5-(Carbamoylmethylene)-oxazolidin-2-ones as a Promising Class of Heterocycles Inducing Apoptosis Triggered by Increased ROS Levels and Mitochondrial Dysfunction in Breast and Cervical Cancer


 , , , ,
, , , ,  and
and
Abstract
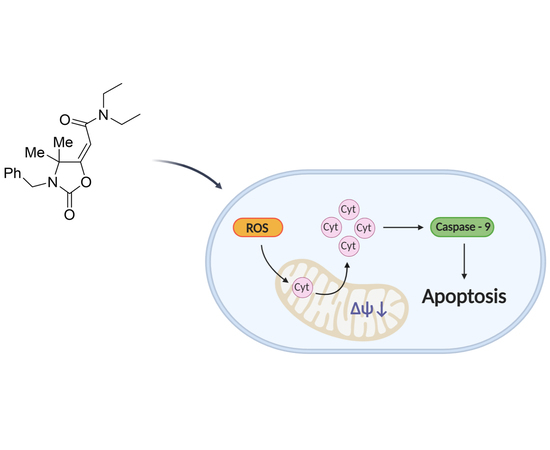
:
1. Introduction
2. Experimental Section
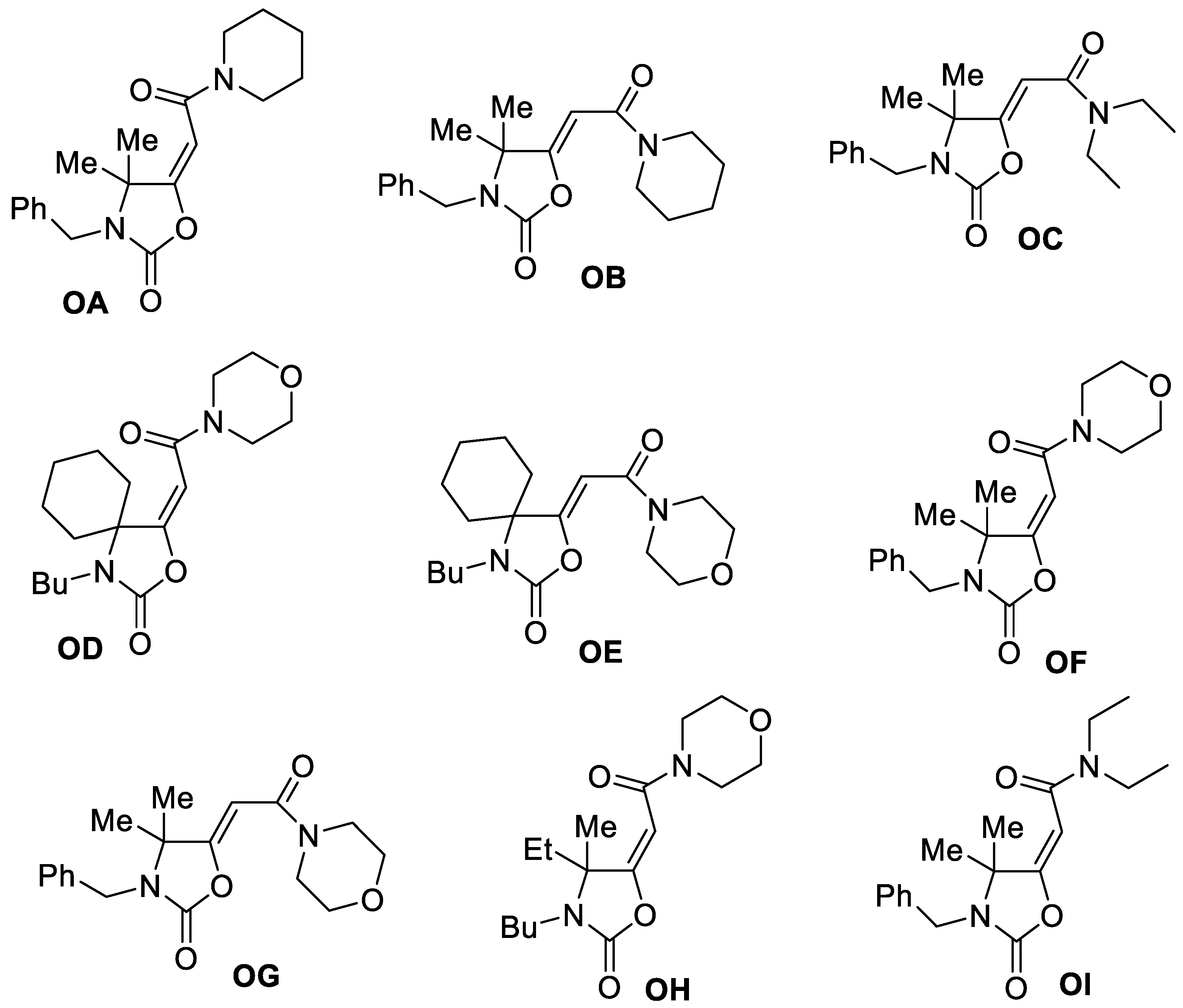
