Use of Oral Anticoagulation and Diabetes Do Not Inhibit the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes


 and
and
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Collective
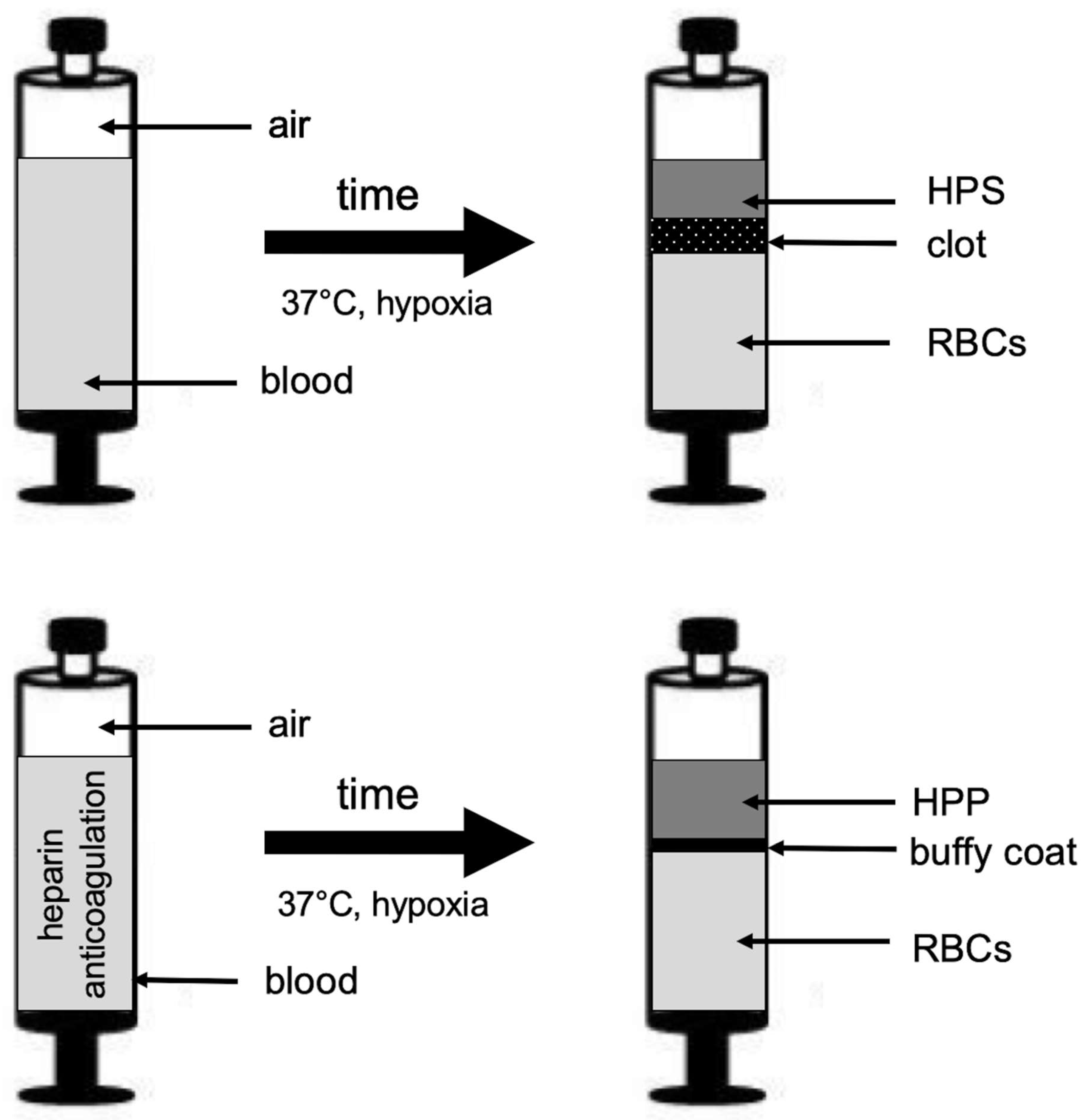
2.2. Preparation of Blood Plasma/Serum and Hypoxia Preconditioned Plasma (HPP)/Serum (HPS) Samples
2.3. Quantitative Analysis of VEGF, TPS-1, and PF-4 Concentrations in Blood-Derived Secretomes
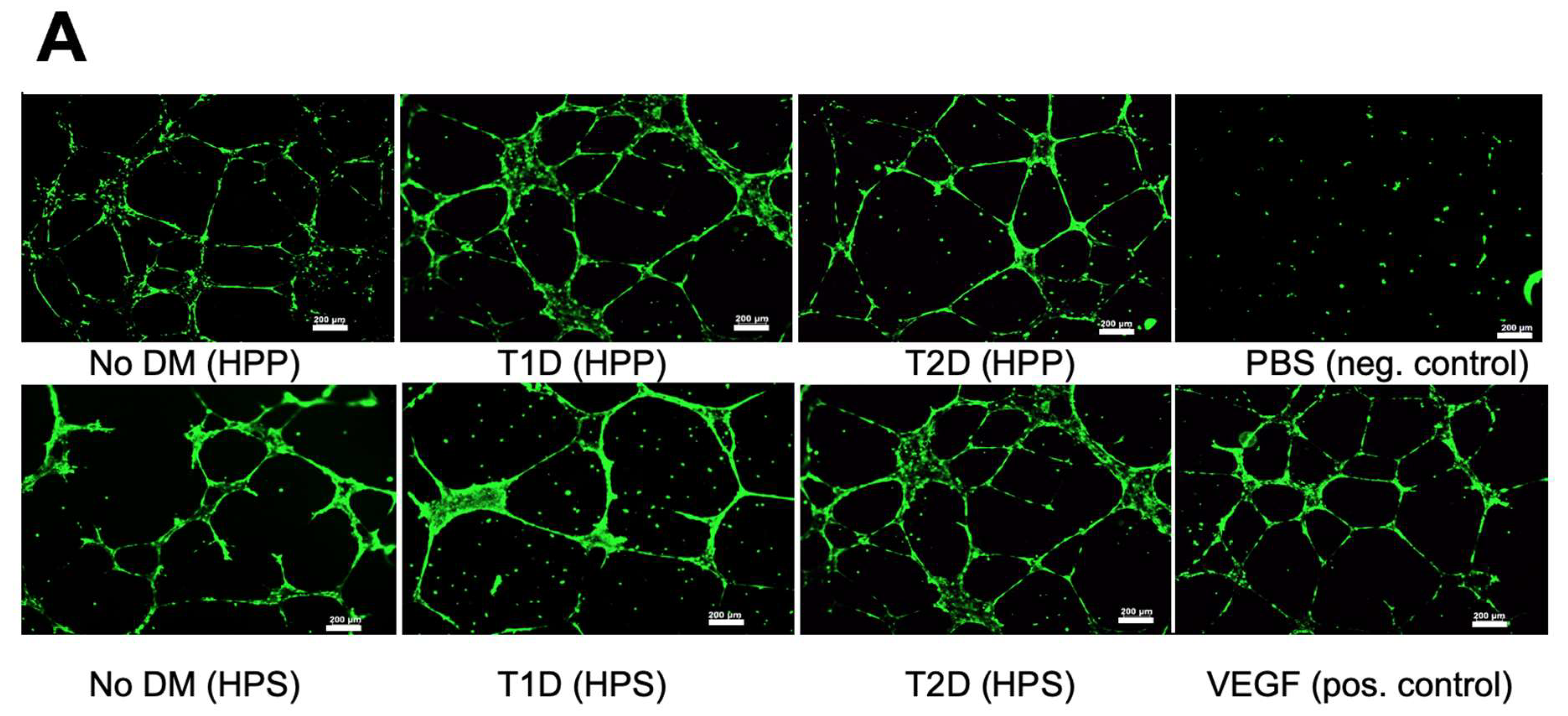
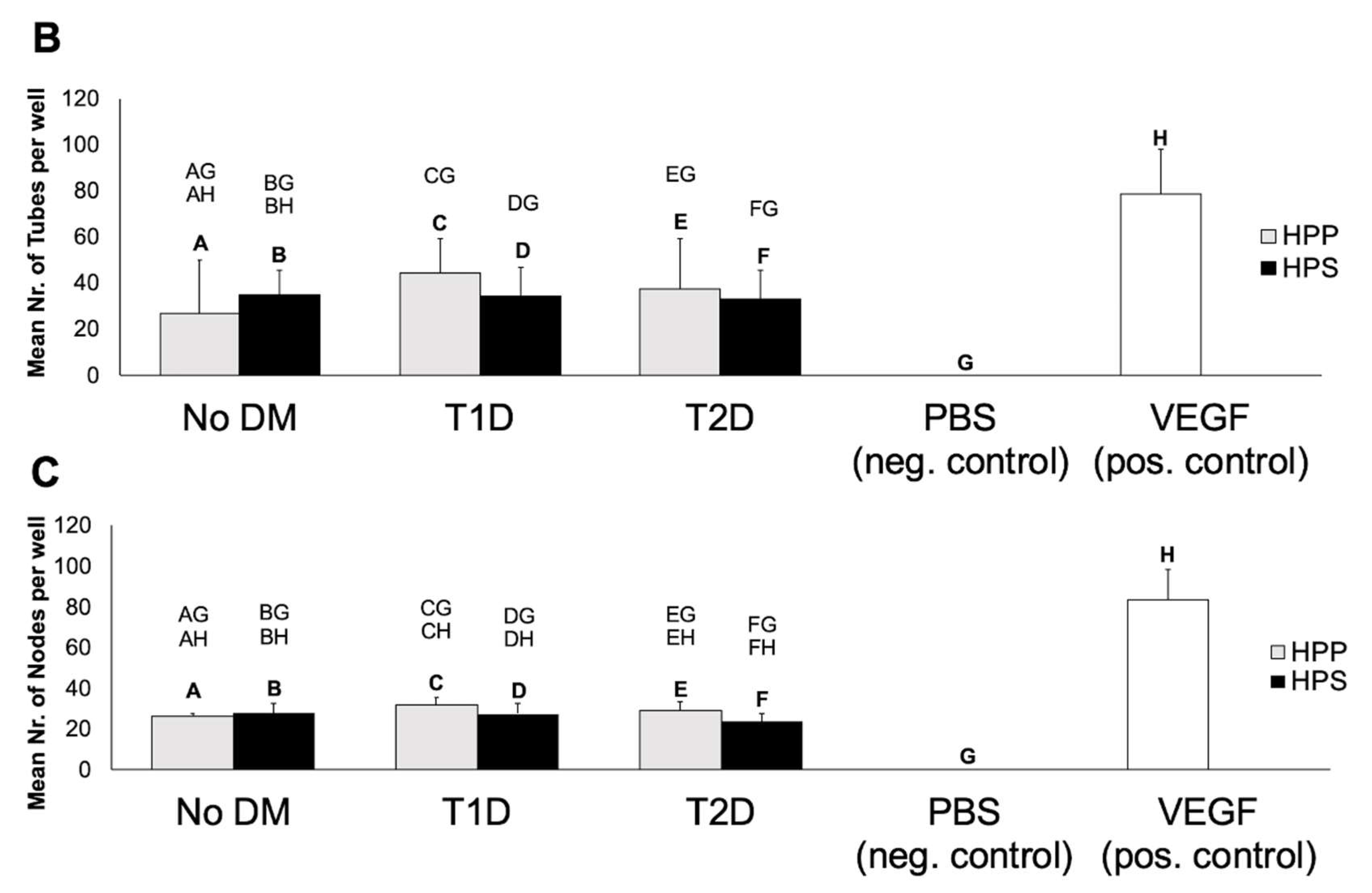
2.4. Analysis of the Effect of Blood-Derived Secretomes on Microvessel Formation In Vitro
2.5. Statistical Analysis
3. Results
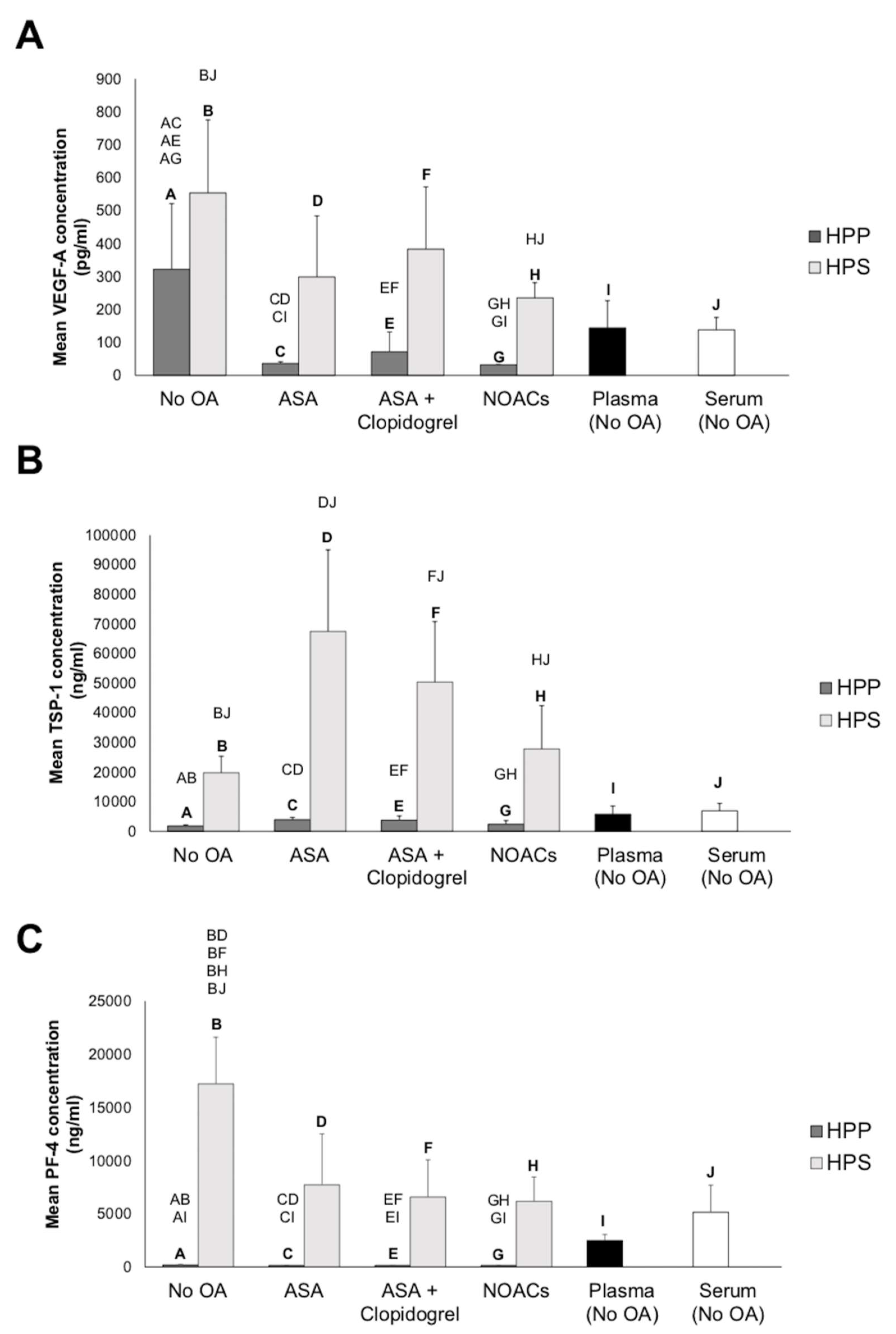
3.1. Effect of Oral Anticoagulation on Pro- (VEGF) and Anti-(TSP-1 and PF-4)Angiogenic Growth Factor Concentrations in Hypoxia Preconditioned Blood-Derived Secretomes
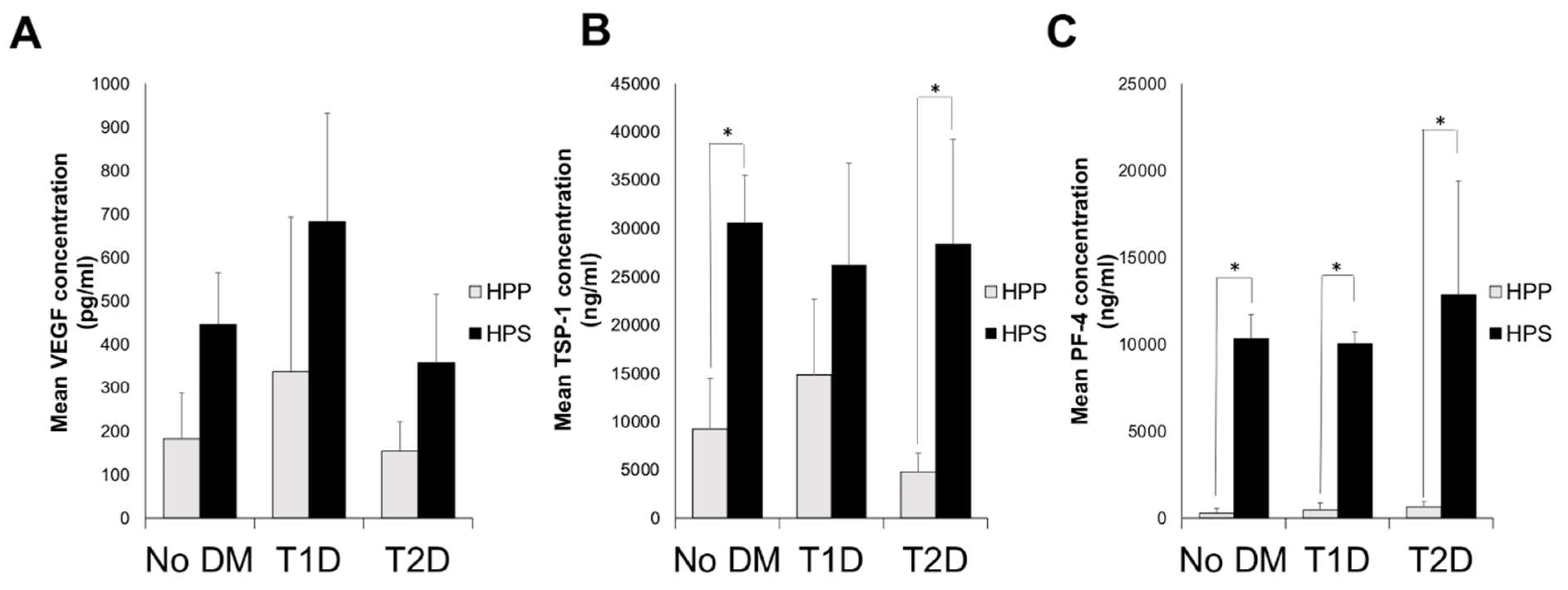
3.2. Quantitative Analysis of Blood-Derived Secretome Pro- (VEGF) and Anti-(TSP-1 and PF-4)Angiogenic Growth Factor Concentrations in Diabetic Subjects
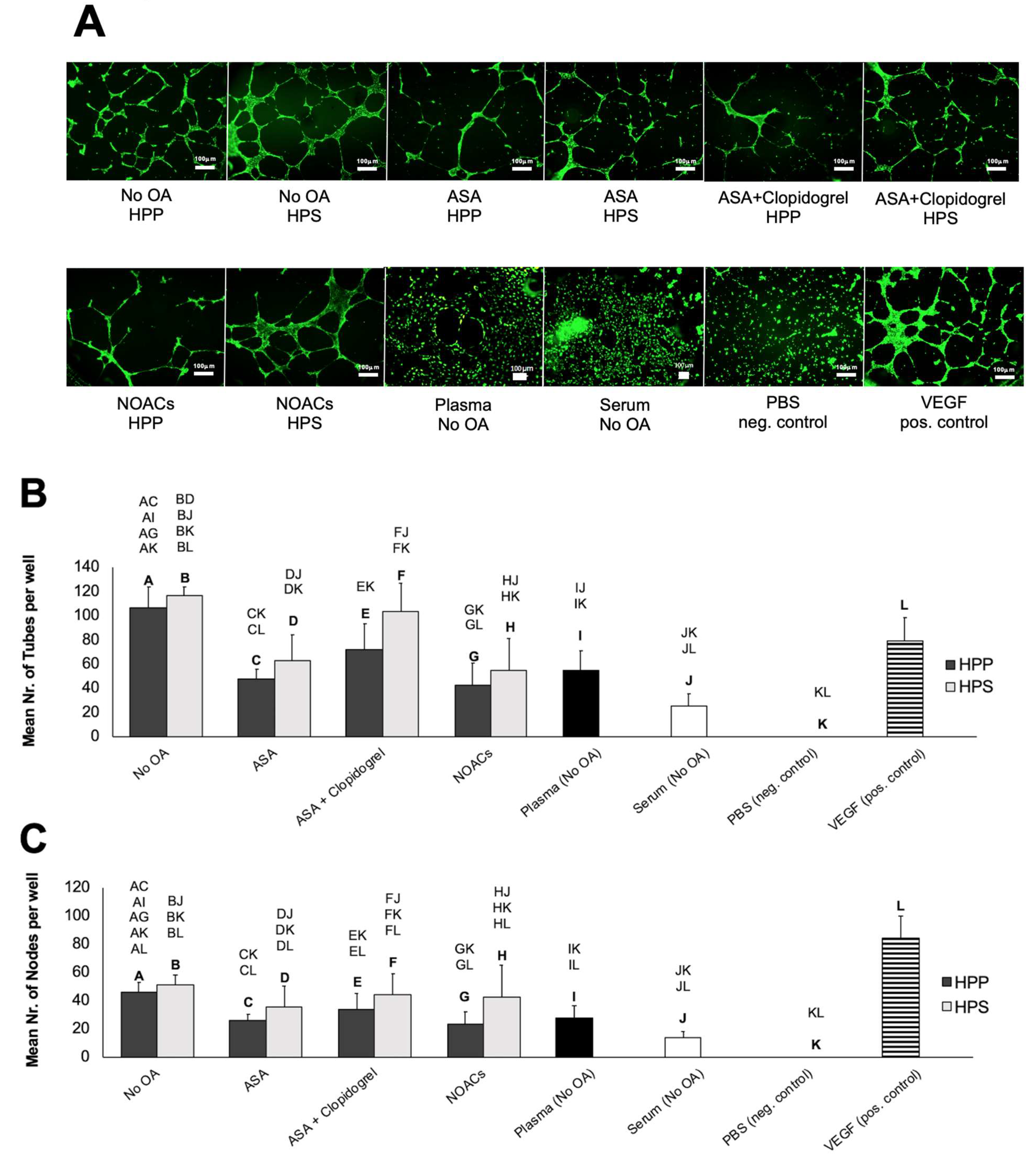
3.3. Effects of Oral Anticoagulation on the Ability of Hypoxia Preconditioned Blood-Derived Secretomes to Induce Angiogenesis In Vitro
3.4. Influence of Diabetes Mellitus on the Ability of Hypoxia Preconditioned Blood-Derived Secretomes to Induce Angiogenesis In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASA | Acetylsalicylic acid |
| bFGF | basic fibroblast growth factor |
| COX1 | Cyclooxygenase-1 |
| EGF | epidermal growth factor |
| EWS | Extracorporeal Wound Simulation |
| HPP | hypoxia preconditioned plasma |
| HPS | hypoxia preconditioned serum |
| HUVECs | human umbilical vein endothelial cells |
| IGF-1 | insulin-like growth factor-1 |
| NOACs | Nonvitamin K antagonist oral anticoagulants |
| OA | oral anticoagulation |
| PAR1 | Protease-activated receptor-1 |
| PBC | peripheral blood cells |
| PBS | phosphate buffered saline |
| PDGF | platelet-derived growth factor |
| PF-4 | platelet factor-4 |
| TGF-beta1 | transforming growth factor beta-1 |
| TSP-1 | thrombospondin-1 |
| TXA2 | thromboxan-A2 |
| VEGF | vascular endothelial growth factor |
References
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Stadelmann, W.K.; Digenis, A.G.; Tobin, G.R. Physiology and healing dynamics of chronic cutaneous wounds. Am. J. Surg. 1998, 176, 26–38. [Google Scholar] [CrossRef]
- Avishai, E.; Yeghiazaryan, K.; Golubnitschaja, O. Impaired wound healing: Facts and hypotheses for multi-professional considerations in predictive, preventive and personalised medicine. EPMA J. 2017, 8, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarus, G.S.; Cooper, D.M.; Knighton, D.R.; Margolis, D.J.; Pecoraro, R.E.; Rodeheaver, G.; Robson, M.C. Definitions and quidlines for assessment of wounds and evaluation of healing. Arch. Dermatol. 