Method Standardization for Conducting Innate Color Preference Studies in Different Zebrafish Strains


 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Animal Ethics and Animal Used in the Study
2.2. Color Preference Assay
2.3. Data Analysis
2.4. Statistics
3. Results
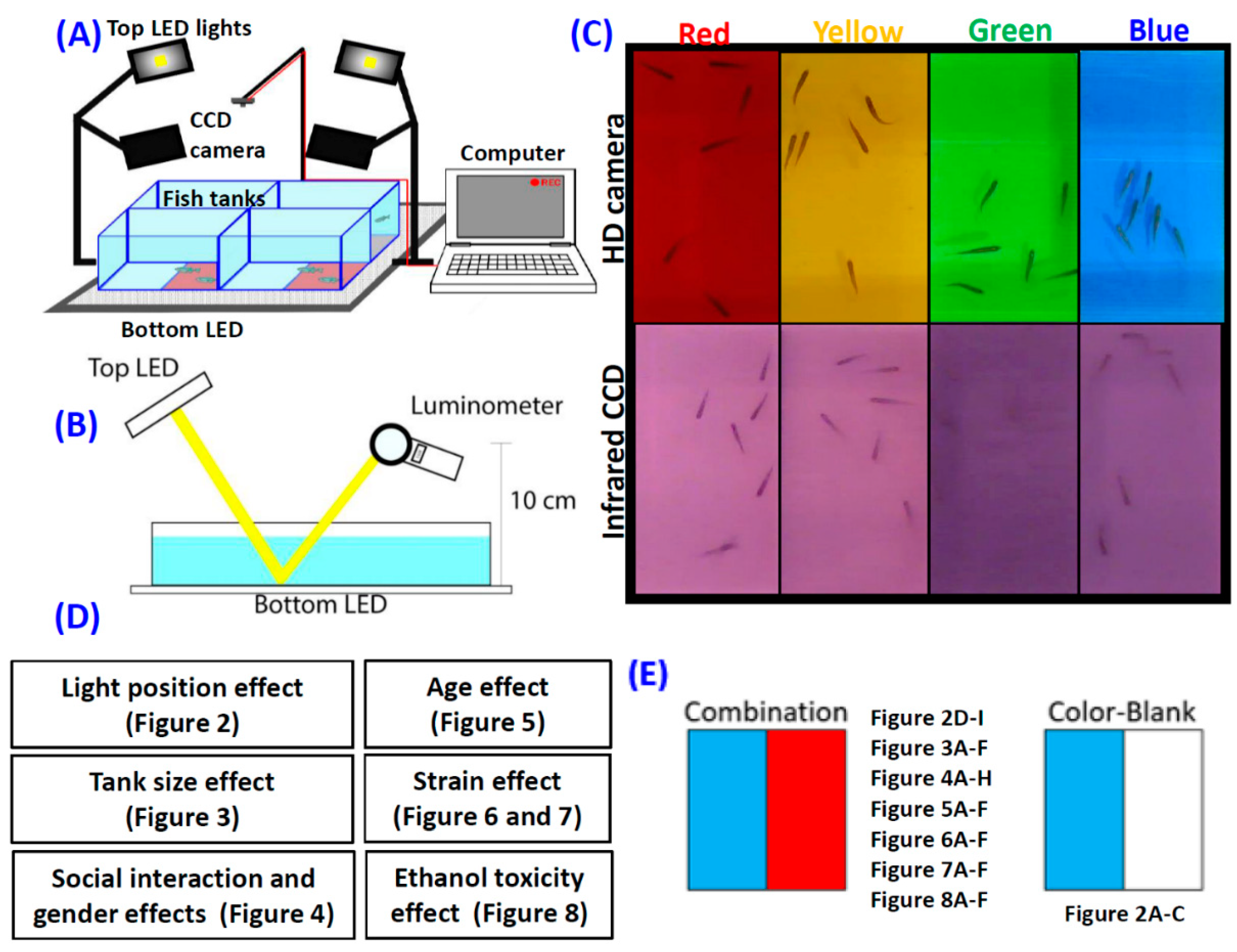
3.1. Overview of Experimental Design and Instrument Setting
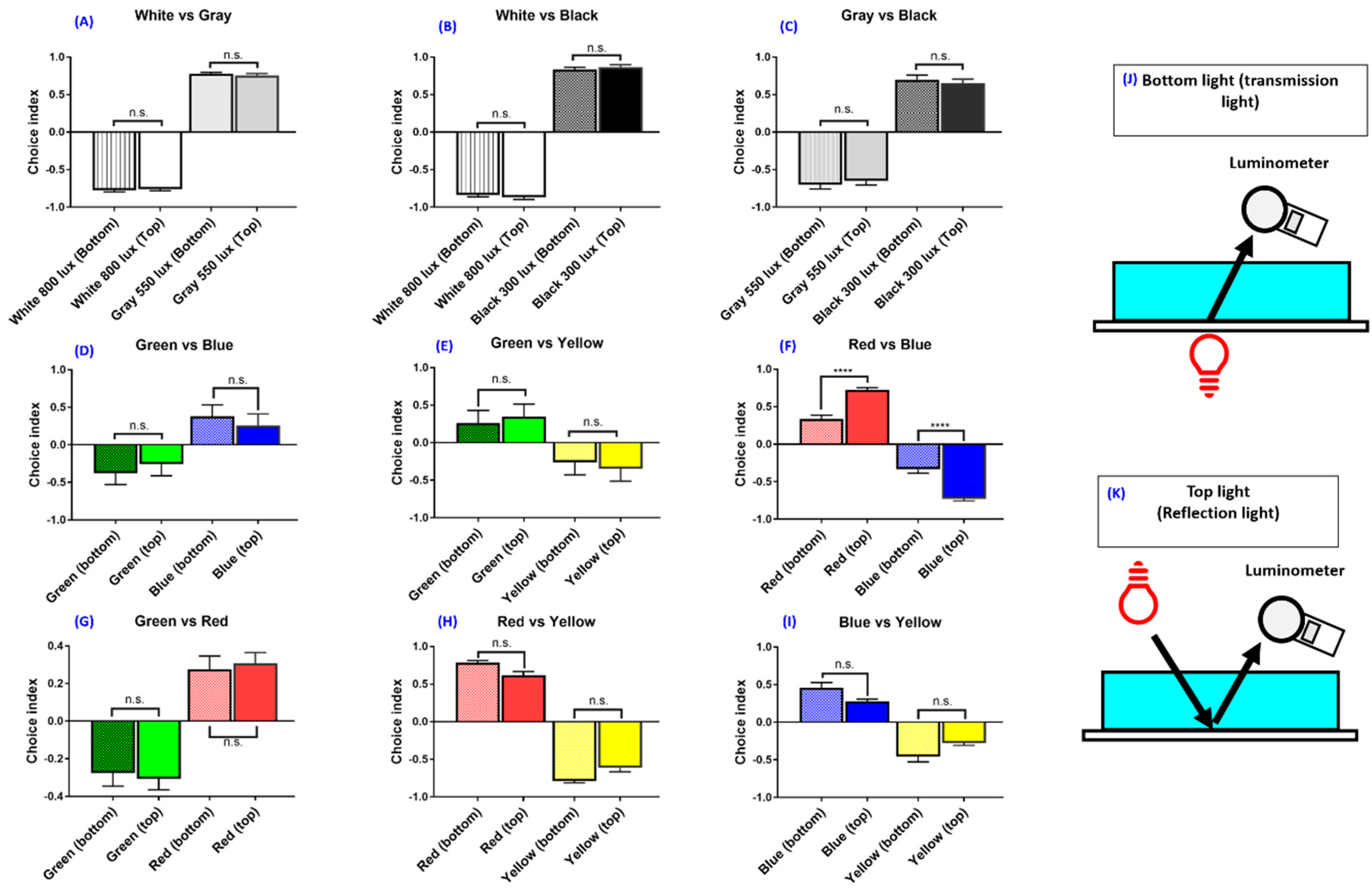
3.2. Experiment 1: Light Source Position on Zebrafish Color Preferences
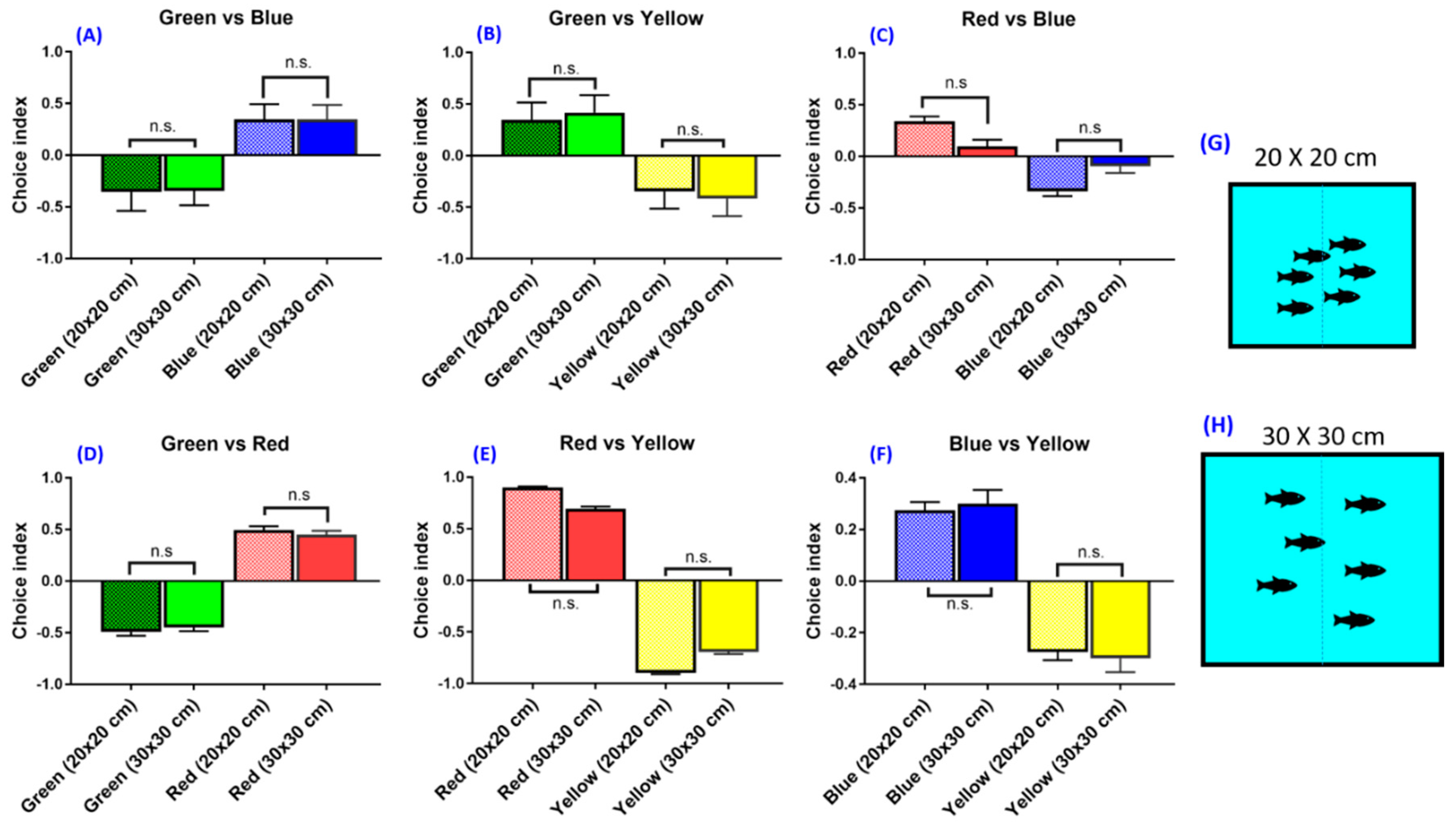
3.3. Experiment 2: Tank Size (Animal Density) on Zebrafish Color Preferences
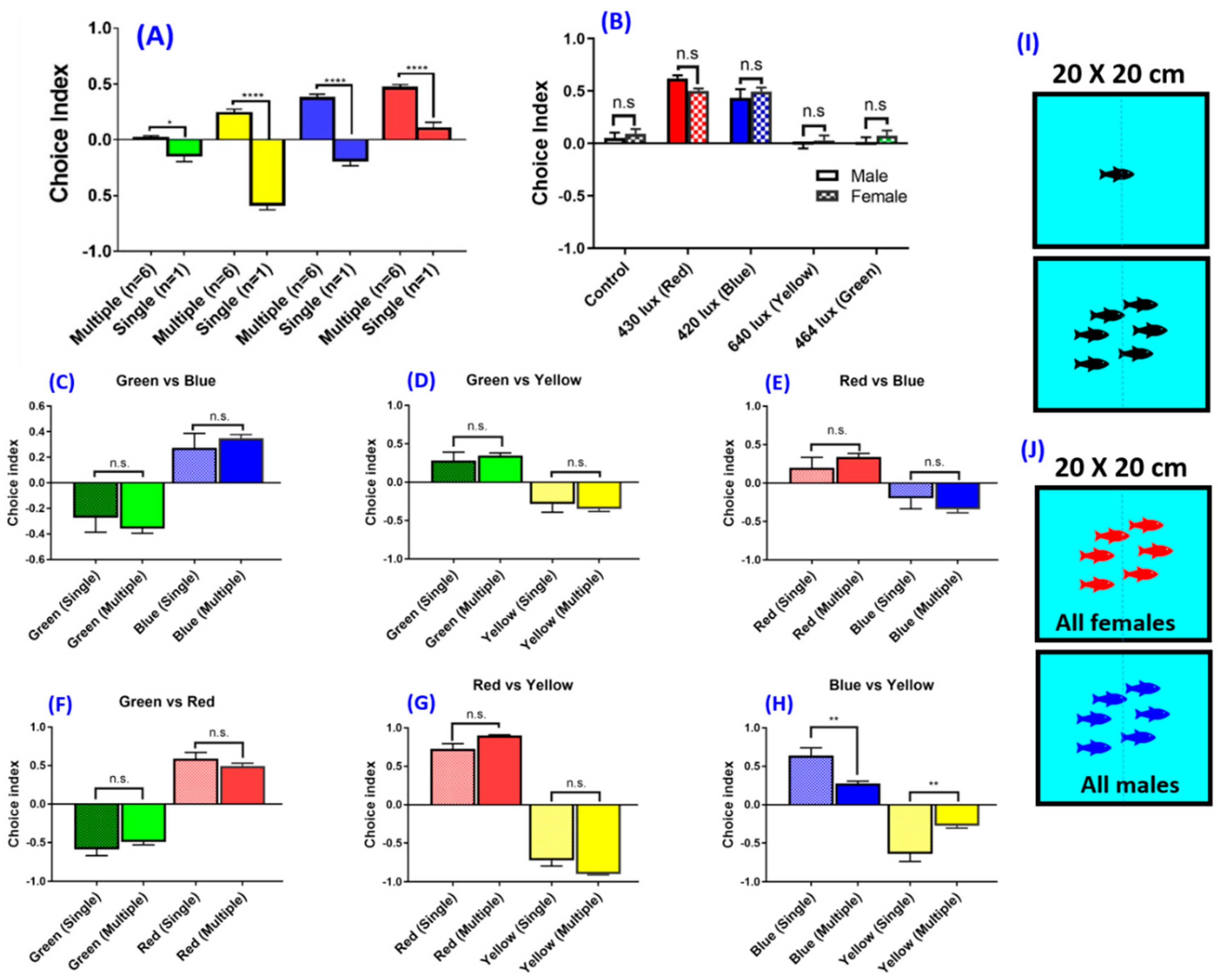
3.4. Experiment 3 and 4: Social Interaction and Gender on Zebrafish Color Preference
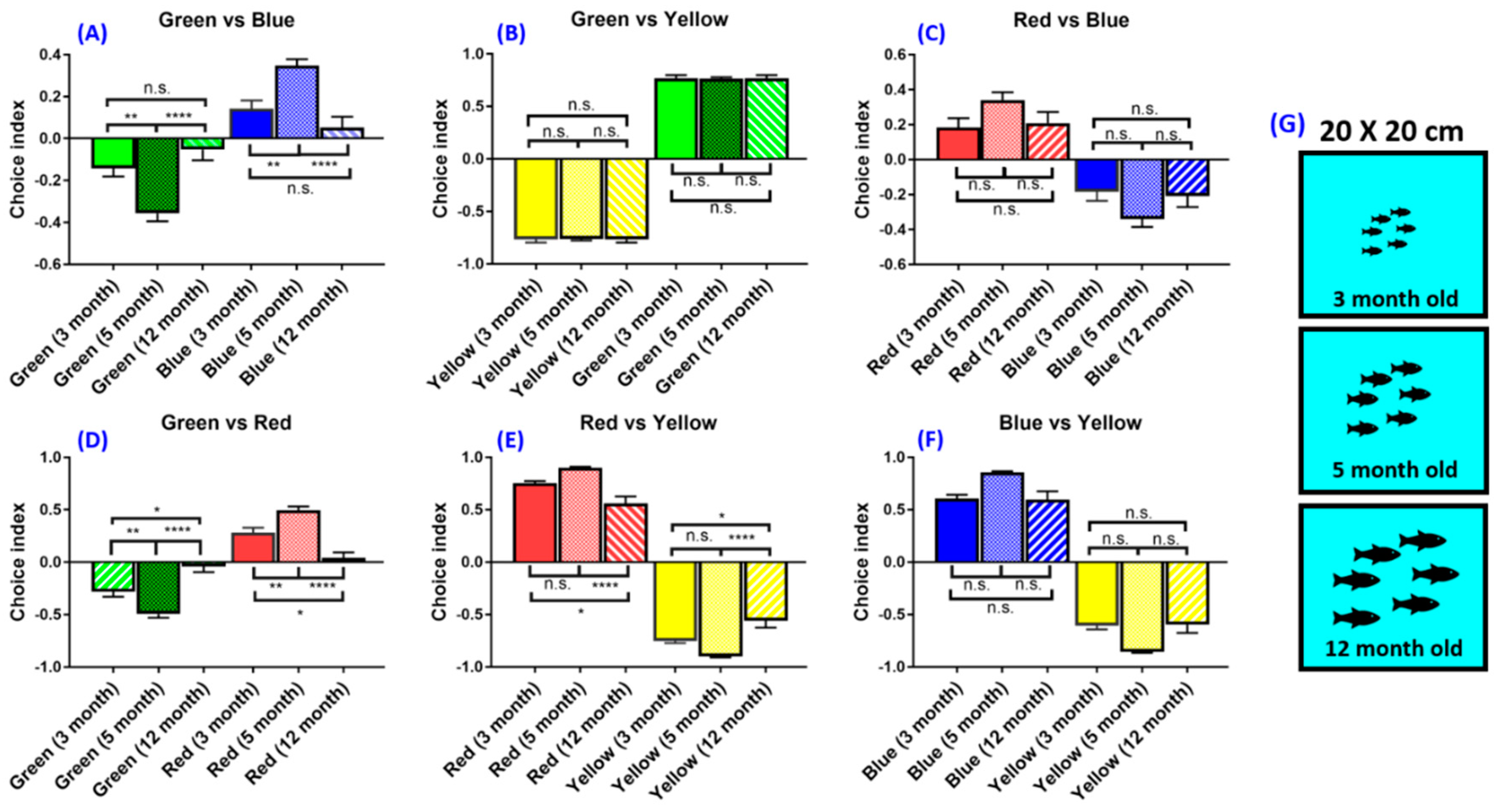
3.5. Experiment 5: Age Effect on Zebrafish Color Preferences
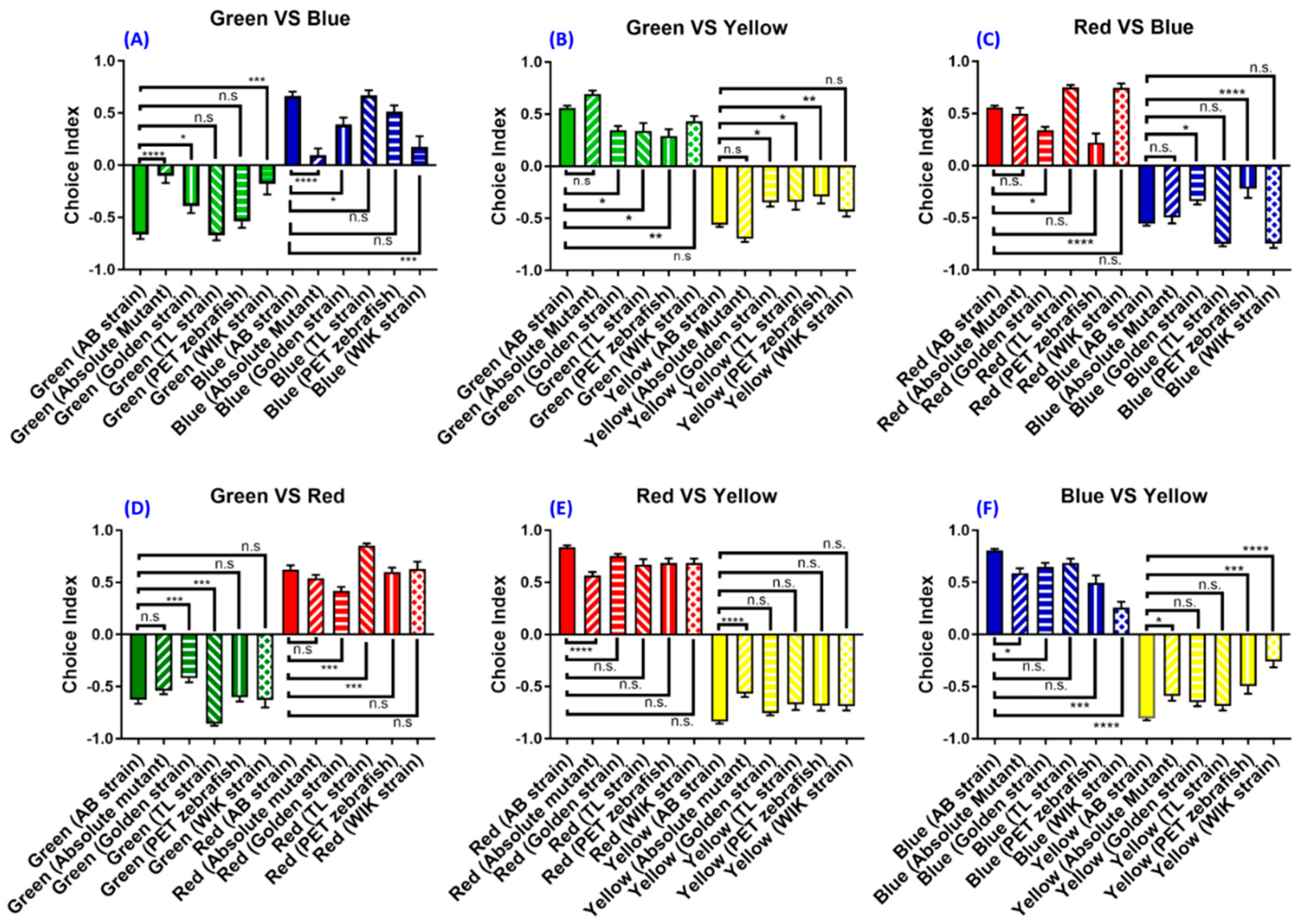
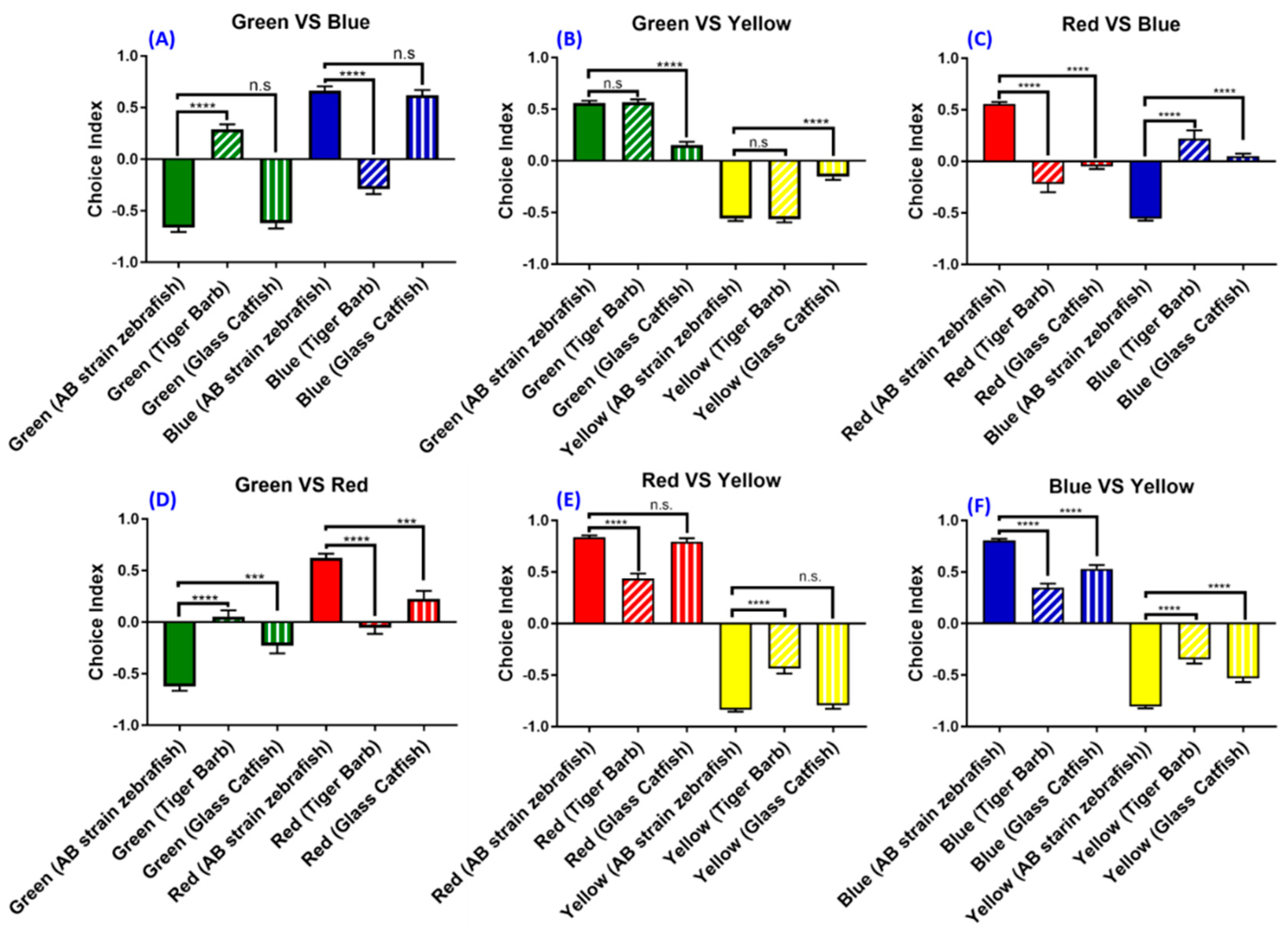
3.6. Experiment 6: The Strain- and Species-Specific Effect on Zebrafish Color Preference
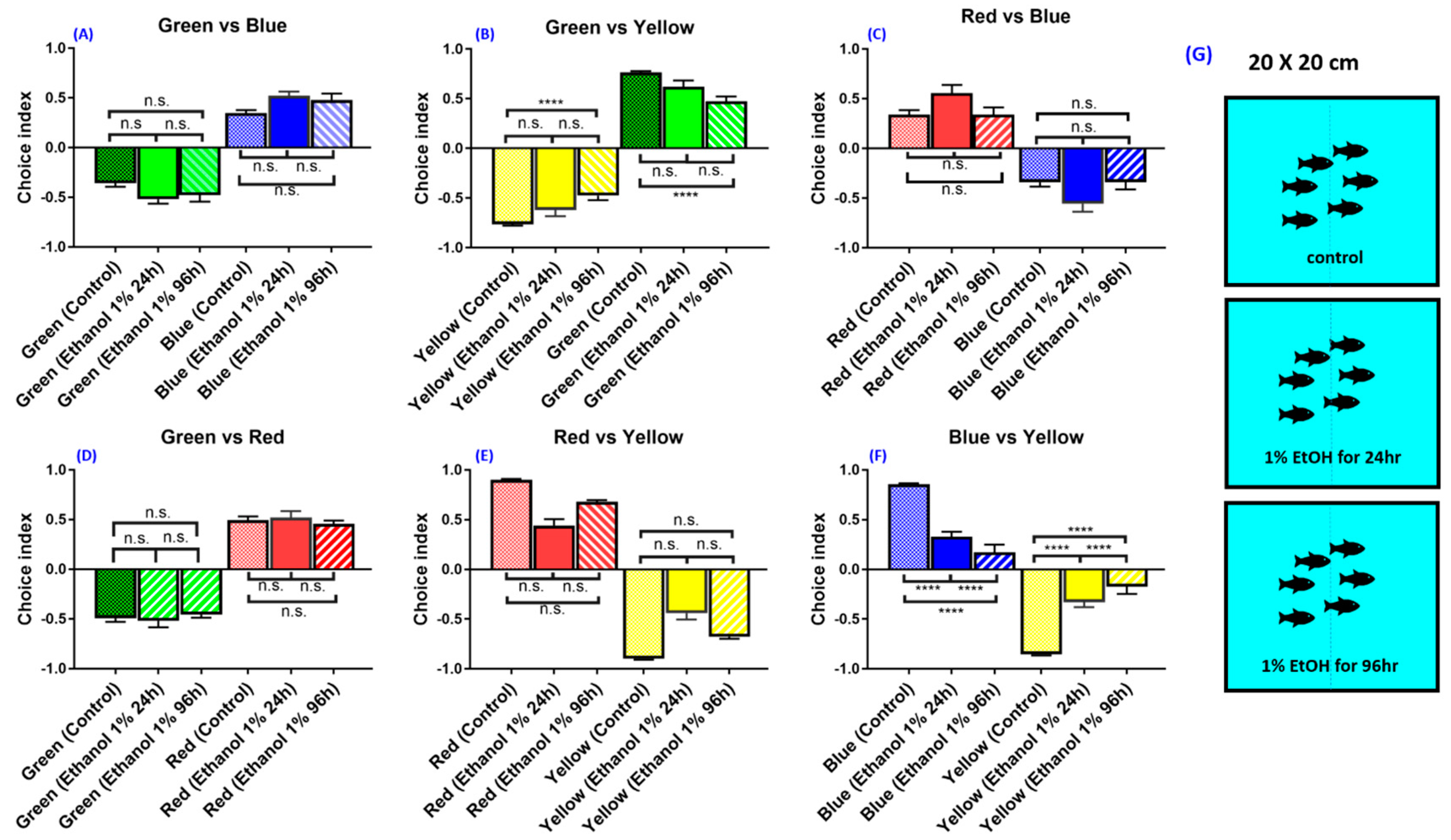
3.7. Experiment 7: Effect of Ethanol on Zebrafish Color Preference
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Color | Bottom Light Intensity (Lux) | Top Light Intensity (Lux) |
|---|---|---|
| Red | 440 | 430 |
| Yellow | 450 | 533 |
| Green | 430 | 497 |
| Blue | 410 | 424 |
| Color | Wavelength Spectrum (nm) and Maximum Wavelength | Reflectance (%) | Irradiance (uWatt/nm) |
|---|---|---|---|
| Red | 625–740 (625) | 97 | 0.000489 |
| Yellow | 565–590 (572) | 99 | 0.000587 |
| Green | 500–565 (541) | 100 | 0.000376 |
| Blue | 450–485 (456) | 98 | 0.000358 |
| White | 380–700 (561) | 100 | 0.000691 |
| Gray | 400–700 (564) | 96 | 0.000207 |
| Black | 500–700 (588) | 80 | 0.0000342 |
| Illumination Direction | Tank Type | Experimental Design | Fish Gender | Fish Strain | Color Preferences Ranking | References |
|---|---|---|---|---|---|---|
| Top | T-Maze with colored plastic sleeves | Single fish | N.D. | AB | Red = Green > Yellow = Blue = No color | [11] |
| Two-chambered PP (23 × 15 × 15 cm) | n = 12 | |||||
| Bottom (LED plate) | Transparent acrylic plastic sheeting with a thickness of 3 mm. The dimensions of the tank 230 × 150 × 150 mm | Single fish | N.D. | AB | Blue > White > Red | [13] |