2.1. Synthesis of oxazolidinones OA-OI
2.2. Cell Cultures
2.3. Cell Viability Assay
2.4. Cell Cycle Analysis
2.5. Mitochondria Extraction
2.6. Western Blot Analysis
2.7. Mitochondrial Staining
2.8. ROS Production
2.9. Tunel Assay
2.10. Statistical Analysis
3. Results
3.1. Antiproliferative Activity
3.2. Oxazolidinone OI Induces Cell Cycle Arrest
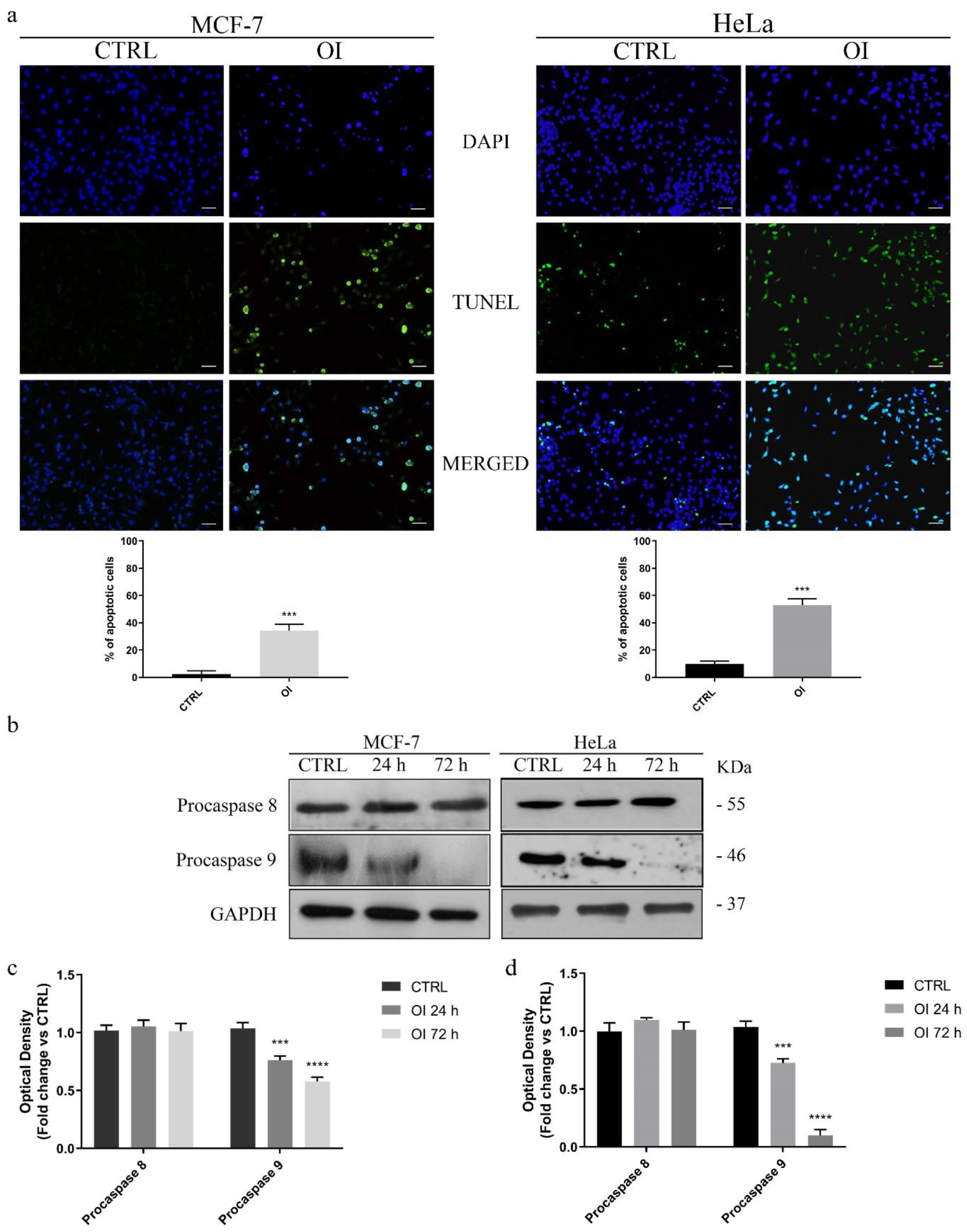
