos, A.J. Food Hygiene and Toxicology in Ready-to-Eat Foods; Molds and mycotoxins in nuts; Elsevier: Amsterdam, The Netherlands, 2016; pp. 295–312. [Google Scholar]
- EFSA. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- Lehmann, L.; Wagner, J.; Metzler, M. Estrogenic and clastogenic potential of the mycotoxin alternariol in cultured mammalian cells. Food Chem. Toxicol. 2006, 44, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Ndossi, D.; Kalayou, S.; Eriksen, G.S.; Verhaegen, S.; Sørlie, M.; Connolly, L. An in vitro investigation of endocrine disrupting effects of the mycotoxin alternariol. Toxicol. Appl. Pharmacol. 2013, 271, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Wang, L.F. Theoretical elucidation on structure–antioxidant activity relationships for indolinonic hydroxylamines. Bioorg. Med. Chem. Lett. 2002, 12, 225–227. [Google Scholar] [CrossRef]
- Vaquero, M.R.; Alberto, M.R.; De Nadra, M.M. Antibacterial effect of phenolic compounds from different wines. Food Control 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Liu, K.Y.; Hu, S.; Chan, B.C.; Wat, E.C.; Lau, C.; Hon, K.L.; Wong, C.K. Anti-inflammatory and anti-allergic activities of Pentaherb formula, Moutan Cortex (Danpi) and gallic acid. Molecules 2013, 18, 2483–2500. [Google Scholar] [CrossRef]
- Guimaraes, T.A.; Farias, L.C.; Fraga, C.A.; Feltenberger, J.D.; Melo, G.A.; Coletta, R.D.; Guimaraes, A.L. Evaluation of the antineoplastic activity of gallic acid in oral squamous cell carcinoma under hypoxic conditions. Anticancer Drugs 2016, 27, 407–416. [Google Scholar] [CrossRef]
- Tsai, C.L.; Chiu, Y.M.; Ho, T.Y.; Hsieh, C.T.; Shieh, D.C.; Lee, Y.J.; Wu, Y.Y. Gallic acid induces apoptosis in human gastric adenocarcinoma cells. Anticancer Res. 2018, 38, 2057–2067. [Google Scholar]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef]
- Hsu, C.L.; Yen, G.C. Effect of gallic acid on high fat diet-induced dyslipidaemia, hepatosteatosis and oxidative stress in rats. Br. J. Nutr. 2007, 98, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.; Asokkumar, K.; Umamaheswari, M.; Sivashanmugam, A.T.; Subhadradevi, V. Antiulcerogenic effect of gallic acid in rats and its effect on oxidant and antioxidant parameters in stomach tissue. Indian J. Pharm. Sci. 2013, 75, 149. [Google Scholar]
- Ozmen, A.; Canbek, M.; Senturk, H.; Bayramoglu, G.; Uyanoglu, M. Possible protective effects of gallic acid against hepatic ischemia reperfusion injury in rats. Spatula DD 2012, 2, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Puntscher, H.; Hankele, S.; Tillmann, K.; Attakpah, E.; Braun, D.; Kütt, M.L.; Marko, D. First insights into Alternaria multi-toxin in vivo metabolism. Toxicol. Lett. 2019, 301, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Institutional Animal Care and Use Committee (IACUC). Office of Research Compliance (ORC) 2013. Available online: https://research.iu.edu/doc/compliance/animal-care/iupui/iupui-iacuc-euthanasia.pdf (accessed on 1 September 2019).