1994, 130, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Woo, K.; Brandys, T.; Marin, J. Assessing chronic wound perfusion in the lower extremity: Current and emerging approaches. Chronic Wound Care Manag. Res. 2015, 2, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Thiruvoipati, T.; Kielhorn, C.E.; Armstrong, E.J. Peripheral artery disease in patients with diabetes: Epidemiology, mechanisms, and outcomes. World J. Diabetes 2015, 6, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Tuttolomondo, A.; Maida, C.; Pinto, A. Diabetic foot syndrome: Immune-inflammatory features as possible cardiovascular markers in diabetes. World J. Orthop. 2015, 6, 62–76. [Google Scholar] [CrossRef] [Green Version]
- Skelly, C.L.; Cifu, A.S. Screening, Evaluation, and Treatment of Peripheral Arterial Disease. JAMA 2016, 316, 1486–1487. [Google Scholar] [CrossRef] [PubMed]
- Emini Veseli, B.; Perotta, P.; De Meyer, G.R.A.; Roth, L.; Van der Donckt, C.; Martinet, W.; De Meyer, G.R.Y. Animal models of atherosclerosis. Eur. J. Pharmacol. 2017, 5, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Luisis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Rafieian-Kopaei, M.; Setorki, M.; Doudi, M.; Baradaran, A.; Nasri, H. Atherosclerosis: Process, Indicators, Risk Factors and New Hopes. Int. J. Prev. Med. 2014, 5, 927–946. [Google Scholar] [PubMed]
- Bentzon, J.F.; Otusua, F.; Virmani, R.; Falk, E. Mechanisms of Plaque Formation and Rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef] [PubMed]
- Aronson, D.; Edelmann, E.R. Revascularization for coronary artery disease in diabetes mellitus: Angioplasty, stents and coronary artery bypass grafting. Rev. Endocr. Metab. Disord. 2010, 11, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Bolton, L. Peripheral arterial disease: Scoping review of patient-centred outcomes. Int. Wound J. 2019, 16, 1521–1532. [Google Scholar] [CrossRef]
- Abu Dabrh, A.M.; Steffen, M.W.; Undavalli, C.; Asi, N.; Wang, Z.; Elamin, M.B.; Conte, M.S.; Murad, M.H. The natural history of untreated severe or critical limb ischemia. J. Vasc. Surg. 2015, 62, 1642–1651. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.Z.; Ng, N.S.; Thomas, C. Prevention and treatment of diabetic foot ulcers. J. R. Soc. Med. 2017, 110, 104–109. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, D.F.; Caruso, G.J.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The Treatment of Impaired Wound Healing in Diabetes: Looking among Old Drugs. Pharmaceuticals 2020, 3, 60. [Google Scholar] [CrossRef] [Green Version]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Davis, F.M.; Kimball, A.; Boniakowski, A.; Gallagher, K. Dysfunctional Wound Healing in Diabetic Foot Ulcers: New Crossroads. Curr. Diabetes Rep. 2018, 18, 2. [Google Scholar] [CrossRef]
- Dinh, T.; Elder, S.; Veves, A. Delayed wound healing in diabetes: Considering future treatments. Diabetes Manag. 2011, 1, 509–519. [Google Scholar] [CrossRef]