| n = 18 | ||||||
| Top | T-Maze | Single Fish | Male and Female | AB | Blue > Purple > Green > Yellow = Orange > Blank | [3] |
| n = 7–10 | ||||||
| Bottom | T-maze setup | n = 12 | N.D. | AB | Blue > Red > Green > Yellow | [7] |
| (Noldus IT (Information Technology)) | ||||||
| Bottom | Cross maze | Larvae | N.D. | Wild-type purchased from local pet stores | Blue > Red > Green | [6] |
| n = 40 | ||||||
| Top | 20 l glass aquaria | Group of 10 fish | N.D. | Zebrafish obtained from a commercial supplier | Red > Green > >White > Blue | [2] |
| (Colored Light) | (40 × 25 × 30 cm) | |||||
| Bottom | 50 cm diameter transparent plastic tanks were divided into four laterals compartments of a similar size of 7 cm | n = 12 | N.D. | Zebrafish obtained from a local fish farm | Blue = Green > Red = Yellow | [1] |
| Top | Cross-maze with sidearm covered in different colors | Single fish | N.D. | AB | Blue > Red = Green > Yellow | [14] |
| n minimum = 8 |
References
- Oliveira, J.; Silveira, M.; Chacon, D.; Luchiari, A. The Zebrafish World of Colors and Shapes: Preference and Discrimination. Zebrafish 2015, 12, 166–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, R.; Smith, C. Innate and Learned Colour Preference in the Zebrafish, Danio rerio. Ethology 2008, 114, 582–588. [Google Scholar] [CrossRef]
- Bault, Z.A.; Peterson, S.M.; Freeman, J.L. Directional and color preference in adult zebrafish: Implications in behavioral and learning assays in neurotoxicology studies. J. Appl. Toxicol. JAT 2015, 35, 1502–1510. [Google Scholar] [CrossRef]
- Xu, X.; Weber, D.; Burge, R.; VanAmberg, K. Neurobehavioral impairments produced by developmental lead exposure persisted for generations in zebrafish (Danio rerio). Neurotoxicology 2016, 52, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.H.; Cho, H.J.; Han, E.; Hong, T.I.; Ariyasiri, K.; Choi, J.H.; Hwang, K.S.; Jeong, Y.M.; Yang, S.Y.; Yu, K.; et al. Zebrafish knockout of Down syndrome gene, DYRK1A, shows social impairments relevant to autism. Mol. Autism 2017, 8. [Google Scholar] [CrossRef]
- Park, J.S.; Ryu, J.H.; Choi, T.I.; Bae, Y.K.; Lee, S.; Kang, H.J.; Kim, C.H. Innate Color Preference of Zebrafish and Its Use in Behavioral Analyses. Mol. Cells 2016, 39, 750–755. [Google Scholar] [CrossRef] [Green Version]