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill Livingstone: London, UK; New York, NY, USA; Philadelphia, PA, USA, 2008. [Google Scholar]
- Ramos-Vara, J.A.; Kiupel, M.; Baszler, T.; Bliven, L.; Brodersen, B.; Chelack, B.; Ehrhart, E.J. Suggested guidelines for immunohistochemical techniques in veterinary diagnostic laboratories. J. Vet. Diagn. Investig. 2008, 20, 393–413. [Google Scholar] [CrossRef] [PubMed]
- Ayache, J.; Beaunier, L.; Boumendil, J.; Ehret, G.; Laub, D. Sample Preparation Handbook for Transmission Electron Microscopy Techniques; Springer Science and Businesss Media: New York, NY, USA, 2010. [Google Scholar]
- Jackson, P.; Pedersen, L.M.; Kyjovska, Z.O.; Jacobsen, N.R.; Saber, A.T.; Hougaard, K.S.; Wallin, H. Validation of freezing tissues and cells for analysis of DNA strand break levels by comet assay. Mutagenesis 2013, 28, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Dietary exposure to Alternaria toxins. EFSA J. 2016, 14, e04654. [Google Scholar]
- Schuchardt, S.; Ziemann, C.; Hansen, T. Combined toxicokinetic and in vivo genotoxicity study on Alternaria toxins. EFSA Support. Publ. 2014, 11, 679E. [Google Scholar] [CrossRef]
- de Paula, F.; Teshima, T.H.N.; Hsieh, R.; Souza, M.M.; Nico, M.M.S.; Lourenco, S.V. Overview of human salivary glands: Highlights of morphology and developing processes. Anat. Rec. (Hoboken) 2017, 300, 1180–1188. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Oxidative stress of alternariol in Caco-2 cells. Toxicol. Lett. 2014, 229, 458–464. [Google Scholar] [CrossRef]
- Tiessen, C.; Fehr, M.; Schwarz, C.; Baechler, S.; Domnanich, K.; Böttler, U.; Marko, D. Modulation of the cellular redox status by the Alternaria toxins alternariol and alternariol monomethyl ether. Toxicol. Lett. 2013, 216, 23–30. [Google Scholar] [CrossRef]
- Burkhardt, B.; Wittenauer, J.; Pfeiffer, E.; Schauer, U.M.; Metzler, M. Oxidative metabolism of the mycotoxins alternariol and alternariol-9-methyl ether in precision-cut rat liver slices in vitro. Mol. Nutr. Food Res. 2011, 55, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Fussell, K.C.; Udasin, R.G.; Smith, P.J.; Gallo, M.A.; Laskin, J.D. Catechol metabolites of endogenous estrogens induce redox cycling and generate reactive oxygen species in breast epithelial cells. Carcinogenesis 2011, 32, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambudkar, I.S. Ca2+ signaling and regulation of fluid secretion in salivary gland acinar cells. Cell Calcium 2014, 55, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.F.; Ho, K.Y.; Ho, Y.P.; Wu, Y.M.; Yang, Y.H.; Tsai, C.C. The investigation of glutathione peroxidase, lactoferrin, myeloperoxidase and interleukin-1β in gingival crevicular fluid: Implications for oxidative stress in human periodontal diseases. J. Periodontal Res. 2004, 39, 287–293. [Google Scholar] [CrossRef]
- Solhaug, A.; Vines, L.L.; Ivanova, L.; Spilsberg, B.; Holme, J.A.; Pestka, J.; Collins, A.; Eriksen, G.S. Mechanisms involved in alternariol-induced cell cycle arrest. Mutat. Res. 2012, 738, 1–11. [Google Scholar] [CrossRef]
- Sohar, N.; Sohar, I.; Hammer, H. Lysosomal enzyme activities: New potential markers for Sjögren’s syndrome. Clin. Biochem. 2005, 38, 1120–1126. [Google Scholar] [CrossRef]