- Greenhalgh, D.G. Wound healing and diabetes mellitus. Clin. Plast. Surg. 2003, 30, 37–45. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; DiPietro, L. Diabetes and Wound Angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinh, T.; Veves, A. Microcirculation of the Diabetic Foot. Curr. Pharm. Des. 2005, 11, 2301–2309. [Google Scholar] [CrossRef]
- Honnegowda, T.M.; Kumar, P.; Udupa, E.G.; Kumar, S.; Kumar, U.; Rao, P. Role of angiogenesis and angiogenic factors in acute and chronic wound healing. Plast. Aesthet. Res. 2015, 2, 243–249. [Google Scholar]
- Mustoe, T.A.; Tae, A.S.; Tarpley, J.E.; Pierce, G.F. Role of hypoxia in growth factor responses: Differential effects of basic fibroblast growth factor and platelet-derived growth factor in an ischemic wound model. Wound Repair Regen. 1994, 2, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Al Said, S.; Bode, C.; Duerschmied, D. Anticoagulation in Atherosclerotic Disease. Hamostaseologie 2018, 38, 240–246. [Google Scholar]
- Krankenberg, K. Recommended interventions for the treatment of peripheral artery disease: Keep the patients moving. Internist 2019, 60, 1235–1239. [Google Scholar] [CrossRef]
- Olinic, D.M.; Tataru, D.; Homorodean, C.; Spinu, M.; Olinic, M. Antithrombotic treatment in peripheral artery disease. VASA 2018, 47, 99–108. [Google Scholar] [CrossRef]
- Phelps, E.A.; Garcia, A.J. Update on therapeutic vascularization strategies. Regen Med. 2009, 4, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Kosaric, N.; Kiwanuka, H.; Gurtner, G.C. Stem cell therapies for wound healing. Expert Opin. Biol. Ther. 2019, 19, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.M.; Bauer, R.J.; Velazquez, O.C. Angiogenesis, vasculogenesis, and induction of healing in chronic wounds. Vasc. Endovasc. Surg. 2005, 39, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjipanayi, E.; Schilling, A.F. Regeneration through autologous hypoxia preconditioned plasma. Organogenesis 2014, 10, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Hadjipanayi, E.; Schilling, A.F. Hypoxia- based strategies for angiogenic induction. Organog. Landes Biosci. 2013, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hadjipanayi, E.; Bauer, A.T.; Moog, P.; Salgin, B.; Kükrek, H.; Fersch, B.; Hopfner, U.; Meissner, T.; Schlüter, A.; Ninkovic, M.; et al. Cell-free Carrier System for Localised Delivery of Peripheral Blood Cell-Derived Engineered Factor Signaling: Towards Development of a One-Step Device for Autologous Angiogenic Therapy. J. Control. Release 2013, 169, 91–102. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Moog, P.; Bekeran, S.; Kirchhoff, K.; Berezhnoi, A.; Aguirre, J.; Bauer, A.T.; Kükrek, H.; Schmauss, D.; Hopfner, U.; et al. In Vitro Characterization of Hypoxia Preconditioned Serum (HPS)-Fibrin Hydrogels: Basis for an Injectable Biomimetic Tissue Regeneration Therapy. J. Funct. Biomater. 2019, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Hadjipanayi, E.; Bekeran, S.; Moog, P. Extracorporeal Wound Simulation as a Foundation for Tissue Repair und Regeneration Therapies. Int. J. Transpl. Plast. Surg. 2018, 2, 1–10. [Google Scholar]