- Peeters, B.W.M.M.; Moeskops, M.; Veenvliet, A.R.J. Color Preference in Danio rerio: Effects of Age and Anxiolytic Treatments. Zebrafish 2016, 13, 330–334. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Lee, Y.; Kim, D.; Jung, M.W.; Lee, C.-J. Scopolamine-induced learning impairment reversed by physostigmine in zebrafish. Neurosci. Res. 2010, 67, 156–161. [Google Scholar] [CrossRef]
- Yong-seok, C.; Chang-Joong, L.; Yeon-Hwa, K. MK-801-induced learning impairments reversed by physostigmine and nicotine in zebrafish. Anim. Cells Syst. 2011, 15, 115–121. [Google Scholar] [CrossRef]
- Cognato Gde, P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-Maze memory task in zebrafish (Danio rerio): The role of glutamatergic and cholinergic systems on the acquisition and consolidation periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef]
- Avdesh, A.; Martin-Iverson, M.T.; Mondal, A.; Chen, M.; Askraba, S.; Morgan, N.; Lardelli, M.; Groth, D.M.; Verdile, G.; Martins, R.N. Evaluation of color preference in zebrafish for learning and memory. J. Alzheimer’s Dis. JAD 2012, 28, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. Camb. Philos. Soc. 2008, 83, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Raghupathy, R.K.; Albalawi, A.; Zhao, Z.; Reilly, J.; Xiao, Q.; Shu, X. A colour preference technique to evaluate acrylamide-induced toxicity in zebrafish. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2017, 199, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Liu, X.; Sun, M.; Zhang, Q.; Li, T.; Li, X.; Xu, J.; Zhao, X.; Chen, D.; Feng, X. Reversal of reserpine-induced depression and cognitive disorder in zebrafish by sertraline and Traditional Chinese Medicine (TCM). Behav. Brain Funct. BBF 2018, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iovine, M.K.; Johnson, S.L. Genetic analysis of isometric growth control mechanisms in the zebrafish caudal Fin. Genetics 2000, 155, 1321–1329. [Google Scholar] [PubMed]
- Watanabe, M.; Iwashita, M.; Ishii, M.; Kurachi, Y.; Kawakami, A.; Kondo, S.; Okada, N. Spot pattern of leopard Danio is caused by mutation in the zebrafish connexin41. 8 gene. EMBO Rep. 2006, 7, 893–897. [Google Scholar] [CrossRef] [Green Version]