- Solhaug, A.; Wisbech, C.; Christoffersen, T.E.; Hult, L.O.; Lea, T.; Eriksen, G.S.; Holme, J.A. The mycotoxin alternariol induces DNA damage and modify macrophage phenotype and inflammatory responses. Toxicol. Lett. 2015, 239, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Bansal, M.; Singh, N.; Alam, S.; Pal, S.; Satyanarayana, G.N.V.; Singh, D.; Ansari, K.M. Alternariol induced proliferation in primary mouse keratinocytes and inflammation in mouse skin is regulated via PGE2/EP2/cAMP/p-CREB signaling pathway. Toxicology 2019, 412, 79–88. [Google Scholar] [CrossRef]
- Schrader, T.J.; Cherry, W.; Soper, K.; Langlois, I. Further examination of the effects of nitrosylation on Alternaria alternata mycotoxin mutagenicity in vitro. Mutat. Res. 2006, 606, 61–71. [Google Scholar] [CrossRef]
- Solhaug, A.; Holme, J.A.; Haglund, K.; Dendele, B.; Sergent, O.; Pestka, J.; Eriksen, G.S. Alternariol induces abnormal nuclear morphology and cell cycle arrest in murine RAW 264.7 macrophages. Toxicol. Lett. 2013, 219, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Brugger, E.M.; Wagner, J.; Schumacher, D.M.; Koch, K.; Podlech, J.; Metzler, M.; Lehmann, L. Mutagenicity of the mycotoxin alternariol in cultured mammalian cells. Toxicol. Lett. 2006, 164, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Sauter, F.; Pfeiffer, E.; Metzler, M.; Hartwig, A.; Köberle, B. DNA damage and repair kinetics of the Alternaria mycotoxins alternariol, altertoxin II and stemphyltoxin III in cultured cells. Mutat. Res. 2016, 798, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Tiessen, C.; Ellmer, D.; Mikula, H.; Pahlke, G.; Warth, B.; Gehrke, H.; Marko, D. Impact of phase I metabolism on uptake, oxidative stress and genotoxicity of the emerging mycotoxin alternariol and its monomethyl ether in esophageal cells. Arch. Toxicol. 2017, 91, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, P.; Kashyap, P.L.; Goswami, S.K.; Chakdar, H.; Srivastava, A.K.; Saxena, A.K. Genome-wide analysis of microsatellites in Alternaria arborescens and elucidation of the function of Polyketide Synthase (PksJ). Interdiscip. Sci. 2018, 10, 813–822. [Google Scholar] [CrossRef]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemöller, M.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the IIα isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef]
- Somma, S.; Amatulli, M.T.; Masiello, M.; Moretti, A.; Logrieco, A.F. Alternaria species associated to wheat black point identified through a multilocus sequence approach. Int. J. Food Microbial. 2019, 293, 34–43. [Google Scholar] [CrossRef]
- Farhoosh, R.; Nyström, L. Antioxidant potency of gallic acid, methyl gallate and their combinations in sunflower oil triacylglycerols at high temperature. Food Chem. 2018, 244, 29–35. [Google Scholar] [CrossRef]
- Marino, T.; Galano, A.; Russo, N. Radical scavenging ability of gallic acid toward OH and OOH radicals. Reaction mechanism and rate constants from the density functional theory. J. Phys. Chem. B 2014, 118, 10380–10389. [Google Scholar] [CrossRef]
- Reckziegel, P.; Dias, V.T.; Benvegnú, D.M.; Boufleur, N.; Barcelos, R.C.S.; Segat, H.J.; Bürger, M.E. Antioxidant protection of gallic acid against toxicity induced by Pb in blood, liver and kidney of rats. Toxicol. Rep. 2016, 3, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Ghaznavi, H.; Fatemi, I.; Kalantari, H.; Hosseini Tabatabaei, S.M.T.; Mehrabani, M.; Gholamine, B.; Goudarzi, M. Ameliorative effects of gallic acid on gentamicin-induced nephrotoxicity in rats. J. Asian Nat. Prod. Res. 2018, 20, 1182–1193. [Google Scholar] [CrossRef]