- Moog, P.; Kirchhoff, K.; Bekeran, S.; Bauer, A.T.; Isenburg, S.; Dornseifer, U.; Machens, H.G.; Schilling, A.F.; Hadjipanayi, E. Comparative Evaluation of the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes and Platelet-Rich Plasma: An In Vitro Analysis. Biomedicines 2020, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Koutsoumpelis, A.; Argyriou, C.; Tasopoulou, K.M.; Georgakarakos, E.I.; Georgiadis, G.S. Novel Oral Anticoagulants in Peripheral Artery Disease: Current Evidence. Curr. Pharm. Des. 2018, 24, 4511–4515. [Google Scholar] [CrossRef]
- Gupta, A.; Lee, M.S.; Gupta, K.; Kumar, V.; Reddy, S. A Review of Antithrombotic Treatment in Critical Limb Ischemia After Endovascular Intervention. Cardiol. Ther. 2019, 8, 193–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjipanayi, E.; Kuhn, P.H.; Moog, P.; Bauer, A.T.; Kuekrek, H.; Mirzoyan, L.; Hummel, A.; Kirchhoff, K.; Salgin, B.; Isenburg, S.; et al. The Fibrin Matrix regulates Angiogenic Responses within the Hemostatic Microenvironment through Biochemical Control. PLoS ONE 2015, 10, e0135618. [Google Scholar] [CrossRef] [PubMed]
- Simon, E.M.; Streitz, M.J.; Sessions, D.J.; Kaide, C.G. Anticoagulation Reversal. Emerg. Med. Clin. N. Am. 2018, 36, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Tickner, A.; Klinghard, C.; Arnold, J.F.; Marmolejo, V. Total Contact Cast Use in Patients With Peripheral Arterial Disease: A Case Series and Systematic Review. Wounds 2018, 30, 49–56. [Google Scholar]
- Sampson, S.; Gerhardt, M.; Mandelbaum, B. Platelet rich plasma injection grafts for musculoskeletal injuries: A review. Curr. Rev. Musculoskelet. Med. 2008, 1, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.J.; Fylling, C.P.; Parnell, L.K. Use of platelet rich plasma gel on wound healing: A systematic review and meta-analysis. Eplasty 2011, 11, e38. [Google Scholar]
- Thun, M.J.; Henley, S.J.; Patrono, C. Nonsteroidal anti-inflammatory drugs as anticancer agents: Mechanistic, pharmacologic, and clinical issues. J. Natl. Cancer Inst. 2002, 94, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, E.J.; Newton, C.; Stevens, V.L.; Campbell, P.T.; Freedland, S.J.; Gapstur, S.M. Daily aspirin use and prostate cancer-specific mortality in a large cohort of men with nonmetastatic prostate cancer. J. Clin. Oncol. 2014, 32, 3716–3722. [Google Scholar] [CrossRef]
- Li, H.; Liu, K.; Boardman, L.A.; Zhao, Y.; Wang, L.; Sheng, Y.; Oi, N.; Limbgurg, P.J.; Bode, A.M.; Dong, Z. Circulating prostaglandin biosynthesis in colorectal cancer and potential clinical significance. Ebiomedicine 2015, 2, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Yan, J.; Fu, X.; Pan, Q.; Sun, D.; Xu, Y.; Wang, J.; Nie, L.; Tong, L.; Shen, A.; et al. Aspirin Inhibits Cancer Metastasis and Angiogenesis via Targeting Heparanase. Clin. Cancer Res. 2017, 23, 6267–6278. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lu, J.; Jiao, Y.; Chen, Q.; Li, M.; Wang, Z.; Yu, Z.; Huang, X.; Yao, A.; Gao, Q.; et al. Aspirin Inhibits Natural Killer/T-Cell Lymphoma by Modulation of VEGF Expression and Mitochondrial Function. Front. Oncol. 2018, 8, 679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, N.; Williams, J.; Rigas, B. NO-donating Aspirin Inhibits Angiogenesis by Suppressing VEGF Expression in HT-29 Human Colon Cancer Mouse Xenografts. Carcinogenesis 2008, 29, 1794–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, Z.; Wang, Z.; Zhang, Y.; Jia, Q.; Wu, L.; Zhang, W. Impact of Acetylsalicylic Acid on Tumor Angiogenesis and Lymphangiogenesis Through Inhibition of VEGF Signaling in a Murine Sarcoma Model. Oncol. Rep. 2013, 29, 1907–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadar, S.; Blann, A.D.; Lip, G.Y.H. Effects of Aspirin on Intra-Platelet Vascular Endothelial Growth Factor, Angiopoietin-1, and P-Selectin Levels in Hypertensive Patients. Am. J. Hypertens. 2006, 19, 970–977. [Google Scholar] [CrossRef]
- Gerrah, R.; Fogel, M.; Gilon, D. Aspirin decreases vascular endothelial growth factor release during myocardial ischemia. Int. J. Cardiol. 2004, 94, 25–29. [Google Scholar] [CrossRef]
- Jayaram, P.; Yeh, P.; Patel, S.J.; Sela, R.; Shybut, T.B.; Grol, M.W.; Lee, B.H. Effects of Aspirin on Growth Factor Release From Freshly Isolated Leukocyte-Rich Platelet-Rich Plasma in Healthy Men: A Prospective Fixed-Sequence Controlled Laboratory Study. Am. J. Sports Med. 2019, 47, 1223–1229. [Google Scholar] [CrossRef]
- Marx, R.E.; Carlson, E.R.; Eichstaedt, R.M.; Schimmele, S.R.; Strauss, J.E.; Georgeff, K.R. Platelet-rich plasma: Growth factor enhancement for bone grafts. Oral Surg. Endod. 1998, 85, 638–646. [Google Scholar]
- Hom, D.B. New Developments in wound healing relevant to facial plastic surgery. Arch. Facial Plast. Surg. 2008, 10, 402–406. [Google Scholar] [CrossRef]
- Van den Dolder, J.; Mooren, R.; Vloon, A.P.; Stoelinga, P.J.; Jansen, J.A. Platelet-Rich Plasma: Quantification of Growth Factor Levels and the Effect on Growth and Differentiation of Rat Bone Marrow Cells. Tissue Eng. 2006, 12, 3067–3073. [Google Scholar] [CrossRef] [Green Version]
- Aghaloo, T.L.; Moy, P.K.; Freymiller, E.G. Investigation of platelet-rich plasma in rabbit cranial defects: A pilot study. J. Oral Maxillofac. Surg. 2002, 60, 1176–1181. [Google Scholar] [CrossRef]
- Kuffler, D.P. Platelet-Rich Plasma Promotes Axon Regeneration, Wound Healing, and Pain Reduction: Fact or Fiction. Mol. Neurobiol. 2015, 52, 990–1014. [Google Scholar] [CrossRef] [PubMed]