- Trainor, P. Neural Crest Cells: Evolution, Development and Disease; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Parichy, D.M.; Mellgren, E.M.; Rawls, J.F.; Lopes, S.S.; Kelsh, R.N.; Johnson, S.L. Mutational analysis of endothelin receptor b1 (rose) during neural crest and pigment pattern development in the zebrafish Danio rerio. Dev. Biol. 2000, 227, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Sumbre, G.; De Polavieja, G.G. The world according to zebrafish: How neural circuits generate behavior. Front. Neural Circuits 2014, 8, 91. [Google Scholar]
- Freeman, A.; Holland, R.; Hwang-Shum, J.-J.; Lains, D.; Matthews, J.; Murray, K.; Nasiadka, A.; Quinn, E.; Varga, Z.M.; Westerfield, M. 7 The Zebrafish International Resource Center. Biol. Resour. Model Org. Collect. Charact. Appl. 2019, 1, 113. [Google Scholar]
- Lamason, R.L.; Mohideen, M.-A.P.; Mest, J.R.; Wong, A.C.; Norton, H.L.; Aros, M.C.; Jurynec, M.J.; Mao, X.; Humphreville, V.R.; Humbert, J.E. SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 2005, 310, 1782–1786. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B.M.; Froehlich, J.M.; Galt, N.J.; Biga, P.R. Inbred strains of zebrafish exhibit variation in growth performance and myostatin expression following fasting. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; de Polavieja, G.G. idTracker: Tracking individuals in a group by automatic identification of unmarked animals. Nat. Methods 2014, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.Y.B.; Mathur, P.; Gould, G.G.; Guo, S. Identification of a brain center whose activity discriminates a choice behavior in zebrafish. Proc. Natl. Acad. Sci. USA 2011, 108, 2581–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Yao, Y.; Zhang, H.; Kawakami, K.; Du, J. Left habenula mediates light-preference behavior in zebrafish via an asymmetrical visual pathway. Neuron 2017, 93, 914–928.e914. [Google Scholar] [CrossRef] [Green Version]
- E Engeszer, R.; Alberici Da Barbiano, L.; Ryan, M.; M Parichy, D. Timing and Plasticity of Shoaling Behavior in the Zebrafish, Danio rerio. Anim. Behav. 2007, 74, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Norton, W.; Bally-Cuif, L. Adult zebrafish as a model organism for behavioural genetics. BMC Neurosci. 2010, 11, 90. [Google Scholar] [CrossRef] [Green Version]
