Ectopic Expression of Hematopoietic SHIP1 in Human Colorectal Cancer

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Cell Culture
2.2. Protein Lysates and Western Blotting
2.3. Multi-Tumor Tissue Microarray
2.4. Immunohistochemistry
2.5. Bioinformatical Analysis of TCGA Data
2.6. Phosphatase Assay
3. Results
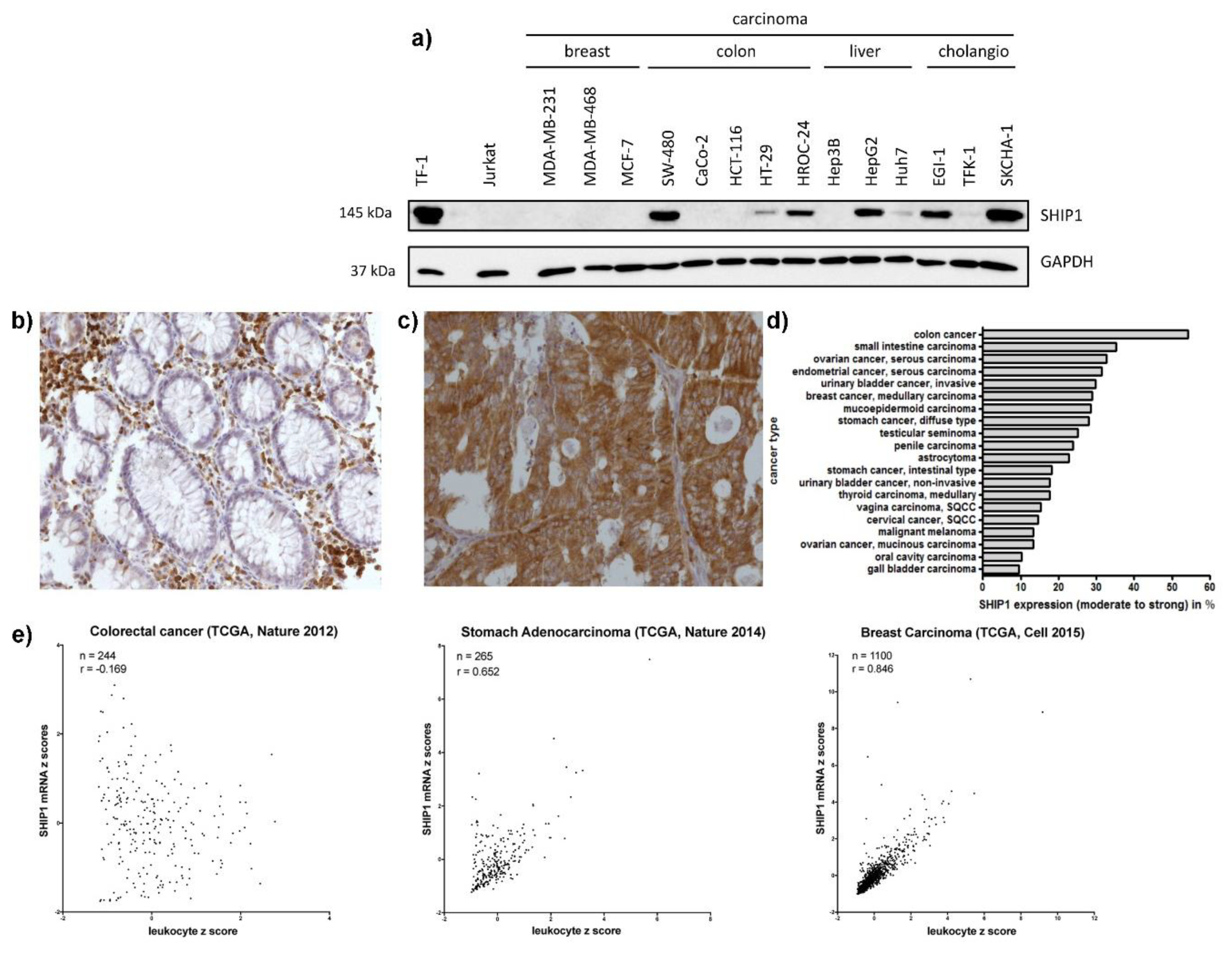
3.1. SHIP1 Expression in Human Carcinoma Cell Lines and Clinical Samples
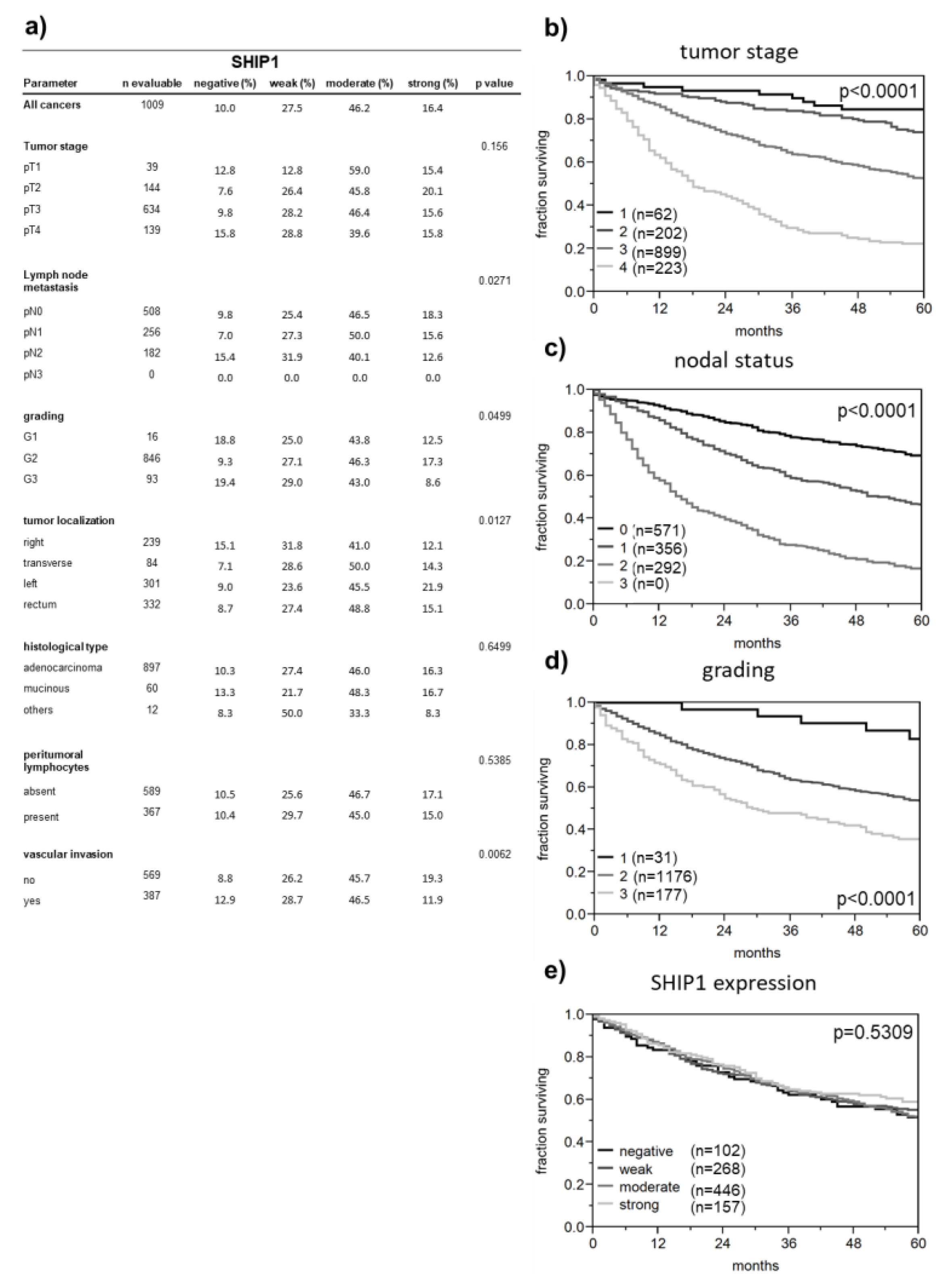
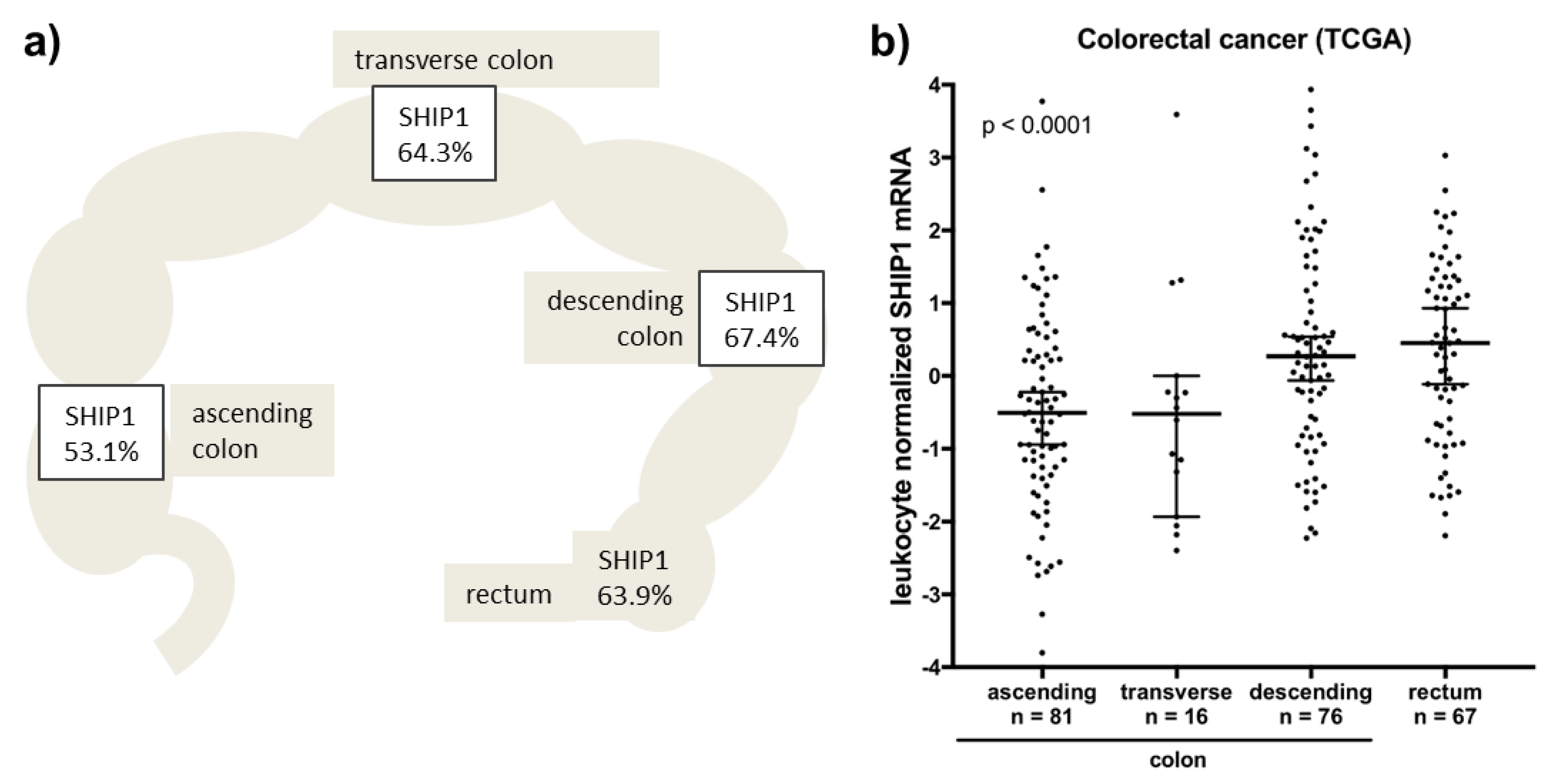
3.2. Correlation between SHIP1 Expression and Clinicopathological Parameters
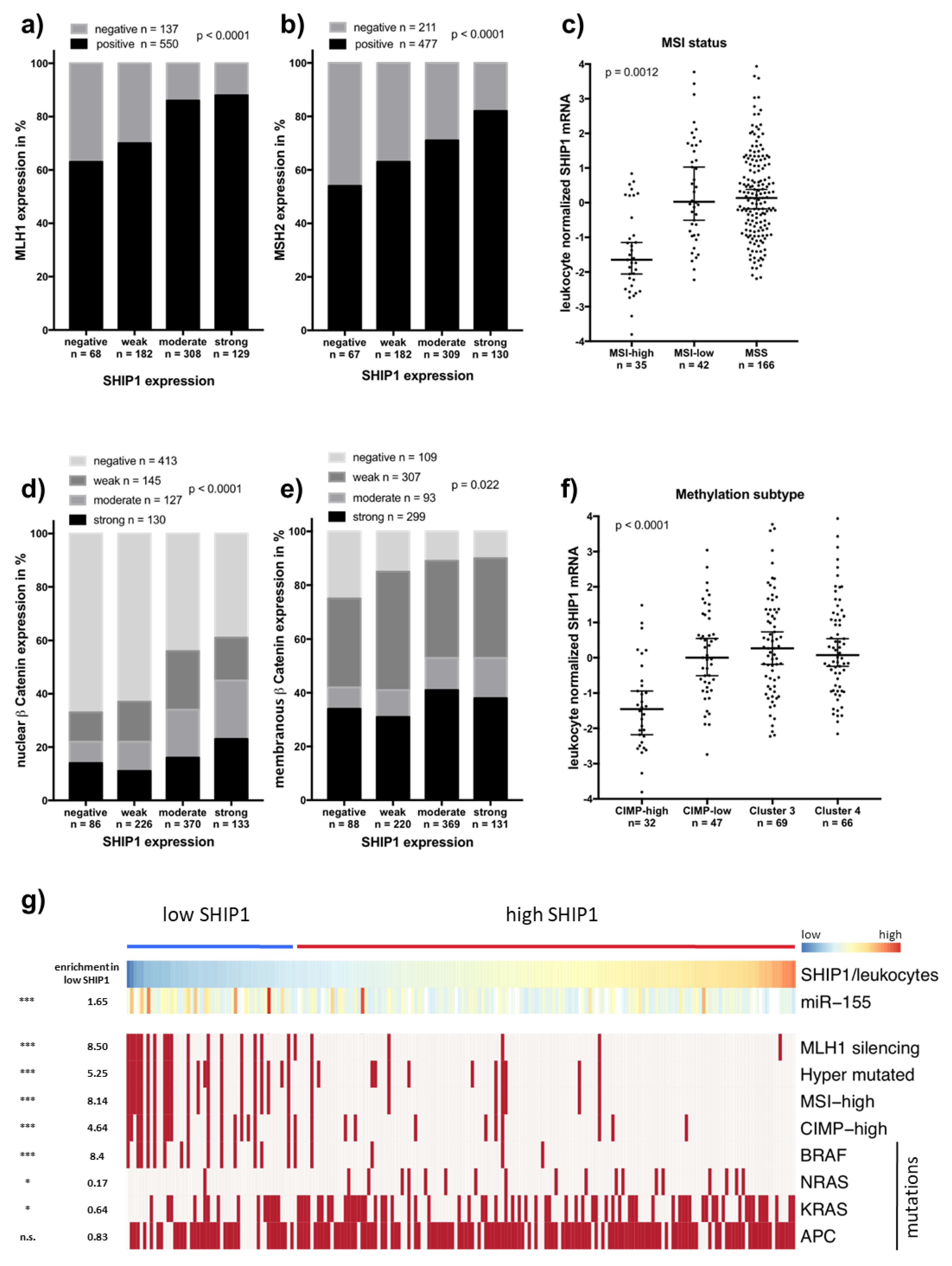
3.3. Downregulation of SHIP1 in Microsatellite Unstable Colorectal Cancers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, N.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Sinicrope, F.A.; Sargent, D.J. Molecular Pathways: Microsatellite Instability in Colorectal Cancer: Prognostic, Predictive, and Therapeutic Implications. Clin. Cancer Res. 2012, 18, 1506–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, C.R.; Goel, A. Microsatellite Instability in Colorectal Cancer. Gastroenterology 2010, 138, 2073–2087.e3. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Fu, M.; Ding, Y.; Weng, Y.; Fan, W.; Pu, X.; Ge, Z.; Zhan, F.; Ni, H.; Zhang, W.; et al. High SHIP2 expression indicates poor survival in colorectal cancer. Dis. Markers 2014, 2014, 218968. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.P.; Eddy, R.; Entenberg, D.; Kai, M.; Gertler, F.B.; Condeelis, J. Tks5 and SHIP2 regulate invadopodium maturation, but not initiation, in breast carcinoma cells. Curr. Biol. 2013, 23, 2079–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Chetrit, N.; Chetrit, D.; Russell, R.; Körner, C.; Mancini, M.; Abdul-Hai, A.; Itkin, T.; Carvalho, S.; Cohen-Dvashi, H.; Koestler, W.J.; et al. Synaptojanin 2 is a druggable mediator of metastasis and the gene is overexpressed and amplified in breast cancer. Sci. Signal. 2015, 8, ra7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauer, H.; Strauss, J.; Wegner, W.; Müller-Tidow, C.; Horstmann, M.; Jücker, M. Leukemia-associated mutations in SHIP1 inhibit its enzymatic activity, interaction with the GM-CSF receptor and Grb2, and its ability to inactivate PI3K/AKT signaling. Cell. Signal. 2012, 24, 2095–2101. [Google Scholar] [CrossRef] [PubMed]
- Gilby, D.C.; Goodeve, A.C.; Winship, P.R.; Valk, P.J.; Delwel, R.; Reilly, J.T. Gene structure, expression profiling and mutation analysis of the tumour suppressor SHIP1 in Caucasian acute myeloid leukaemia. Leukemia 2007, 21, 2390–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, S.; Endl, E.; Fehse, B.; Weck, M.M.; Mayr, G.W.; Jücker, M. Restoration