- Javeed, A.; Hou, Y.; Duan, K.; Zhang, B.; Shen, H.; Cao, Y.; Zhao, Y. Aspirin Significantly Decreases the Nonopsonic Phagocytosis and Immunogenicity of Macrophages in Mice. Inflamm. Res. 2011, 60, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Javeed, A.; Ashraf, M.; Zhao, Y.; Mukhtar, M.M.; Rehman, M.U. Aspirin and immune system. Int. Immunopharmacol. 2012, 12, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, R.E.; Alford, P.B.; Xue, Y.; Thai, S.F.; Adams, D.O.; Pizzo, S. Aspirin inhibits tumor necrosis factoralpha gene expression in murine tissue macrophages. Mol. Pharmacol. 1997, 52, 421–429. [Google Scholar] [CrossRef]
- Jones, M.K.; Wang, H.; Peskar, B.M.; Levin, E.; Itani, R.M.; Sarfeh, I.J.; Tarnawski, A.S. Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs: Insight into mechanisms and implications for cancer growth and ulcer healing. Nat. Med. 1999, 5, 1418–1423. [Google Scholar] [CrossRef]
- Maity, G.; Chakraborty, J.; Ghosh, A.; Haque, I.; Banerjee, S.; Banerjee, S.K. Aspirin suppresses tumor cell-induced angiogenesis and their incongruity. J. Cell Commun. Signal. 2019, 13, 491–502. [Google Scholar] [CrossRef]
- Smadja, D.M.; Bura, A.; Szymezak, J.; Blanchard, A.; Remones, V.; Azizi, M.; Gaussem, P. Effect of clopidogrel on circulating biomarkers of angiogenesis and endothelial activation. J. Cardiol. 2012, 59, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.C.; Peng, Y.C.; Chen, T.S.; Huo, T.I.; Hou, M.C.; Huang, H.C.; Lin, H.C.; Lee, F.Y. Clopidogrel inhibits angiogenesis of gastric ulcer healing via downregulation of vascular endothelial growth factor receptor 2. J. Formos. Med. Assoc. 2016, 115, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Verdecchia, P.; Angeli, F.; Aita, A.; Bartolini, C.; Reboldi, G. Why Switch From Warfarin to NOACs? Intern. Emerg. Med. 2016, 11, 289–293. [Google Scholar] [CrossRef]
- Hart, R.G.; Pearce, L.A.; Aguilar, M.I. Meta-analysis: Antithrombotic therapy to prevent stroke in patients who have nonvalvular atrial fibrillation. Ann. Intern. Med 2007, 146, 857–867. [Google Scholar] [CrossRef]
- Gómez-Outes, A.; Suarez-Gea, M.L.; Lecumberri, R.; Terleira-Fernández, A.I.; Vargas-Castrillón, E.; Rocha, E. Potential role of new anticoagulants for prevention and treatment of venous thromboembolism in cancer patients. Vasc. Health Risk Manag. 2013, 9, 207–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.; Gonzales, I.; Pinto, M.P.; Arce, M.; Valenzuela, R.; Aranda, E.; Elliot, M.; Alvarez, M.; Henriquez, S.; Velasquez, E.V.; et al. Independent anti-angiogenic capacities of coagulation factors X and Xa. J. Cell Physiol. 2014, 229, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Pearson, T.; Manning, G.; Donnelly, R. In vitro study of thrombin on tubule formation and regulators of angiogenesis. Clin. Appl. Thromb. Hemost. 2010, 16, 674–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haralabopoulos, C.; Grant, D.S.; Kleinman, H.K.; Maragoudakis, M.E. Thrombin promotes endothelial cell alignment in Matrigel In Vitro and angiogenesis in vivo. Am. J. Physiol. 1997, 273, C239–C245. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.; Merchan, J.R.; Kale, S.; Sukhatme, V.P. Antiangiogenic property of human thrombin. Microvasc. Res. 2003, 66, 1–14. [Google Scholar] [CrossRef]