- Okiyama, D.C.; Soares, I.D.; Cuevas, M.S.; Crevelin, E.J.; Moraes, L.A.; Melo, M.P.; Oliveira, A.L.; Rodrigues, C.E. Pressurized liquid extraction of flavanols and alkaloids from cocoa bean shell using ethanol as solvent. Food Res. Int. 2018, 114, 20–29. [Google Scholar] [CrossRef]
- Baümler, E.R.; Carrín, M.E.; Carelli, A.A. Extraction of sunflower oil using ethanol as solvent. J. Food Eng. 2016, 178, 190–197. [Google Scholar] [CrossRef]
- Santamaria, R.; Reyes-Duarte, M.; Barzana, E.; Fernando, D.; Gama, F.; Mota, M.; Lopez-Munguia, A. Selective enzyme-mediated extraction of capsaicinoids and carotenoids from chili guajillo puya (Capsicum annuum L.) using ethanol as solvent. J. Agric. Food Chem. 2000, 48, 3063–3067. [Google Scholar] [CrossRef] [Green Version]
- Barnes, G. The effects of ethyl alcohol on visual pursuit and suppression of the vestibulo-ocular reflex. Acta Oto-Laryngol. 1983, 96, 161–166. [Google Scholar] [CrossRef]
- Barnes, G.; Crombie, J.; Edge, A. The effects of ethanol on visual–vestibular interaction during active and passive head movements. Aviat. Space Environ. Med. 1985, 56, 695–701. [Google Scholar] [PubMed]
- Umeda, Y.; Sakata, E. Alcohol and the oculomotor system. Ann. Otol. Rhinol. Laryngol. 1978, 87, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, I. The influence of drugs and alcohol upon human eye movement. Proc. R. Soc. Med. 1976, 69, 479. [Google Scholar] [PubMed]
- Aschan, G. Different types of alcohol nystagmus. Acta Oto-Laryngol. 1958, 49, 69–78. [Google Scholar] [CrossRef]
- Baloh, R.; Sharma, S.; Moskowitz, H.; Griffith, R. Effect of alcohol and marihuana on eye movements. Aviat. Space Environ. Med. 1979, 50, 18–23. [Google Scholar]
- Harder, T.; Reker, U. Influence of low dose alcohol on fixation suppression. Acta Oto-Laryngol. 1995, 115, 33–36. [Google Scholar] [CrossRef]
- Takahashi, M.; Akiyama, I.; Tsujita, N.; Yoshida, A. The effect of alcohol on the vestibulo-ocular reflex and gaze regulation. Arch. Oto-Rhino Laryngol. 1989, 246, 195–199. [Google Scholar] [CrossRef]