of SHIP activity in a human leukemia cell line downregulates constitutively activated phosphatidylinositol 3-kinase/Akt/GSK-3β signaling and leads to an increased transit time through the G1 phase of the cell cycle. Leukemia 2004, 18, 1839–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Oliveira-Dos-Santos, A.J.; Mariathasan, S.; Bouchard, D.; Jones, J.; Sarao, R.; Kozieradzki, I.; Ohashi, P.S.; Penninger, J.M.; Dumont, D.J. The inositol polyphosphate 5-phosphatase ship is a crucial negative regulator of B cell antigen receptor signaling. J. Exp. Med. 1998, 188, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-M.; Yoshida, H.; Komura, S.; Ohishi, N.; Pan, L.; Shigeno, K.; Hanamura, I.; Miura, K.; Iida, S.; Ueda, R.; et al. Possible dominant-negative mutation of the SHIP gene in acute myeloid leukemia. Leukemia 2003, 17, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.-M.; Liu, Z.-L.; Hao, H.-L.; Wang, F.-X.; Dong, Z.-R.; Ohno, R. Mutation analysis of SHIP gene in acute leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2004, 12, 420–426. [Google Scholar] [PubMed]
- Täger, M.; Horn, S.; Latuske, E.; Ehm, P.; Schaks, M.; Nalaskowski, M.; Fehse, B.; Fiedler, W.; Stocking, C.; Wellbrock, J.; et al. SHIP1, but not an AML-derived SHIP1 mutant, suppresses myeloid leukemia growth in a xenotransplantation mouse model. Gene Ther. 2017, 24, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Grabinski, N.; Bartkowiak, K.; Grupp, K.; Brandt, B.; Pantel, K.; Jücker, M. Distinct functional roles of Akt isoforms for proliferation, survival, migration and EGF-mediated signalling in lung cancer derived disseminated tumor cells. Cell. Signal. 2011, 23, 1952–1960. [Google Scholar] [CrossRef] [PubMed]
- Tennstedt, P.; Fresow, R.; Simon, R.; Marx, A.H.; Terracciano, L.; Petersen, C.; Sauter, G.; Dikomey, E.; Borgmann, K. RAD51 overexpression is a negative prognostic marker for colorectal adenocarcinoma. Int. J. Cancer 2013, 132, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Panarelli, N.C.; Vaughn, C.P.; Samowitz, W.S.; Yantiss, R.K. Sporadic Microsatellite Instability-High Colon Cancers Rarely Display Immunohistochemical Evidence of Wnt Signaling Activation. Am. J. Surg. Pathol. 2015, 39, 313–317. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Chaudhuri, A.A.; Rao, D.S.; Baltimore, D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc. Natl. Acad. Sci. USA 2009, 106, 7113–7118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, T.F.; Kaplan, D.R.; Cantley, L.C.; Toker, A. Direct regulation of the Akt proto-oncogene product by phosphatidylinositol-3,4-bisphosphate. Science 