3.3. OI Triggers Apoptotic Cell Death in MCF-7 and HeLa cells
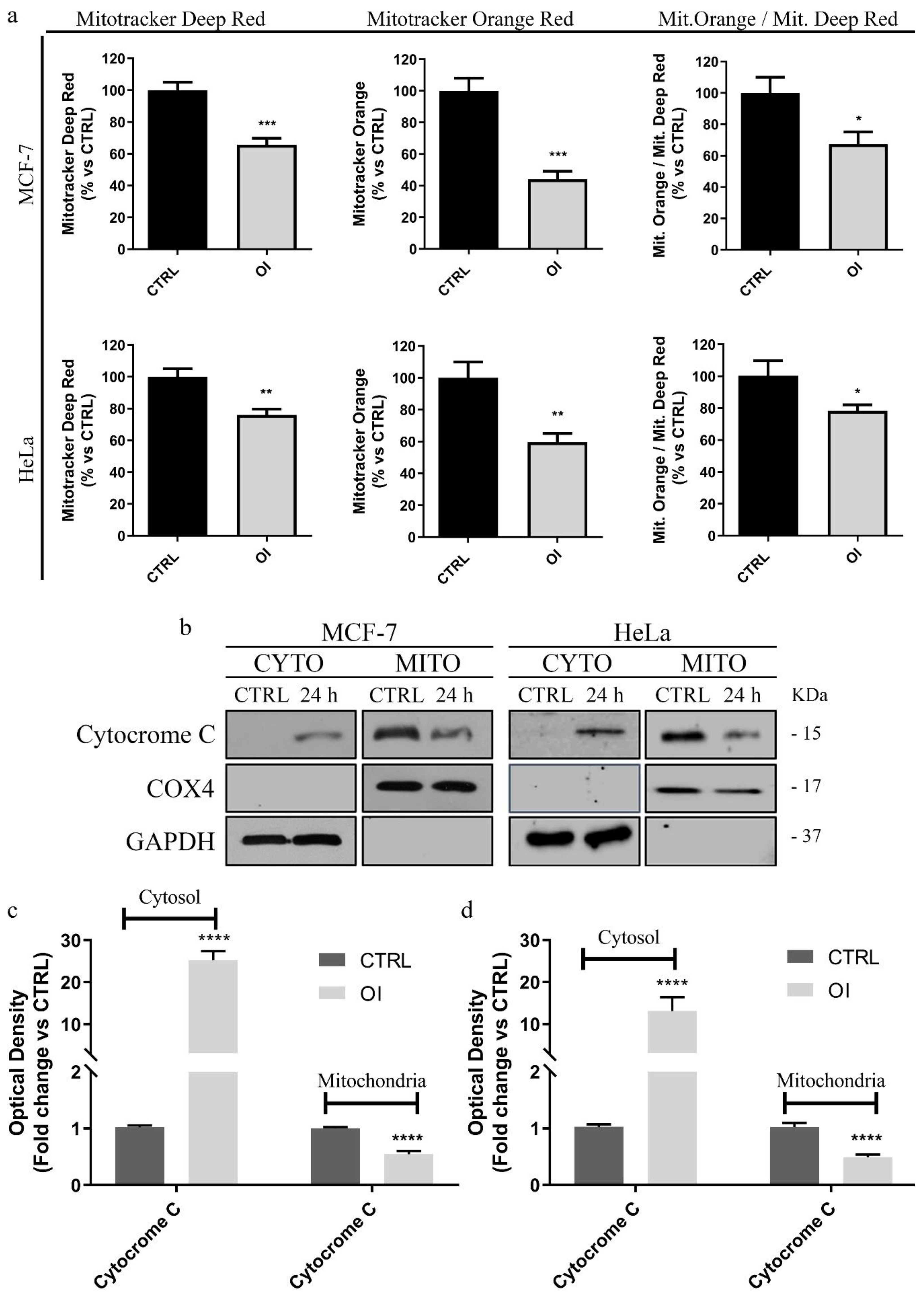
3.4. OI Reduces Mitochondrial Membrane Potential with a Significant Increase in ROS Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cancer Fact Sheet N 297. Available online: www.who.int/mediacentre/factsheets/fs297/en/ (accessed on 25 November 2019).