- Matsui, J.I.; Egana, A.L.; Sponholtz, T.R.; Adolph, A.R.; Dowling, J.E. Effects of ethanol on photoreceptors and visual function in developing zebrafish. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4589–4597. [Google Scholar] [CrossRef] [Green Version]
- Miller, R. The effect of ingested alcohol on fusion latency at various viewing distances. Percept. Psychophys. 1991, 50, 575–583. [Google Scholar] [CrossRef]
- Sherwin, C.M.; Glen, E.F. Cage colour preferences and effects of home cage colour on anxiety in laboratory mice. Anim. Behav. 2003, 66, 1085–1092. [Google Scholar] [CrossRef] [Green Version]
- Lunau, K.; Papiorek, S.; Eltz, T.; Sazima, M. Avoidance of achromatic colours by bees provides a private niche for hummingbirds. J. Exp. Biol. 2011, 214, 1607–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endler, J.A.; Westcott, D.A.; Madden, J.R.; Robson, T. Animal visual systems and the evolution of color patterns: Sensory processing illuminates signal evolution. Evolution 2005, 59, 1795–1818. [Google Scholar] [CrossRef] [PubMed]
- Dangles, O.; Irschick, D.; Chittka, L.; Casas, J. Variability in sensory ecology: Expanding the bridge between physiology and evolutionary biology. Q. Rev. Biol. 2009, 84, 51–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cachat, J.; Stewart, A.; Utterback, E.; Hart, P.; Gaikwad, S.; Wong, K.; Kyzar, E.; Wu, N.; Kalueff, A.V. Three-dimensional neurophenotyping of adult zebrafish behavior. PLoS ONE 2011, 6, e17597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faustino, A.I.; Tacão-Monteiro, A.; Oliveira, R.F. Mechanisms of social buffering of fear in zebrafish. Sci. Rep. 2017, 7, 44329. [Google Scholar] [CrossRef] [Green Version]
- Agrillo, C.; Piffer, L.; Bisazza, A. Number versus continuous quantity in numerosity judgments by fish. Cognition 2011, 119, 281–287. [Google Scholar] [CrossRef]
- Morris, A.; Fadool, J. Studying rod photoreceptor development in zebrafish. Physiol. Behav. 2005, 86, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Liu, H.; Huang, B.; Wagle, M.; Guo, S. Identification of environmental stressors and validation of light preference as a measure of anxiety in larval zebrafish. BMC Neurosci. 2016, 17, 63. [Google Scholar] [CrossRef] [Green Version]
- Maximino, C.; de Brito, T.M.; de Mattos Dias, C.A.G.; Gouveia, A.J.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209. [Google Scholar] [CrossRef]