- Ahn, C.B.; Jung, W.K.; Park, S.J.; Kim, Y.T.; Kim, W.S.; Je, J.Y. Gallic acid-g-Chitosan modulates inflammatory responses in LPS-stimulated RAW264.7 cells via NF-κB, AP-1, and MAPK pathways. Inflammation 2016, 39, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.P.; Jung, T.; Grune, T.; Siems, W. 4-Hydroxynonenal (HNE) modified proteins in metabolic diseases. Free Radic. Biol. Med. 2017, 111, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Sadhu, A.; Moriyasu, Y.; Acharya, K.; Bandyopadhyay, M. Nitric oxide and ROS mediate autophagy and regulate Alternaria alternata toxin-induced cell death in tobacco BY-2 cells. Sci. Rep. 2019, 9, 8973. [Google Scholar] [CrossRef] [PubMed]
- Akinrinde, A.S.; Adebiyi, O.E. Neuroprotection by luteolin and gallic acid against cobalt chloride-induced behavioural, morphological and neurochemical alterations in Wistar rats. Neurotoxicology 2019, 74, 252–263. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Area Percent (Mean ± SD) | ANOVA | LSD POST Hoc Test (Compared to Control Group) |
|---|---|---|---|
| Control group I | 4.69 ± 0.43 | 0.000 | |
| Alternariol group III | 15.52 ± 0.74 a | 0.000 | |
| Gallic + AOH group IV | 5.12 ± 0.62 b | 0.086 |
| Group | COMET % (Mean ± SD) | ANOVA | LSD POST Hoc Test (Compared to Control Group) |
|---|---|---|---|
| Control group I | 9.9 ± 2.7 | 0.000 | |
| Alternariol group III | 16.6 ± 5.3 a | 0.000 | |
| Gallic+ AOH group IV | 11.7 ± 4.7 b | 0.021 | |
| TAIL LENGTH (Mean ± SD) | |||
| Control group I | 5.68 ± 1.9 | 0.000 | |
| Alternariol group III | 3.84 ± 0.9 a | 0.011 | |
| Gallic+ AOH group IV | 5.13 ± 1.1 b | 0.045 | |
| TAIL DNA % (Mean ± SD) | |||
| Control group I | 9.51 ± 1.4 | 0.000 | |
| Alternariol group III | 10.58 ± 1.7 a | 0.034 | |
| Gallic+ AOH group IV | 9.65 ± 1.5 b | 0.142 | |
| TAIL MOMENT (Mean ± SD) | |||
| Control group I | 0.48 ± 0.09 | 0.000 | |
| Alternariol group III | 0.62 ± 0.13 a | 0.014 | |
| Gallic+ AOH group IV | 0.51 ± 0.11 b | 0.236 |
| Group | MDA (Mean ± SD) | ANOVA | LSD POST Hoc Test (Compared to Control Group) |
|---|---|---|---|
| Control group I | 9.1 ± 4.3 | 0.000 | |
| Alternariol group III | 19.3 ± 6.7 a | 0.000 | |
| Gallic + AOH group IV | 10.7 ± 5.4 b | 0.042 | |
| CAT (Mean ± SD) | |||
| Control group I | 0.49 ± 0.03 | 0.023 | |
| Alternariol group III | 0.31 ± 0.01 a | 0.012 | |
| Gallic+ AOH group IV | 0.47 ± 0.02 b | 0.134 | |
| SOD (Mean ± SD) | |||
| Control group I | 22.19 ± 1.12 | 0.000 | |
| Alternariol group III | 9.38 ± 4.59 a | 0.000 | |
| Gallic+ AOH group IV | 20.47 ± 1.19 b | 0.078 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samak, M.A.; Elshatory, A.; Mohamed, E.M. Outcomes of Gallic Acid on Alternariol Induced Cyto-Morphic and Genotoxic In Vivo Changes in Parotid Gland: 4-HNE Incorporated. Biomedicines 2019, 7, 84. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040084
Samak MA, Elshatory A, Mohamed EM. Outcomes of Gallic Acid on Alternariol Induced Cyto-Morphic and Genotoxic In Vivo Changes in Parotid Gland: 4-HNE Incorporated. Biomedicines. 2019; 7(4):84. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040084
Chicago/Turabian StyleSamak, Mai A., Ahmed Elshatory, and Eman M. Mohamed. 2019. "Outcomes of Gallic Acid on Alternariol Induced Cyto-Morphic and Genotoxic In Vivo Changes in Parotid Gland: 4-HNE Incorporated" Biomedicines 7, no. 4: 84. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040084