- Friberg, S.; Nystrom, A. Cancer Metastases: Early Dissemination and Late Recurrences. Cancer Growth Metastasis 2015, 8, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, A.J.; Munoz-Gomez, S.A.; Kamikawa, R. The Origin and Diversification of Mitochondria. Curr. Biol. 2017, 27, R1177–R1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, N.; Singla, R.K.; Shrivastava, B. Current updates on oxazolidinone and its significance. Int. J. Med. Chem. 2012, 2012, 159285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, D.L.; Dotter, B.; Madaras-Kelly, K. A review of linezolid: The first oxazolidinone antibiotic. Expert Rev. Anti-Infect. Ther. 2004, 2, 51–59. [Google Scholar] [CrossRef]
- Barrett, J.F. Linezolid Pharmacia Corp. Curr. Opin. Investig. Drugs 2000, 1, 181–187. [Google Scholar]
- Zhang, H.Z.; Zhao, Z.L.; Zhou, C.H. Recent advance in oxazole-based medicinal chemistry. Eur. J. Med. Chem. 2018, 144, 444–492. [Google Scholar] [CrossRef]
- Meka, V.G.; Gold, H.S. Antimicrobial resistance to linezolid. Clin. Infect. Dis. 2014, 39, 1010–1015. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Jiang, M.; Zhou, S.; Wu, S.; Zhang, X.; Ma, L.; Zhang, K.; Gong, P. Design, synthesis and structure–activity relationship of oxazolidinone derivatives containing novel S4 ligand as FXa inhibitors. Eur. J. Med. Chem. 2015, 96, 369–380. [Google Scholar] [CrossRef]
- Rapposelli, S.; Da Settimo, F.; Digiacomo, M.; La Motta, C.; Lapucci, A.; Sartini, S.; Vanni, M. Synthesis and biological evaluation of 2′-oxo-2,3-dihydro-3′H- spiro[chromene-4,5′-[1,3]oxazolidin]-3′yl]acetic acid derivatives as aldose reductase inhibitors. Arch. Der Pharm. 2011, 344, 372–385. [Google Scholar] [CrossRef]
- Madariaga, M.G.; Swindells, S.; McKee, E.E. Oxazolidinones and human immunodeficiency virus. Antimicrob. Agents Chemother. 2007, 51, 1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brnardic, E.J.; Fraley, M.E.; Garbaccio, R.M.; Layton, M.E.; Sanders, J.M.; Culberson, C.; Jacobson, M.A.; Magliaro, B.C.; Hutson, P.H.; O’Brien, J.A.; et al. 3-Aryl-5-phenoxymethyl-1,3-oxazolidin-2-ones as positive allosteric modulators of mGluR2 for the treatment of schizophrenia: Hit-to-lead efforts. Bioorg Med. Chem. Lett. 2010, 20, 3129–3133. [Google Scholar] [CrossRef] [PubMed]
- Welsch-Kunze, S.; Kuschinsky, K. On the possible involvement of glutamate receptors in conditioning of behavioural effects of apomorphine. Psychopharmacology 1990, 101, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.K.; Hop, C.E.; Pang, J.; Silva Elipe, M.V.; Desai, R.C.; Leung, K.H.; Franklin, R.B. In vitro metabolism of a new oxazolidinedione hypoglycemic agent utilizing liver microsomes and recombinant human cytochrome P450 enzymes. J. Pharm. Biomed. Anal. 2005, 37, 351–358. [Google Scholar] [CrossRef]
- Dow, R.L.; Bechle, B.M.; Chou, T.T.; Clark, D.A.; Hulin, B.; Stevenson, R.W. Benzyloxazolidine-2,4-Diones as Potent Hypoglycemic Agents. J Med. Chem. 1991, 34, 1538–1544. [Google Scholar] [CrossRef]
- Kombian, S.B.; Phillips, O.A. Novel Actions of Oxazolidinones: In vitro Screening of a Triazolyloxazolidinone for Anticonvulsant Activity. Med. Prin. Pr. 2013, 22, 340–345. [Google Scholar] [CrossRef]
- Naresh, A.; Venkateswara Rao, M.; Kotapalli, S.S.; Ummanni, R.; Venkateswara Rao, B. Oxazolidinone derivatives: Cytoxazone-linezolid hybrids induces apoptosis and senescence in DU145 prostate cancer cells. Eur. J. Med. Chem. 2014, 80, 295–307. [Google Scholar] [CrossRef]