- Sabor, M.; Moinuddin, S.; Ilyas, S. Platelets structural, functional and metabolic alterations in diabetes mellitus. Pak. J. Physiol. 2012, 8, 40–43. [Google Scholar]
- Szablewski, L.; Sulima, A. The Structural and Functional Changes of Blood Cells and Molecular Components in Diabetes Mellitus. Biol. Chem. 2017, 398, 411–423. [Google Scholar] [CrossRef]
- Kaur, R.; Kaur, M.; Singh, J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: Molecular insights and therapeutic strategies. Cardiovasc. Diabetol. 2018, 17, 121. [Google Scholar] [CrossRef]
- Assert, R.; Scherk, G.; Bumbure, A.; Pirags, V.; Schatz, H.; Pfeiffer, A.F. Regulation of protein kinase C by short term hyperglycaemia in human platelets in vivo and in vitro. Diabetologia 2001, 44, 188–195. [Google Scholar] [CrossRef]
- Kampfer, H.; Pfeilchifter, J.; Frank, S. Expressional regulation of angiopoietin-1 and -2 and the tie-1 and -2 receptor tyrosine kinases during cutaneous wound healing: A comparative study of normal and impaired repair. Lab. Investig. 2001, 81, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Bitar, M.S.; Labbad, Z. Transforming growth factor-beta and insulin-like growth factor-1 in relation to diabetes-induced impairment of wound healing. J. Surg. Res. 1996, 61, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Rivard, A.; Fabre, J.E.; Silver, M.; Chen, D.; Murohara, T.; Kearney, M.; Magner, M.; Asahara, T.; Isner, J.M. Age-dependent impairment of angiogenesis. Circulation 1999, 99, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, I.; Fujita, M.; Miwa, K.; Hasegawa, K.; Kihara, Y.; Nohara, R.; Miyamoto, S.; Ueda, K.; Tamaki, S.; Sasayama, S. Age-dependent impairment of coronary collateral development in humans. Heart Vessel. 2000, 15, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Ouriel, K.; Veith, F.J. Acute lower limb ischemia: Determinants of outcome. Surgery 1998, 124, 336–341. [Google Scholar] [CrossRef]
- Gosain, A.; DiPietro, L.A. Aging and wound healing. World J. Surg. 2004, 28, 321–326. [Google Scholar] [CrossRef]
- Doria, G.; Frasca, D. Regulation of cytokine production in aging mice. Ann. N. Y. Acad. Sci. 1994, 741, 299–304. [Google Scholar] [CrossRef]
- Ershler, W.B.; Keller, E.T. Age-associated increased interleukin-6 gene expression, late-life diseases, and frailty. Annu. Rev. Med. 2000, 51, 245–270. [Google Scholar] [CrossRef]
- Mascarucci, P.; Taub, D.; Paloma, M.A.; Dawson, H.; Roth, G.S.; Ingram, D.K.; Lane, M.A. Age-related changes in cytokine production by leukocytes in rhesus monkeys. Aging 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Ashcroft, G.S.; Horan, M.A.; Ferguson, M.W. Aging is associated with reduced deposition of specific extracellular matrix components, an up- regulation of angiogenesis, and an altered inflammatory response in a murine incisional wound healing model. J. Investig. Dermatol. 1997, 108, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Swift, M.E.; Kleinman, H.K.; DiPietro, L.A. Impaired wound repair and delayed angiogenesis in aged mice. Lab. Investig. 1999, 79, 1479–1487. [Google Scholar]