1997, 275, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Scheid, M.P.; Huber, M.; Damen, J.E.; Hughes, M.; Kang, V.; Neilsen, P.; Prestwich, G.D.; Krystal, G.; Duronio, V. Phosphatidylinositol (3,4,5)P 3 Is Essential but Not Sufficient for Protein Kinase B (PKB) Activation; Phosphatidylinositol (3,4)P 2 Is Required for PKB Phosphorylation at Ser-473. J. Biol. Chem. 2002, 277, 9027–9035. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaks, M.; Allgoewer, K.; Nelson, N.; Ehm, P.; Heumann, A.; Ewald, F.; Schumacher, U.; Simon, R.; Sauter, G.; Jücker, M. Ectopic Expression of Hematopoietic SHIP1 in Human Colorectal Cancer. Biomedicines 2020, 8, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070215
Schaks M, Allgoewer K, Nelson N, Ehm P, Heumann A, Ewald F, Schumacher U, Simon R, Sauter G, Jücker M. Ectopic Expression of Hematopoietic SHIP1 in Human Colorectal Cancer. Biomedicines. 2020; 8(7):215. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070215
Chicago/Turabian StyleSchaks, Matthias, Kristina Allgoewer, Nina Nelson, Patrick Ehm, Asmus Heumann, Florian Ewald, Udo Schumacher, Ronald Simon, Guido Sauter, and Manfred Jücker. 2020. "Ectopic Expression of Hematopoietic SHIP1 in Human Colorectal Cancer" Biomedicines 8, no. 7: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070215