- Tadesse, M.; Svenson, J.; Jaspars, M.; Strom, M.B.; Abdelrahman, M.H.; Andersen, J.H.; Hansen, E.; Kristiansen, P.E.; Stensvag, K.; Haug, T. Synoxazolidinone C; a bicyclic member of the synoxazolidinone family with antibacterial and anticancer activities. Tetrahedron Lett. 2011, 52, 1804–1806. [Google Scholar] [CrossRef]
- Singh, A.; Ha, H.J.; Park, J.; Kim, J.H.; Lee, W.K. 3,4-Disubstituted oxazolidin-2-ones as constrained ceramide analogs with anticancer activities. Bioorganic Med. Chem. 2011, 19, 6174–6181. [Google Scholar] [CrossRef]
- Winum, J.-Y.; Rami, M.; Scozzafava, A.; Montero, J.-L.; Supuran, C. Carbonic anhydrase IX: A new druggable target for the design of antitumor agents. Med. Res. Rev. 2008, 28, 445–463. [Google Scholar] [CrossRef]
- Pal, T.; Joshi, H.; Ramaa, C.S. Design and development of oxazol-5-ones as potential partial PPAR-gamma agonist against cancer cell lines. Anti-Cancer Agents Med. Chem. 2014, 14, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.F.; Pereira, M.C.; de Sena, W.L.B.; de Barros Martins, C.G.; de Oliveira, J.F.; da Cruz Amorim, C.A.; de Melo Rego, M.J.B.; da Rocha Pitta, M.G.; de Lima, M.; da Rocha Pitta, M.G.; et al. Synthesis and in vitro anticancer activity of new 2-thioxo-oxazolidin-4-one derivatives. Pharmacol. Rep. 2017, 69, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pang, F.H.; Huang, L.; Yang, X.P.; Ma, X.L.; Jiang, C.N.; Li, F.Y.; Lei, F.H. Synthesis and Biological Evaluation of Novel Dehydroabietic Acid-Oxazolidinone Hybrids for Antitumor Properties. Int. J. Mol. Sci. 2018, 19, 3116. [Google Scholar] [CrossRef] [Green Version]
- Hedaya, O.M.; Mathew, P.M.; Mohamed, F.H.; Phillips, O.A.; Luqmani, Y.A. Antiproliferative activity of a series of 5(1H1,2,3triazolyl) methyl and 5acetamidomethyloxazolidinone derivatives. Mol. Med. Rep. 2016, 13, 3311–3318. [Google Scholar] [CrossRef] [Green Version]
- Giofre, S.V.; Cirmi, S.; Mancuso, R.; Nicolo, F.; Lanza, G.; Legnani, L.; Campisi, A.; Chiacchio, M.A.; Navarra, M.; Gabriele, B.; et al. Synthesis of spiro[isoindole-1,5′-isoxazolidin]-3(2H)-ones as potential inhibitors of the MDM2-p53 interaction. Beilstein. J. Org. Chem. 2016, 12, 2793–2807. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, R.; Maner, A.; Ziccarelli, I.; Pomelli, C.; Chiappe, C.; Della Ca, N.; Veltri, L.; Gabriele, B. Auto-Tandem Catalysis in Ionic Liquids: Synthesis of 2-Oxazolidinones by Palladium-Catalyzed Oxidative Carbonylation of Propargylic Amines in EmimEtSO(4). Molecules 2016, 21, 897. [Google Scholar] [CrossRef] [Green Version]
- Gabriele, B.; Plastina, P.; Salerno, G.; Mancuso, R.; Costa, M. An unprecedented Pd-catalyzed, water-promoted sequential oxidative aminocarbonylation-cyclocarbonylation process leading to 2-oxazolidinones. Org. Lett. 2007, 9, 3319–3322. [Google Scholar] [CrossRef]
- Dhanya, D.; Palma, G.; Cappello, A.R.; Mariconda, A.; Sinicropi, M.S.; Francesca, G.; Del Vecchio, V.; Ramunno, A.; Claudio, A.; Pasquale, L.; et al. Phosphonium Salt Displays Cytotoxic Effects Against Human Cancer Cell Lines. Anti-Cancer Agents Med. Chem. 2017, 17, 1796–1804. [Google Scholar] [CrossRef]
- Wang, S.C.; Chow, J.M.; Chien, M.H.; Lin, C.W.; Chen, H.Y.; Hsiao, P.C.; Yang, S.F. Cantharidic acid induces apoptosis of human leukemic HL-60 cells via c-Jun N-terminal kinase-regulated caspase-8/-9/-3 activation pathway. Environ. Toxicol. 2018, 33, 514–522. [Google Scholar] [CrossRef]