- Bilotta, J. Effects of abnormal lighting on the development of zebrafish visual behavior. Behav. Brain Res. 2000, 116, 81–87. [Google Scholar] [CrossRef]
- Anchelin, M.; Alcaraz-Pérez, F.; Martínez, C.M.; Bernabé-García, M.; Mulero, V.; Cayuela, M.L. Premature aging in telomerase-deficient zebrafish. Dis. Models Mech. 2013, 6, 1101–1112. [Google Scholar] [CrossRef] [Green Version]
- Chinen, A.; Hamaoka, T.; Yamada, Y.; Kawamura, S. Gene duplication and spectral diversification of cone visual pigments of zebrafish. Genetics 2003, 163, 663–675. [Google Scholar] [PubMed]
- Nawrocki, L.; Bremiller, R.; Streisinger, G.; Kaplan, M. Larval and adult visual pigments of the zebrafish, Brachydanio rerio. Vis. Res. 1985, 25, 1569–1576. [Google Scholar] [CrossRef]
- Branchek, T. The development of photoreceptors in the zebrafish, brachyDanio rerio. II. Function. J. Comp. Neurol. 1984, 224, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Schmitt, E.A.; Dowling, J.E. Temporal and spatial patterns of opsin gene expression in zebrafish (Danio rerio). Vis. Neurosci. 1995, 12, 895–906. [Google Scholar] [CrossRef]
- Saszik, S.; Bilotta, J.; Givin, C.M. ERG assessment of zebrafish retinal development. Vis. Neurosci. 1999, 16, 881–888. [Google Scholar] [CrossRef]
- Korenbrot, J.I.; Mehta, M.; Tserentsoodol, N.; Postlethwait, J.H.; Rebrik, T.I. EML1 (CNG-modulin) controls light sensitivity in darkness and under continuous illumination in zebrafish retinal cone photoreceptors. J. Neurosci. 2013, 33, 17763–17776. [Google Scholar] [CrossRef] [Green Version]
- Blaser, R.E.; Rosemberg, D.B. Measures of anxiety in zebrafish (Danio rerio): Dissociation of black/white preference and novel tank test. PLoS ONE 2012, 7, e36931. [Google Scholar] [CrossRef] [Green Version]
- Pieróg, M.; Guz, L.; Doboszewska, U.; Poleszak, E.; Wlaź, P. Effects of alprazolam treatment on anxiety-like behavior induced by color stimulation in adult zebrafish. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 82, 297–306. [Google Scholar] [CrossRef]
- Roy, T.; Suriyampola, P.S.; Flores, J.; López, M.; Hickey, C.; Bhat, A.; Martins, E.P. Color preferences affect learning in zebrafish, Danio rerio. Sci. Rep. 2019, 9, 14531. [Google Scholar] [CrossRef] [Green Version]