- Costa, F.; Sores, R. Nicotine: A pro-angiogenic factor. Life Sci. 2009, 84, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Conklin, B.S.; Zhao, W.; Zhong, D.; Chen, C. Nicotine and cotinine up-regulate vascular endothelial growth factor expression in endothelial cells. Am. J. Pathol. 2002, 160, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Heeschen, C.; Jang, J.J.; Weis, M.; Pathak, A.; Kaji, S.; Hu, R.S.; Tsao, P.S.; Johnson, H.L.; Cooke, J.P. Nicotine stimulates angiogenesis and promotes tumor growth and atherosclerosis. Nat. Med. 2001, 7, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, J.; Jang, J.J.; Sundram, U.; Dayoub, H.; Fajardo, L.F.; Cooke, J.P. Nicotine accelerates angiogenesis and wound healing in genetically diabetic mice. Am. J. Pathol. 2002, 161, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Mousa, S.; Mousa, S.A. Cellular and molecular mechanisms of nicotine’s pro-angiogenesis activity and its potential impact on cancer. J. Cell Biochem. 2006, 97, 1370–1378. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | No OA | ASA | ASA + Clopidogrel | NOACs |
| Total number | 5 | 8 | 10 | 7 |
| Male/Female | 4/1 | 6/2 | 9/1 | 5/2 |
| Mean age ± SD (years) | 26.0 ± 3.8 | 71.8 ± 5.9 | 71.5 ± 6.4 | 67.0 ± 8.6 |
| moking (number of subjects) | 0 | 1 | 4 | 0 |
| Diabetes Mellitus (number of subjects) | 0 | 4 | 2 | 1 |
| B | No DM(No OA) | T1D(No OA) | T2D(No OA) | |
| Total number | 8 | 6 | 10 | |
| Male/Female | 4/4 | 4/2 | 1/9 | |
| Mean age ± SD (years) | 30.75 ± 3.99 | 39.0 ± 5.88 | 62.4 ± 18.55 | |
| BMI (kg/m²) | 20.87 ± 2.50 | 26.34 ± 4.92 | 40.7 ± 15.16 | |
| Blood glucoseat the time of blood collection (mg/dL) | 98.75 ± 5.58 | 126.84 ± 37.74 | 122.2 ± 31.58 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moog, P.; Jensch, M.; Hughes, J.; Salgin, B.; Dornseifer, U.; Machens, H.-G.; Schilling, A.F.; Hadjipanayi, E. Use of Oral Anticoagulation and Diabetes Do Not Inhibit the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes. Biomedicines 2020, 8, 283. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080283
Moog P, Jensch M, Hughes J, Salgin B, Dornseifer U, Machens H-G, Schilling AF, Hadjipanayi E. Use of Oral Anticoagulation and Diabetes Do Not Inhibit the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes. Biomedicines. 2020; 8(8):283. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080283
Chicago/Turabian StyleMoog, Philipp, Maryna Jensch, Jessica Hughes, Burak Salgin, Ulf Dornseifer, Hans-Günther Machens, Arndt F. Schilling, and Ektoras Hadjipanayi. 2020. "Use of Oral Anticoagulation and Diabetes Do Not Inhibit the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes" Biomedicines 8, no. 8: 283. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080283