- Bonesi, M.; Brindisi, M.; Armentano, B.; Curcio, R.; Sicari, V.; Loizzo, M.R.; Cappello, M.S.; Bedini, G.; Peruzzi, L.; Tundis, R. Exploring the anti-proliferative, pro-apoptotic, and antioxidant properties of Santolina corsica Jord. & Fourr. (Asteraceae). Biomed. Pharmacother. Biomed. Pharmacother. 2018, 107, 967–978. [Google Scholar] [CrossRef]
- Frattaruolo, L.; Lacret, R.; Cappello, A.R.; Truman, A.W. A Genomics-Based Approach Identifies a Thioviridamide-Like Compound with Selective Anticancer Activity. ACS Chem. Biol. 2017, 12, 2815–2822. [Google Scholar] [CrossRef] [PubMed]
- Cappello, A.R.; Aiello, F.; Polera, N.; Armentano, B.; Casaburi, I.; Di Gioia, M.L.; Loizzo, M.R.; Dolce, V.; Pezzi, V.; Tundis, R. In vitro anti-proliferative and anti-bacterial properties of new C7 benzoate derivatives of pinocembrin. Nat. Prod. Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, I.; Avena, P.; De Luca, A.; Chimento, A.; Sirianni, R.; Malivindi, R.; Rago, V.; Fiorillo, M.; Domanico, F.; Campana, C.; et al. Estrogen related receptor alpha (ERRalpha) a promising target for the therapy of adrenocortical carcinoma (ACC). Oncotarget 2015, 6, 25135–25148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frattaruolo, L.; Fiorillo, M.; Brindisi, M.; Curcio, R.; Dolce, V.; Lacret, R.; Truman, A.W.; Sotgia, F.; Lisanti, M.P.; Cappello, A.R. Thioalbamide, A Thioamidated Peptide from Amycolatopsis alba, Affects Tumor Growth and Stemness by Inducing Metabolic Dysfunction and Oxidative Stress. Cells 2019, 8, 1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frattaruolo, L.; Carullo, G.; Brindisi, M.; Mazzotta, S.; Bellissimo, L.; Rago, V.; Curcio, R.; Dolce, V.; Aiello, F.; Cappello, A.R. Antioxidant and Anti-Inflammatory Activities of Flavanones from Glycyrrhiza glabra L. (licorice) Leaf Phytocomplexes: Identification of Licoflavanone as a Modulator of NF-kB/MAPK Pathway. Antioxidants 2019, 8, 186. [Google Scholar] [CrossRef] [Green Version]
- Curcio, R.; Aiello, D.; Vozza, A.; Muto, L.; Martello, E.; Cappello, A.R.; Capobianco, L.; Fiermonte, G.; Siciliano, C.; Napoli, A.; et al. Cloning, Purification, and Characterization of the Catalytic C-Terminal Domain of the Human 3-Hydroxy-3-methyl glutaryl-CoA Reductase: An Effective, Fast, and Easy Method for Testing Hypocholesterolemic Compounds. Mol. Biotechnol. 2020, 62, 119–131. [Google Scholar] [CrossRef]
- Li, Y.; Cappello, A.R.; Muto, L.; Martello, E.; Madeo, M.; Curcio, R.; Lunetti, P.; Raho, S.; Zaffino, F.; Frattaruolo, L.; et al. Functional characterization of the partially purified Sac1p independent adenine nucleotide transport system (ANTS) from yeast endoplasmic reticulum. J. Biochem. 2018, 164, 313–322. [Google Scholar] [CrossRef]
- Fiorillo, M.; Lamb, R.; Tanowitz, H.B.; Cappello, A.R.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Bedaquiline, an FDA-approved antibiotic, inhibits mitochondrial function and potently blocks the proliferative expansion of stem-like cancer cells (CSCs). Aging 2016, 8, 1593–1607. [Google Scholar] [CrossRef] [Green Version]
- Iacopetta, D.; Carocci, A.; Sinicropi, M.S.; Catalano, A.; Lentini, G.; Ceramella, J.; Curcio, R.; Caroleo, M.C. Old Drug Scaffold, New Activity: Thalidomide-Correlated Compounds Exert Different Effects on Breast Cancer Cell Growth and Progression. ChemMedChem 2017, 12, 381–389. [Google Scholar] [CrossRef]