- Fuller, R.C.; Noa, L.A.; Strellner, R.S. Teasing apart the many effects of lighting environment on opsin expression and foraging preference in bluefin killifish. Am. Nat. 2010, 176, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, S. Cryptic and conspicuous coloration in the pelagic environment. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Protasov, V.R. Vision and Near Orientation of Fish; Israel Program for Scientific Translations: Jerusalem, Israel, 1970; Volume 5738. [Google Scholar]
- Ullmann, J.F.; Gallagher, T.; Hart, N.S.; Barnes, A.C.; Smullen, R.P.; Collin, S.P.; Temple, S.E. Tank color increases growth, and alters color preference and spectral sensitivity, in barramundi (Lates calcarifer). Aquaculture 2011, 322, 235–240. [Google Scholar] [CrossRef]
- Carruthers, H.R.; Morris, J.; Tarrier, N.; Whorwell, P.J. The Manchester Color Wheel: Development of a novel way of identifying color choice and its validation in healthy, anxious and depressed individuals. BMC Med. Res. Methodol. 2010, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Langen, B.; Dietze, S.; Fink, H. Acute effect of ethanol on anxiety and 5-HT in the prefrontal cortex of rats. Alcohol 2002, 27, 135–141. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siregar, P.; Juniardi, S.; Audira, G.; Lai, Y.-H.; Huang, J.-C.; Chen, K.H.-C.; Chen, J.-R.; Hsiao, C.-D. Method Standardization for Conducting Innate Color Preference Studies in Different Zebrafish Strains. Biomedicines 2020, 8, 271. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080271
Siregar P, Juniardi S, Audira G, Lai Y-H, Huang J-C, Chen KH-C, Chen J-R, Hsiao C-D. Method Standardization for Conducting Innate Color Preference Studies in Different Zebrafish Strains. Biomedicines. 2020; 8(8):271. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080271
Chicago/Turabian StyleSiregar, Petrus, Stevhen Juniardi, Gilbert Audira, Yu-Heng Lai, Jong-Chin Huang, Kelvin H.-C. Chen, Jung-Ren Chen, and Chung-Der Hsiao. 2020. "Method Standardization for Conducting Innate Color Preference Studies in Different Zebrafish Strains" Biomedicines 8, no. 8: 271. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080271