- Kroemer, G. Mitochondrial control of apoptosis: An overview. Biochem. Soc. Symp. 1999, 66, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Buttke, T.M.; Sandstrom, P.A. Oxidative stress as a mediator of apoptosis. Immunol. Today 1994, 15, 7–10. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Dang, L.; White, D.W.; Gross, S.; Bennett, B.D.; Bittinger, M.A.; Driggers, E.M.; Fantin, V.R.; Jang, H.G.; Jin, S.; Keenan, M.C.; et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009, 462, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fendt, S.M.; Bell, E.L.; Keibler, M.A.; Olenchock, B.A.; Mayers, J.R.; Wasylenko, T.M.; Vokes, N.I.; Guarente, L.; Vander Heiden, M.G.; Stephanopoulos, G. Reductive glutamine metabolism is a function of the alpha-ketoglutarate to citrate ratio in cells. Nat. Commun. 2013, 4, 2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, D.R.; Ward, P.S.; Shay, J.E.; Cross, J.R.; Gruber, J.J.; Sachdeva, U.M.; Platt, J.M.; DeMatteo, R.G.; Simon, M.C.; Thompson, C.B. Hypoxia promotes isocitrate dehydrogenase-dependent carboxylation of alpha-ketoglutarate to citrate to support cell growth and viability. Proc. Natl. Acad. Sci. USA 2011, 108, 19611–19616. [Google Scholar] [CrossRef] [Green Version]
- Zamzami, N.; Marchetti, P.; Castedo, M.; Zanin, C.; Vayssiere, J.L.; Petit, P.X.; Kroemer, G. Reduction in mitochondrial potential constitutes an early irreversible step of programmed lymphocyte death in vivo. J. Exp. Med. 1995, 181, 1661–1672. [Google Scholar] [CrossRef] [Green Version]
- Zamzami, N.; Marchetti, P.; Castedo, M.; Decaudin, D.; Macho, A.; Hirsch, T.; Susin, S.A.; Petit, P.X.; Mignotte, B.; Kroemer, G. Sequential reduction of mitochondrial transmembrane potential and generation of reactive oxygen species in early programmed cell death. J. Exp. Med. 1995, 182, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Zamzami, N.; Susin, S.A.; Marchetti, P.; Hirsch, T.; Gomez-Monterrey, I.; Castedo, M.; Kroemer, G. Mitochondrial control of nuclear apoptosis. J. Exp. Med. 1996, 183, 1533–1544. [Google Scholar] [CrossRef]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- De Luca, A.; Fiorillo, M.; Peiris-Pages, M.; Ozsvari, B.; Smith, D.L.; Sanchez-Alvarez, R.; Martinez-Outschoorn, U.E.; Cappello, A.R.; Pezzi, V.; Lisanti, M.P.; et al. Mitochondrial biogenesis is required for the anchorage-independent survival and propagation of stem-like cancer cells. Oncotarget 2015, 6, 14777–14795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fillmore, C.M.; Kuperwasser, C. Human breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapy. Breast Cancer Res. 2008, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Rosen, J.M. Stem cells in the etiology and treatment of cancer. Curr. Opin. Genet. Dev. 2006, 16, 60–64. [Google Scholar] [CrossRef]
- Li, X.; Lewis, M.T.; Huang, J.; Gutierrez, C.; Osborne, C.K.; Wu, M.F.; Hilsenbeck, S.G.; Pavlick, A.; Zhang, X.; Chamness, G.C.; et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J. Natl. Cancer Inst. 2008, 100, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Scopelliti, A.; Cammareri, P.; Catalano, V.; Saladino, V.; Todaro, M.; Stassi, G. Therapeutic implications of Cancer Initiating Cells. Expert Opin. Biol. Ther. 2009, 9, 1005–1016. [Google Scholar] [CrossRef]
- Easwaran, H.; Tsai, H.C.; Baylin, S.B. Cancer epigenetics: Tumor heterogeneity, plasticity of stem-like states, and drug resistance. Mol. Cell 2014, 54, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Fulda, S.; Galluzzi, L.; Kroemer, G. Targeting mitochondria for cancer therapy. Nat. Rev. Drug Discov. 2010, 9, 447–464. [Google Scholar] [CrossRef]
- Martinez-Outschoorn, U.E.; Peiris-Pages, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 113. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Kepp, O.; Vander Heiden, M.G.; Kroemer, G. Metabolic targets for cancer therapy. Nat. Rev. Drug Discov. 2013, 12, 829–846. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, D.L.; Sage, J. Cellular mechanisms of tumour suppression by the retinoblastoma gene. Nat. Rev. Cancer 2008, 8, 671–682. [Google Scholar] [CrossRef]
- Lindqvist, A.; Rodriguez-Bravo, V.; Medema, R.H. The decision to enter mitosis: Feedback and redundancy in the mitotic entry network. J. Cell Biol. 2009, 185, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Armentano, M.F.; Bisaccia, F.; Miglionico, R.; Russo, D.; Nolfi, N.; Carmosino, M.; Andrade, P.B.; Valentao, P.; Diop, M.S.; Milella, L. Antioxidant and proapoptotic activities of Sclerocarya birrea [(A. Rich.) Hochst.] methanolic root extract on the hepatocellular carcinoma cell line HepG2. Biomed Res. Int. 2015, 2015, 561589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, M.C.; Porporato, P.E.; Martini, M.; Morandi, A. Signaling Pathways Regulating Redox Balance in Cancer Metabolism. Front. Oncol. 2018, 8, 126. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | HeLa | MCF-7 | MCF-10A | |||
|---|---|---|---|---|---|---|
| IC50 | 95% Confidence Interval | IC50 | 95% Confidence Interval | IC50 | 95% Confidence Interval | |
| DOX | 3.61 | 3.02 to 4.32 | 4.04 | 3.25 to 5.03 | 31.75 | 25.16 to 40.08 |
| OA | 52.53 | 42.81 to 64.45 | 56.90 | 50.43 to 69.18 | 414.3 | 336.3 to 510.5 |
| OB | 98.21 | 81.46 to 118,0 | 121.8 | 107.0 to 139.7 | 469.6 | 378.9 to 582.1 |
| OC | 99.47 | 88.42 to 111.9 | 163.3 | 135.2 to197.2 | 494.5 | 406.2 to 602.2 |
| OD | 154.5 | 136.4 to 175.1 | 338.4 | 284.8 to 402.0 | 506.4 | 410.1 to 625.4 |
| OE | 129.3 | 114.0 to 146.0 | 303.7 | 258.3 to 357.0 | 978.2 | 718.3 to 1332 |
| OF | 159.4 | 138.5 to 183.6 | 409.3 | 348.8 to 480.3 | 718.4 | 506.6 to 1019 |
| OG | 134.4 | 113.5 to 159.2 | 344.9 | 290.4 409.6 | 677.1 | 530.3 to 864.5 |
| OH | 54.21 | 42.67 to 68.88 | 92.91 | 78.99 to 109.3 | 318.8 | 263.1 to 386.3 |
| OI | 31.10 | 26.29 to36.80 | 17.66 | 15.67 to 19.91 | 351.3 | 288.4 to 427.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armentano, B.; Curcio, R.; Brindisi, M.; Mancuso, R.; Rago, V.; Ziccarelli, I.; Frattaruolo, L.; Fiorillo, M.; Dolce, V.; Gabriele, B.; et al. 5-(Carbamoylmethylene)-oxazolidin-2-ones as a Promising Class of Heterocycles Inducing Apoptosis Triggered by Increased ROS Levels and Mitochondrial Dysfunction in Breast and Cervical Cancer. Biomedicines 2020, 8, 35. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8020035
Armentano B, Curcio R, Brindisi M, Mancuso R, Rago V, Ziccarelli I, Frattaruolo L, Fiorillo M, Dolce V, Gabriele B, et al. 5-(Carbamoylmethylene)-oxazolidin-2-ones as a Promising Class of Heterocycles Inducing Apoptosis Triggered by Increased ROS Levels and Mitochondrial Dysfunction in Breast and Cervical Cancer. Biomedicines. 2020; 8(2):35. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8020035
Chicago/Turabian StyleArmentano, Biagio, Rosita Curcio, Matteo Brindisi, Raffaella Mancuso, Vittoria Rago, Ida Ziccarelli, Luca Frattaruolo, Marco Fiorillo, Vincenza Dolce, Bartolo Gabriele, and et al. 2020. "5-(Carbamoylmethylene)-oxazolidin-2-ones as a Promising Class of Heterocycles Inducing Apoptosis Triggered by Increased ROS Levels and Mitochondrial Dysfunction in Breast and Cervical Cancer" Biomedicines 8, no. 2: 35. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8020035